( Description of empidoid wing-type according to ROHDENDORF, 1951)
Representatives of the type
To the lifting many-veined (empidoid) type belong [the] many species of the family Empididae and the rare relict flies Sciadoceratidae of the Southern Hemisphere [the latter family belongs, according to ROHDENDORF, 1964, to the superfamily Platypezidea (= Clythiidea), which itself belongs, not to the infraorder Asilomorpha [largely Brachycera-orthorrapha], to which the Empididea belong, but to the infraorder Myiomorpha [largely Brachycera-cyclorrapha].
Size of wings
Absolute size of wings is not large, their length does not exceed 10-11 mm, usually significantly smaller, sometimes not larger than 1 mm (some Corynetinae). Relative size of the wings is equal to the body, more rarely exceeding it a little. Magnitude of the surface of the wings is known only of certain species of the genera Empis (0.161-0.284 cm2), Ocydromia (0.057 cm2) and Noeza (0.070 cm2) : These figures do not characterize the surface area of the larger species of the genus Empis and especially [not characterize] the many minute forms possessing small wings. The weight of the wings is not known. The load is studied in the above mentioned forms, and is equal to 0.072-0.098 gr/cm2 in representatives of the genus Empis, 0.035 gr/cm2 in those of Ocydromia, and 0.028 gr/cm2 in Noeza.
Some examples of wings of the empidoid functional type :
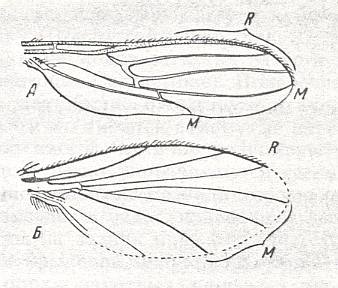
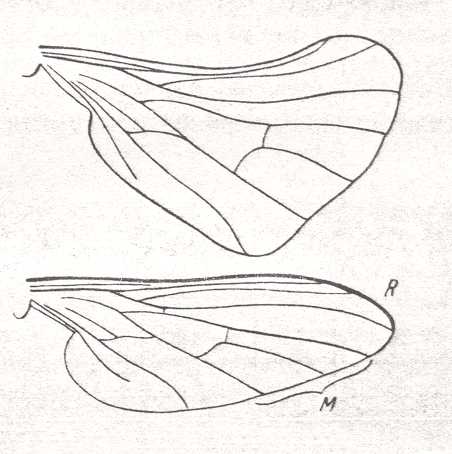
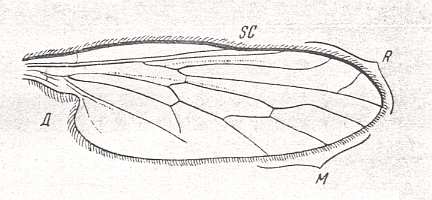
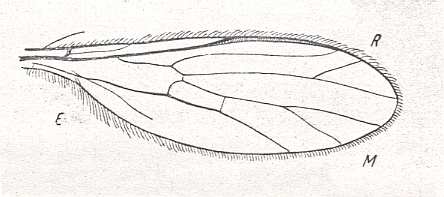
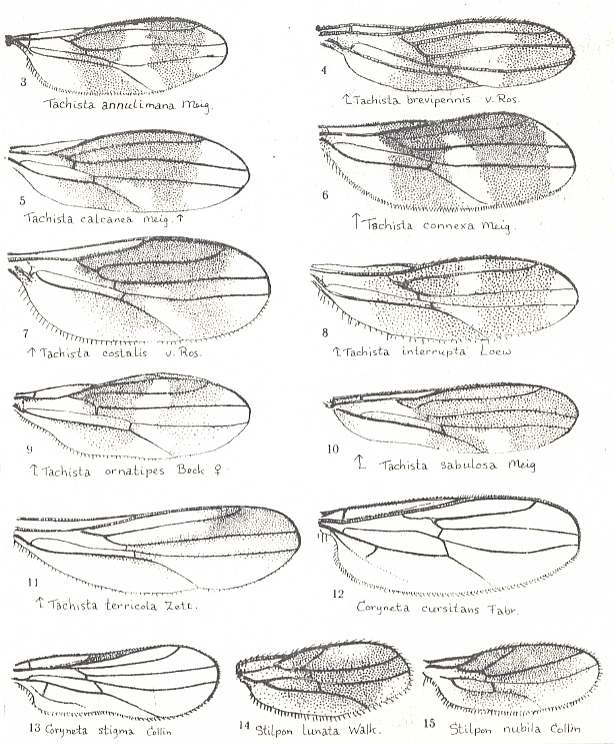
Figure 1. Wings of representatives of the lifting many-veined (empidoid) type. From top to bottom :
Tachista brevipennis Ros., Empididae. Length about 1.5 mm. (After ENGEL)
Sciadocera patagonica Schm., Sciadoceratidae [= Sciadoceridae]. Length about 4 mm. (After SCHMITZ)
Rhamphomyia platyptera Panz., Empididae. Wing of female.(After LINDNER)
Rhamphomyia platyptera Panz., Empididae. Wing of male. (After LINDNER)
Homalocnemis nigripennis Phil., Empididae. Length about 5 mm. (After COLLIN)
Ceratomerus mediocris Coll., Empididae. Length about 3 mm.(After COLLIN)
(All in ROHDENDORF, 1951)
Shape of the wings
To the above images of empidoid wings, given by ROHDENDORF, we here add some from HENNIG, 1954 :
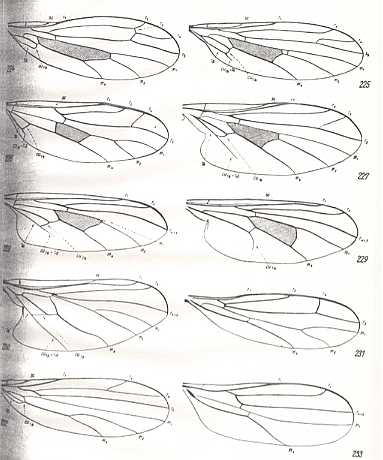
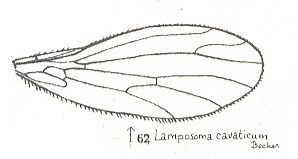
