
[Also in these enquiries into the wing-venation and its transformations in Diptera-Nematocera we will consider the subcostal vein (Sc) at most only occasionally. This vein, especially its terminal branches (Sc1, Sc2), is -- in Diptera -- often difficult to discern.]
We will now discuss the wing-venation and its transformations within, and also (departing from the Rhyphidea) beyond, the infraorder Bibionomorpha (itself still belonging -- as do the Tipulomorpha and the Blephariceromorpha -- to the suborder Nematocera). This infraorder includes first of all the large complex of fungivoroids (fungus-gnats and allies) and the cecidomyiideans (gall-midges), but also the scatopsideans, the bibionideans, and the rhyphideans. Collectively we could call them the "land-midges", contrasting with the water-midges (Tipulomorpha).
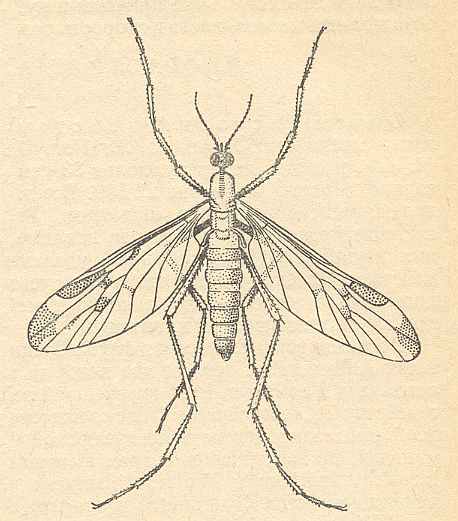
The superfamily Rhyphidea includes three recent families : Rhyphidae [ = Phryneidae] (about 66 recent species), Olbiogastridae (31 species), and Cramptonomyiidae (1 species, Cramptonomyia spenceri ALEX.) [As to the Olbiogastridae I do not yet have enough data]. [Cramptonomyia might as well be placed in the family Rhyphidae.]
The wing-venation of the recent Rhyphidae possesses a 2-branched Radial. But if the interpretation of the recent Cramptonomyiidae is correct, the venational groundplan of the Rhyphidea (or Rhyphidae for that matter) possesses a 3-branched Radial Sector, although only two of them end up in the wing-margin while a third ends up in one of the other two branches of Rs. See next Figure.

Figure 2 : Wing-venation of Cramptonomyia spenceri ALEX. Order Diptera, Family Cramptonomyiidae (or just Rhyphidae), superfamily Rhyphidea, Suborder Nematocera.
(After ALEXANDER, 1931, from HENNIG, 1954)
So thanks to the recent Cramptonomyia we can hold that the venational groundplan of the Rhyphidea has a 3-branched Radial Sector. And this is confirmed by fossil forms (upper Lias [ = youngest period of the oldest period of the Jurassic] ) having a clearly rhyphoid wing-venation with a fully 3-branched Radial Sector, that is, with all three branches ending up at the wing-margin. One of these fossils we therefore designate to possess the venational groundplan of the superfamily :
The venational groundplan of the superfamily Rhyphidea can be represented by the venation as we see it in the Liassic Protorhyphus stigmaticus HANDL. [This, and some other species are usually assigned to a separate (fossil) family, the Protorhyphidae.] :

Figure 3 : Wing of Protorhyphus stigmaticus HANDL. Order Diptera, Family Protorhyphidae. Upper Lias of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)
Other fossil Rhyphidea, among which Protorhyphidae, are depicted in the next Figures :
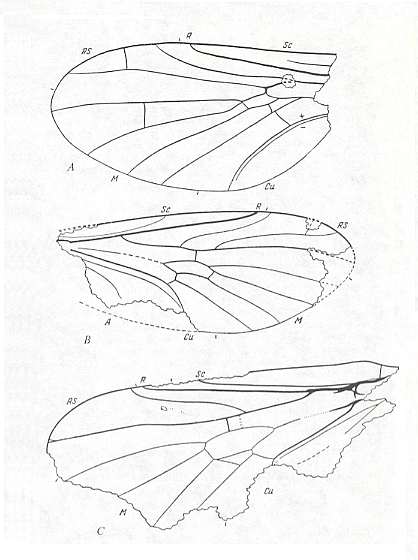
Figure 4 : Rhyphidea from the lower Jurassic of Issic Kul (central Asia).