The last three wings of the above Figure (231, 232, 233), although being wings of empidids, may not belong to the lifting many-veined (empidoid) functional wing-type, as also the following empidid wings :
Nevertheless, ROHDENDORF does consider, for example, the wings of Tachista brevipennis, Figure 1, top image ( Empididae) to belong to the present functional wing-type, despite its rather poor venation. Apart from 231 (Hemerodromia) and 232 (Asymphyloptera), 233, Tachista, may belong to ROHDENDORF's corynetoid subtype of empidoid wings, described below, while 231 may belong to ROHDENDORF's atalantoid subtype.
Skeleton of the wing (of the empidoid type)
Coverings of the wing
Functional features
History and transformations of the type
1. The first, undoubtedly original subtype, possessing the least changed wings with rich venation is the brachystomatoid subtype, for which is characteristic the presence of a long Subcostal vein, four Radial [including the Radius proper] and three Medial branches, and, especially, a long cubital cell formed by the converging cubital and anal veins [CuA and 1A]. In addition, the Costal vein runs all around the wing although it is thinner and more delicate along the wing's hind-margin. This subtype includes few, clearly relict forms of the subfamily Brachystomatinae (the genera Brachystoma, Homalocnemis, see Figure 1, fifth image, and others) and is a connecting link between the tabanoid (cyrtosioid subtype) ( Part LXXI, 9th subtype) and the lifting many-veined (empidoid) types.
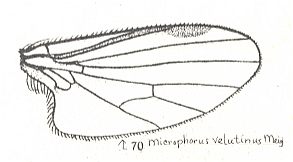
2. The most diverse subtype, including the main mass of empidids, is the microphoroid subtype, for which it is characteristic the reduction [i.e. shortening] of the cubital cell, being more simple [i.e. shorter] than the [so-called] posterior basal [cell] [with the latter apparently is meant the cell lying directly above it] (as a result of shortening and curving of the Cubital vein), the weakening of the Subcostal vein, the frequent reduction of the fourth radial vein [i.e. the 3rd branch of the Radial Sector] and the strong development of the anal lobe reaching a large size. A rudimentary phragm is present in the basiala. Costalization of the venation is strongest expressed precisely in this subtype. To it belong representatives of the subfamilies Empidinae, Figure 1, 3rd and 4th image, Ocydromiinae and Noezinae, the most species-rich groups of Empidids. This subtype was the point of departure for the origin of the vast muscoid type [later to be introduced].
To ROHDENDORF's figures of wings of the microphoroid subtype may be added that of Microphorus velutinus (Empididae) :
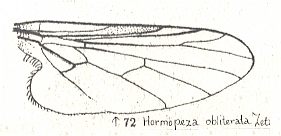
and that of Hormopeza obliterata (Empididae), in which the Radial Sector is 3-branched :
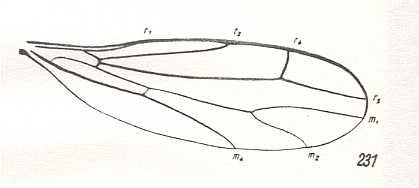
3. Fairly diverse groups together form the special atalantoid subtype, characterized by the development of peculiar wings narrowed in their basal part. Anal lobe not projecting, in the form of a smooth outgrowth. The cubital cell and the number of Radial and Medial branches as in the previous subtype. Sometimes the cubital cell is totally reduced (Ceratomerinae, Figure 1, bottom image ). Characteristic is the shortening of the Subcostal and first Radial vein [i.e. the Radius proper] never running beyond the middle of the anterior wing-margin. To this subtype belong many species of the subfamilies Atalantinae, Hemerodromiinae, and Ceratomerinae.
To ROHDENDORF's figure of a wing of this subtype we must add some more wings. Note that in them, as also in the wing depicted by ROHDENDORF, the Radial Sector is 3-btanched :
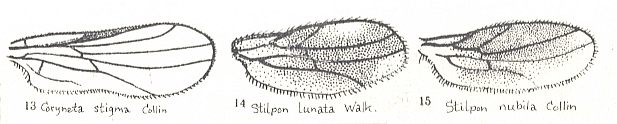
4. A very peculiar structure of the wings is found in representatives of the corynetoid subtype, which is characterized by a decrease of absolute size (up to 1 mm!) and by the relative lengthening of the wings, almost always lacking the cubital cell, the Subcostal vein, and a projecting anal lobe. Media always with only two branches. Intermedial cell [= discoidal cell] coalesced with the posterior basal one [i.e. now forming just one single cell] and shifted to the base of the wing. The wing as a whole has a characteristic tongue-like shape. To this subtype belong the characteristic species of the subfamily Corynetinae (= Tachydromiidae of earlier authors!, Figure 1, first image ).
To the one Figure given by ROHDENDORF of a wing representing the corynetoid subtype we may add the following from LINDNER, Die Fliegen der palaearktischen Region :
5. A very special structure of the wings is found in the sciadoceratoid subtype. Shape of wings shortened. The wings have an all-out convex anterior margin and a still more convex posterior margin. Subcostal vein very short, ending up in the Radius. The three Radial veins are unbranched. The Medial veins are weaker, three branches. There is almost no cubital vein. All cross-veins have shifted towards the base of the wing. Anal lobe weakly convex. The basiala is individualized, apparently with a rudimentary phragm. The wing as a whole has a very characteristic look as a result of its radiating straight veins. To this subtype belong the few relict forms of the family Sciadoceratidae ( second image of Figure 1 ).
Evaluating the variability and evolutionary pathways of the empidoid type, one can clearly see the microphoroid wings as representing the most progressive subtype (= 2nd subtype of the present type), in the wings of which one finds the most clearly expressed processes of costalization, together with the individualization and strenghening of the basiala. The other subtypes show various narrow-specialized features : such as the narrowing of the wings and the disappearance of the anal lobe, which we saw in the atalantoid wings or in the radiating venation in the sciadoceratoid subtype. Finally, a significant narrowing of the wings, decrease of their general size, dilution of the venation, the presence, in addition, of "reduced-winged" or even of totally wingless forms -- all this is characteristic of the corynetoid subtype and non-flying empidids [the latter] undoubtedly having originated from those who had wings of that subtype. It is completely legitimate to consider the corynetoid subtype as representing a regressive form of empidoid wings. The first, primitive brachystomatoid subtype is still insufficiently known, and judging more precisely about its evolutionary significance is not yet possible.
In the next document we will set up the venational derivational lines and morphoklines of the representatives of the empidoid type.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXXIIIa.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII
Back to Evolutionary Part LXVIII
Back to Evolutionary Part LXIX
Back to Evolutionary Part LXXa
Back to Evolutionary Part LXXb
Back to Evolutionary Part LXXI
Back to Evolutionary Part LXXI
Back to Evolutionary Part LXXII
Back to Evolutionary Part LXXIIa