A -- Oligophryne fungivoroides ROHD. (Oligophryneidae). Holotype, Coll. PIN No. 358/120. Length of fossil 1.65 mm. Length of whole wing approximately 2.0 mm.
B -- Protorhyphus turanicus ROHD. (Protorhyphidae). Holotype, Coll. PIN No. 371/1. Length of fossil 2.7 mm. Length of whole wing about 3.0 mm.
C -- Protolbiogaster rhaetica ROHD. (Protolbiogastridae). Holotype, Coll. PIN No. 371/114. Length of wing 4.7 mm.
(After ROHDENDORF, 1962, from ROHDENDORF, 1964)
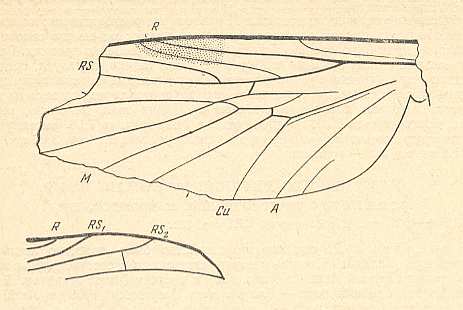
Figure 5 : A representative of the Rhyphidea from the middle Jurassic of Karatau (southern Kazachstan) : Archirhyphus asiaticus ROHD. (Protorhyphidae). Holotype, Coll. PIN No. 2452/334.
Upper image : Left wing, length 2.9 mm, width 1.05 mm.
Lower image : Distal part of right wing.
(After ROHDENDORF, 1964)
The Radial Sector of the Protorhyphidae is 3-branched (all three branches ending up in the wing-margin). We can also describe this same Radial Sector as being one vein (branching off from the Radius) from which branch off two anterior branches, one before, and the other after the radiomedial cross-vein ta ( = r-m). If the second of the just mentioned two anterior branches evolutionarily disappears (as is indicated by Cramptonomyia -- see Figure 2 ) we obtain a venation much like that of recent Rhyphidea (in any case of the Rhyphidae s.str.) :
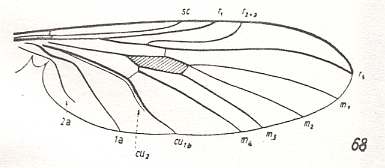
Figure 6 : Wing-venation of Phryne fuscipennis MACQ. Order Diptera, Family Rhyphidae, Superfamily Rhyphidea, Suborder Nematocera. Recent.
(After HENNIG, 1954)
This concludes our investigation of the wing-venation -- groundplan and further development -- in the superfamily Rhyphidea.
But apart from this there can be said much more about this superfamily : It is, namely, supposed that the Rhyphidea -- including within itself in addition to the recent families Rhyphidae, Olbiogastridae, and Cramptonomyiidae, also a number of fossil families such as Oligophryneidae and Protolbiogastridae (see Figure 4A and 4C ) and, most importantly, the (fossil) family Protorhyphidae (see Figure 4B and Figure 3 and Figure 5 ) -- gave rise to all the higher flies. So because of the central position of this superfamily within the system and history of Diptera we will dwell upon them much longer before continuing the investigation of the wing-venation in the other Bibionomorpha.
We hypothesize that all the families of the suborders Brachycera-orthorrapha and Brachycera-cyclorrapha arose from a rather broad stock of lower Jurassic or upper Triassic insects that belong within and around the fossil family Protorhyphidae ( Figure 4B and Figure 3 and Figure 5 ). Because these lower Jurassic fossils (Protorhyphidae) have been found in three widely separated locations (Germany, Kazachstan, and central Asia) we can assume that the Protorhyphidae and close relatives lived in many locations in the Eastern Hemisphere and undoubtedly have been present in the Western as well. Based on this, we can further assume that they were rather diverse, at least diverse enough to give rise to all the families of the two mentioned suborders. They -- the Protorhyphidae and close relatives -- in turn must have evolved from a rather broad particular mecopterous stock (that is, a stock of scorpion-flies, Mecoptera) somewhere in the late Permian or early Triassic, together with the other bibionoid and tipuloid lines that arose from other mecopterous stocks.
Another important fossil fly is Protobrachyceron liasinum HANDL. :
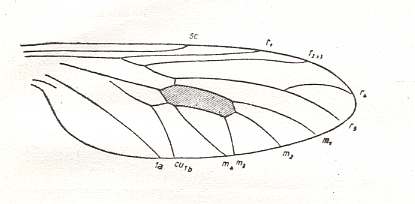
Figure 7 : Wing of Protobrachyceron liasinum HANDL. Upper Lias of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)
Apart from the suppression of the cross-vein tb ( = m-cu) (in fact the suppression of its anterior part, tb1) and the shorter last fork of Rs, this venation looks exactly like that in Protorhyphidae : compare, for example, with Figure 5 . So I don't know why this form was determined to belong already to true Brachycera-orthorrapha, instead of simply belonging to the nematocerous family Protorhyphidae. And, of course, we could think the other way around : Are the "Protothyphidae" perhaps already true Brachycera-orthorrapha? ( Problems like this one naturally occur because most fossil insects are represented by their wings only).
We will interpret the wing-venation of all primitive Brachycera-orthorrapha, including those whose venation leads to that in Brachycera-cyclorrapha, against the background of the venation as it is in the (fossil) Protorhyphidae and close allies, because they are supposed to have given rise to the mentioned Brachycera. Let us depict one of them again (Figure 3) :
Figure 3 : Wing of Protorhyphus stigmaticus HANDL. Order Diptera, Family Protorhyphidae. Upper Lias of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)
The characterization of the wing-venation in Protorhyphidae (allowing some play of variability), insofar as it is relevant to the venational features in the primitive Brachycera which (Brachycera) evolved from them, can read as follows (see also diagram below) :
Radial Sector 3-branched : One long vein (ending up at the wing-margin) from which branch off two anterior veins (also ending up at the wing-margin) : One branching off before the radio-medial cross-vein ta ( = r-m), the other after it. This cross-vein ta connects the Radial Sector (more precisely R4+5) with the anterior boundary of the Discoidal cell. This cell begins (reading the wing from base to tip) where the chief trunk of the Media (which, if we depart from the mecopterous venation, in fact is M1+2+3+4) divides into M3+4 and M1+2. The proximal part of the posterior boundary of the discoidal cell is formed by M3+4. Halfway this boundary M3+M4 splits up into M3 and M4. The posterior boundary then continues as M3, while M4, after it connects with CuA ( = Cu1b) (in fact with the (cross-)vein, tb2, connecting CuA with M4), continues its course to the wing-margin, at which it finally ends up. The basal piece of M4 can be called tb1 ( = anterior part of the "cross-vein" tb), while the alledged basal piece of Cu1a ( = the anterior branch of a supposedly (according to some authors) forked Cu1 (CuA)) can be called tb2 ( = posterior part of the cross-vein tb). (See diagram below)
The anterior boundary of the discoidal cell is formed by M1+2. At the end of the cell M1+2 splits into M1 and M2. The apical boundary of the Discoidal Cell consists of two parts : the anterior part is the basal section of M2, while the posterior part is the cross-vein tp ( = intermedial cross-vein). In wings where this cell is lost this usually has come about by the loss of this cross-vein.
The Anterior Cubitus, CuA (Cu1b), arches down to the wing-margin and reaches it.
The First Anal Vein, 1a (1A), is long and well developed, it reaches the wing-margin not far from the endpoint of CuA.
Later in evolution CuA and 1A meet before reaching the wing-margin, their common stalk reaching the wing-margin. The space then enclosed between CuA and 1A is called the Anal Cell. See next diagram.

The next Figures depict wings of fossil flies (middle Jurassic) that have been assigned to the suborder Brachycera-orthorrapha :
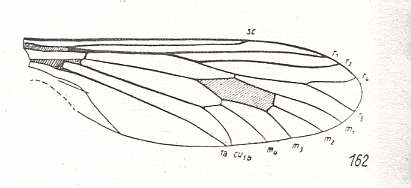
Figure 8 : Wing of Archirhagio obscurus ROHD. Middle Jurassic of Karatau, southern Kazachstan. Length of wing 13 mm.
(After ROHDENDORF, 1938, from HENNIG, 1954)
The venation of this wing totally complies with that of the Protorhyphidae.
Apart from a small extra cross-vein the next wing also so complies :
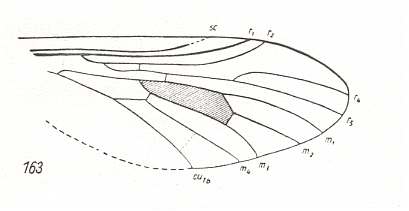
Figure 9 : Wing of Protorhagio capitatus ROHD. Middle Jurassic of Karatau, southern Kazachstan. Length of wing 5.0 mm.
(After ROHDENDORF, 1938, from HENNIG, 1954)
And, insofar as preserved, also the next wing complies with those of Protorhyphidae (Perhaps here M4 is not separated from the Discoidal Cell, forming its posterior boundary) :
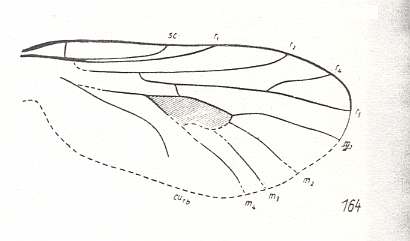
Figure 10 : Wing of Rhagionempis tabanicornis ROHD. Middle Jurassic of Karatau, southern Kazachstan. Length of wing 3.5 mm.
(After ROHDENDORF, 1938, from HENNIG, 1954)
In the next wing this is clearly the case, that is, M4 forms the posterior boundary of the Discoidal Cell :
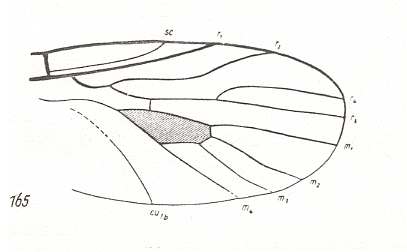
Figure 11 : Wing of Palaeostratiomyia pygmaea ROHD. Middle Jurassic of Karatau, southern Kazachstan. Length of wing 2.5 mm.
(After ROHDENDORF, 1938, from HENNIG, 1954)
We can compare these fossils, together with those of the Protorhyphidae, with the wings of recent representatives of the order Brachycera-orthorrapha :

Figure 12 : Wing-venation of Rhagio (Asilomorpha-Rhagionidae) (Diptera). Recent.
1A = first anal vein (often denoted as A1, or An1 , or 1a).
2A = second anal vein (often denoted as A2, or An2 , or 2a).
R = radial system of veins. M = medial system of veins.
Cu1 (often denoted as CuA, or Cu1, or Cu1b) = anterior branch of cubital system of veins (the posterior branch [ Cu2 , CuP ] is in Diptera either a fold or is absent altogether).
(After RICHARDS & DAVIES, 1977, Imms' General Textbook of Entomology)
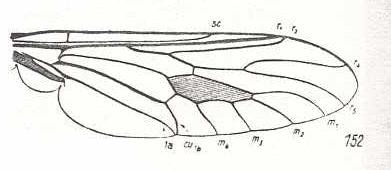
Figure 13 : Wing-venation of Coenomyia ferruginea SCOP. Recent. Order Diptera, Family Erinnidae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
Here (Figure 13) M4 forms part of the posterior boundary of the Discoidal Cell (Anterior part of tb (tb1) lost). Further, the ends of CuA and 1A have come closer together : Anal Cell almost closed. (CuA = Cu1b, 1A = 1a)
In the next wing also tb1 is lost :
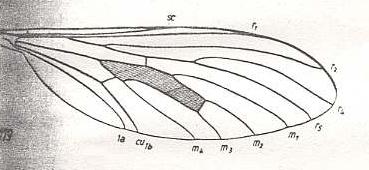
Figure 14 : Wing-venation of Leptogaster cylindrica DEG. Recent. Order Diptera, Family Asilidae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
The next wing is the first one of a series of wings of recent Empididae (still Brachycera-orthorrapha). The venation in this family is well under way to become the venation as we shall see it in all Brachycera-cyclorrapha (such as fruit-flies, blowflies and the house-fly) :
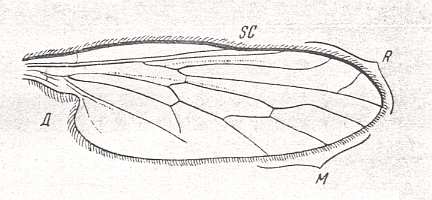
Figure 15 : Wing of Homalocnemis nigripennis PHIL. Recent. Order Diptera, Family Empididae, subfamily Brachystomatinae, Suborder Brachycera-orthorrapha. Length of wing approximately 5 mm. (After COLLIN, from ROHDENDORF, 1951)
The venation of this wing (Figure 15) is the most primitive within the Family Empididae, that is, most primitive with respect to the morphological ascent that eventually leads to the muscoid wing-venation (as we see it in most Brachycera-cyclorrapha). In it the Radial Sector is still 3-branched, like in Protorhyphidae and Rhagionidae. The Discoidal Cell is still present, as it will be in most Brachycera-cyclorrapha. M3 has already vanished as a result of the coalescence of M3 and M4 : M4, instead of M3, now forms the posterior boundary of the Discoidal Cell. The anal cell (in fact more properly called the cubital cell -- as ROHDENDORF (1951) does) is closed and its contraction has just begun (the stalk CuA+1A still very short).
With this form as a point of departure we see the venation in Empididae successively be transformed along a number of independent lines (morphoklines) : The Radial Sector becomes 2-branched (as it is in all Brachycera-cyclorrapha), the anal cell further contracts as a result of the shift of the endpoint of CuA along 1A toward the wing-base, at last resulting in a very short cell.
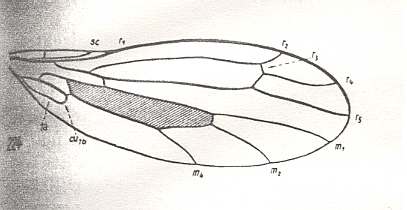
Figure 16 : Wing-venation of Dolichocephala irrorata FALL. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
Here (Figure 16) the Anal Cell has already become very small, that is, CuA has shifted its endpoint up along 1A all the way to the wing-base (a transitional stage of this can be seen in Figure 15 ). In addition, the common end-stalk of CuA and 1A has retreated from the wing-margin. Further, the Radial Sector (Rs) seems to have here four branches (instead of three), of which one does not end at the wing-margin but in R2 instead. This reminds one of the 4-branched Rs in Tanyderidae (Suborder Nematocera) (but here all ending up at the wing-margin). However, in the Suborder Brachycera, such a 4-branched Rs only occurs in Dolichocephala irrorata, that is, in the species depicted here (Figure 16). We consider the veinlet connecting R4 and R2 not to represent R3, but to be just an extra cross-vein, or a re-activation of an old "bed", and thus consider this species to have simply a 3-branched Rs like in Protorhyphidae and most other Brachycera-orthorrapha.
M4 forms the posterior boundary of the Discoidal Cell, and that means that in this form M3 has vanished.
Apart from a small extra cross-vein the next wing is similar to the previous one, except that it has the 'normal' 3-branched Rs, and that a small piece of CuA+1A has been preserved :
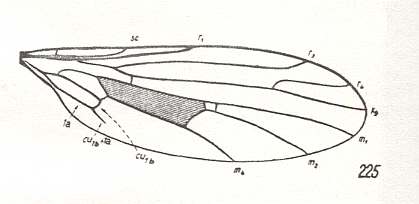
Figure 17 : Wing-venation of Clinocera inermis LW. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
The next wing is more or less like the previous one, except that R4 has vanished, leaving the Radial Sector only 2-branched as in all Brachycera-cyclorrapha, and that M1 is reduced (Only a vestige is left, indicated by the arrow) :
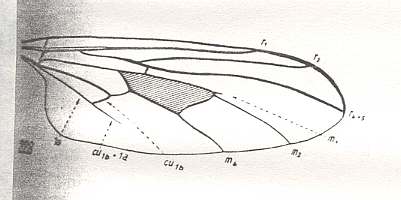
Figure 18 : Wing-venation of Leptopeza flavipes MEIG. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
This 2-branched Radial Sector we also find in Rhamphomyia (Empididae) :
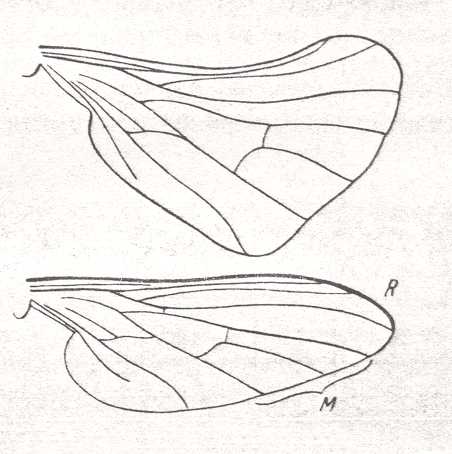
Figure 19 :
Upper image : Wing of female of Rhamphomyia platyptera PANZ. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha.
Bottom image : The same, wing of male.
(After LINDNER, from ROHDENDORF, 1951)
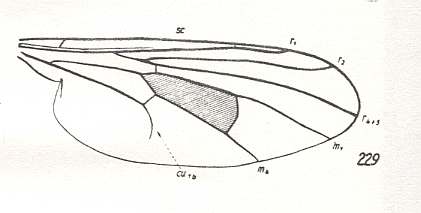
Figure 20 : Wing-venation of Hybos culiciformis FABR. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
This form (Figure 20) belongs to a line (morphokline) in which the formation and contraction of the anal cell was well underway, although the contraction had not yet proceeded very far. The Radial Sector had already become 2-branched, and M2 had vanished. Then the first anal vein (1A [ = 1a) was totally reduced.
Whithin the family Empididae the wing-venation in a number of species has proceeded further into the direction of reduction of the cubito-anal veins (in this family normally expressed by the contraction of the anal cell and the reduction of the CuA+1A stalk) : Complete reduction (i.e. vanishing) of CuA as well as 1A :
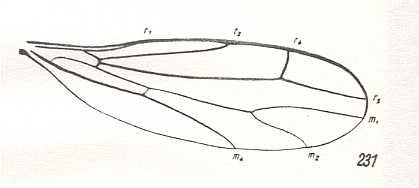
Figure 21 : Wing-venation of Hemerodromia raptoria MEIG. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
Apart from the mentioned reductions this form (Figure 21) shows even more of them : the Discoidal Cell has disappeared [The Discoidal Cell, where it is present, begins with the basal piece of M4 (or of M3+4 for that matter) (see Figure 17). In the present form (Hemerodromia) it is not closed distally]. So the venation of this wing seems to indicate a late development with respect to that in many other representatives of the family. But strangely enough, its Radial Sector is 3-branched !
If the Radial Sector in this species is really in an original state, as it is, for example in Figure 17 , then we cannot derive this wing (Figure 21) from say that of, say, Figure 19 , because in that case we have to assume a reversal of the evolution of the Radial Sector (from 2-branched to 3-branched). It can, however, be derived from that of Figure 17 , but only formally so : Although the number of branches of the Radial Sector is equal in both cases, its shape is totally different. So it is clear that the wing-venation in Hemerodromia is part of a separate line of venational development within the family Empididae, as probably so many (wing-venations) are.
The next wing shows the same kind of reduction in the cubito-anal region, but the Discoidal Cell is still present, and also here the Radial Sector is stil 3-branched :
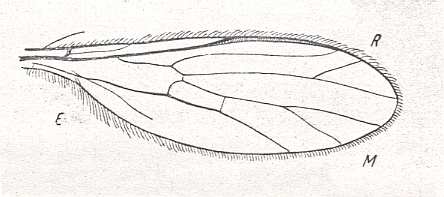
Figure 22 : Wing of Ceratomerus mediocris COLL. Recent. Order Diptera, Family Empididae, subfamily Ceratomerinae, Suborder Brachycera-orthorrapha. Length of wing approximately 3 mm.
(After COLLIN, from ROHDENDORF, 1951)
From the foregoing examples it is clear that the ascent up to the muscoid wing (as we see it in most Brachycera-cyclorrapha) has proceeded along several different and independent lines. Each such a line represents a transformational series (a morphokline), and each one results in a muscoid wing. However, these muscoid wings are not identical to each other, but different. ROHDENDORF, 1951, distinguishes 14 subtypes of muscoid wings!
The wing in the next Figure belongs to the only representative of the family Hilarimorphidae -- itself standing at the beginning of the typical empidid series -- namely Hilarimorpha singularis EGG. :
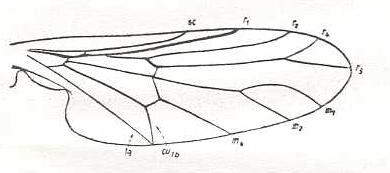
Figure 23 : Wing-venation of Hilarimorpha singularis EGG. Recent. Order Diptera, Family Hilarimorphidae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
In this wing (Figure 23) we again find the Radial Sector in its relatively primitive condition of being 3-branched. And also the anal cell is still in its primitive condition (CuA and 1A meeting just at the wing-margin, anal cell still very long). But in contrast to these relatively original conditions we see that the Discoidal Cell has disappeared [The Discoidal Cell, where it is present, begins with the basal piece of M4 (or of M3+4 for that matter) (see Figure 17). In the present form (Hilarimorpha) it is not closed distally]. So also here we have in one single wing a mixture of (relatively) derived (apomorphous) and original (primitive, plesiomorphous) characters, implying that this form (Hilarimorpha) cannot so easily (formally) be derived from another member of its circle of close relatives (here the Empididae).
The next wing is that of a primitive (with respect to wing-venation at least) member of the Brachycera-cyclorrapha, namely a representative of the family Clythiidae ( = Platypezidae) :
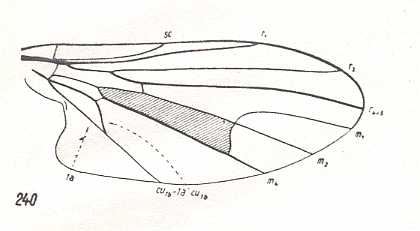
Figure 24 : Wing-venation of Platypezina connexa WAHLB. Recent. Order Diptera, Family Clythiidae ( = Platypezidae), Suborder Brachycera-cyclorrapha.
(After HENNIG, 1954)
In this form (Figure 24) the Radial Sector is 2-branched like in all Brachycera-cyclorrapha. The bifurcation-point of M1+2 lies on the boundary of the Discoidal Cell. M4 forms the posterior boundary of that cell : M3 has vanished, the anterior part of tb (tb1) is suppressed. The shift of the end-point of CuA (Cu1b) toward the wing-base is in its intermediate state. The long stalk CuA+1A (Cu1b+1a) reaches the wing-margin. The Discoidal Cell is still present.
In the next Figure we depict a true muscoid wing (Brachycera-cyclorrapha) :
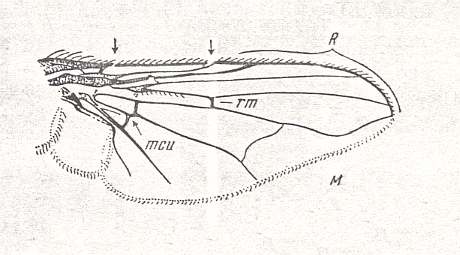
Figure 25 : Wing of Graphomyia maculata SCOP. Recent. Order Diptera, Family Muscidae, Suborder Brachycera-cyclorrapha.
Specimen No. 3770. Length of wing 9.5 mm.
(After ROHDENDORF, 1951)
Rs is, of course, 2-branched in this wing (Figure 25). M2 has vanished. The upward bending vein is M1. Discoidal Cell present, large. Shifting of the endpoint of CuA toward the wing-base has proceeded far. CuA almost recurrent. CuA+1A still long, but not reaching the wing-margin. Strong development of the Alula. After the endpoint of R5 the Costa is reduced, leaving the posterior wing-margin without costal vein (as in many Diptera).
The next wing has about the same venation :
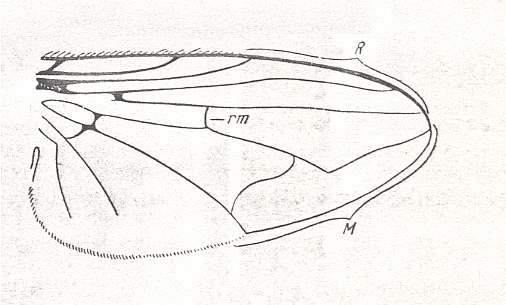
Figure 26 : Wing of Phasia crassipennis FABR. Recent. Order Diptera, Subfamily Phasiinae, Family Tachinidae, Suborder Brachycera-cyclorrapha. Male. Length of wing approximately 11 mm. (After ROHDENDORF, 1951)
The next two wings (Figure 27) also belong to representatives of the Brachycera-cyclorrapha, but being assigned to quite different subgroups of this suborder than have the other two (Figure 25 and 26) :
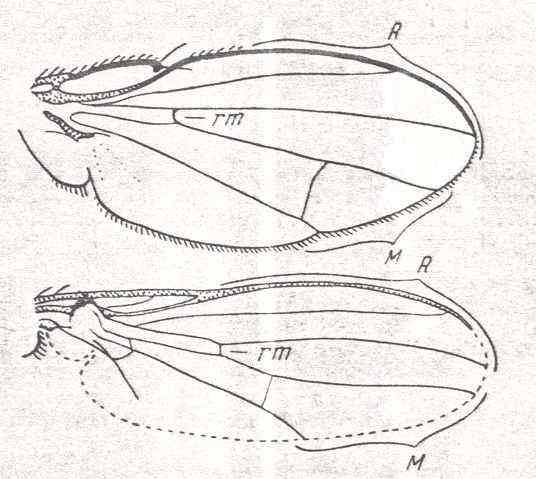
Figure 27 : Some more wings of representatives of the Brachycera-cyclorrapha.
Top image : Dichaeta caudata FALL. Ephydrinae. Specimen No. 2766. Length of wing 4 mm.
Bottom image : Sepsis communis MEIG. Sepsidae. Specimen No. 2897. Length of wing 2.25 mm.
(After ROHDENDORF, 1951)
Also here (Figure 27) we see the Radial Sector being 2-branched and also having the same shape (that is, the Radial Sector splitting up already very soon [proximally of the cross-vein r-m] resulting in two long branches) as in all other Cyclorrapha. But the reductions in the cubito-anal region have proceeded further.
In the next document we will take a closer look at the possible transformational lines -- especially at the fact that there are more than one such lines (morphoklines) -- leading, in a parallel fashion -- from the protorhyphoid wing-venation to the several different types of muscoid wing-venation.
REMARK :
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXII.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII