
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) At the end of the previous document we set out what to do next -- now that we have finished the systematic treatment of all (larger) groups of Diptera-Nematocera (crane-flies, mosquitoes, fungus-gnats, black-flies, gall-midges, etc.) -- before we continue with the systematic treatment of the remaining groups of the Diptera. We give it here once more, because it is an instructive introduction to the present document and to some documents following upon it :
The order Mecoptera contains a number of suborders :
The order Mecoptera is known to have existed already in the lower Permian (that is, before the age of the Dinosaurs), and probably even already in the upper Carboniferous. It flourished in upper Permian times but gradually began to decline during the Mesozoic. Today it is just a relict group.
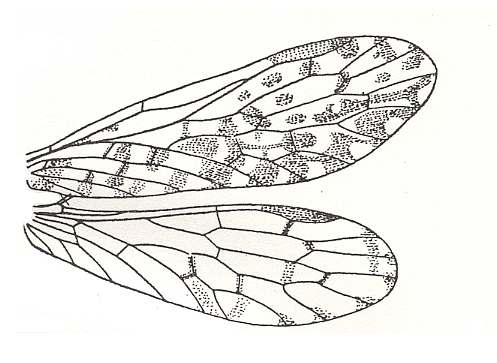
Now let us, to begin with, depict some recent representatives of the order Mecoptera (Scorpion-flies). This small order comprises less than 400 species in the recent fauna, the greater number of which belong to the genera Panorpa and Bittacus. See next Figure.

Figure :1
1 - Panorpa communis, Family Panorpidae, Order Mecoptera. Length 20 mm, wing-span 25-30 mm.
2 - Bittacus italicus, Family Bittacidae, Order Mecoptera. Wing-span 35-40 mm.
(Afer ZAHRADNIK and SEVERA, 1977, Thieme's insektengids)
The Mecoptera are -- like the Diptera, Hymenoptera (bees, wasps, ants), Lepidoptera (butterflies and moths), Coleoptera (beetles), etc. -- holometabolous insects, meaning that the larva and adult of the same individual are totally different from each other structurally as well as in their way of life. Between these phases there is inserted a resting phase, the pupa. In non-holometabolous insects the successive individual developmental stages are not so different from each other, and a pupal stage is absent.
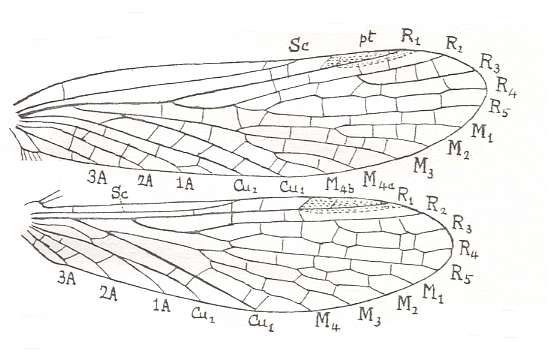
Figure 2 : Wings of Chorista australis KLUG. Order Mecoptera, Suborder Eumecoptera, Family Choristidae. Recent, Australia.
We can clearly see the chief systems of longitudinal veins : Sc = Subcosta, R1-R5 = Radial System (R1 = Radius, R2-R5 = Radial Sector), M1-M4 = Medial system, Cu1-Cu2 = Cubital system, 1A-3A = Anal system. pt = pterostigma, which is a more or less chitinized spot found in many insect wings. (After TILLYARD, 1935)
Figure 3 : Nannochorista holostigma TILL., male, (ca. x 11), in position of rest. Order Mecoptera, Family Nannochoristidae. (After TILLYARD, 1917)
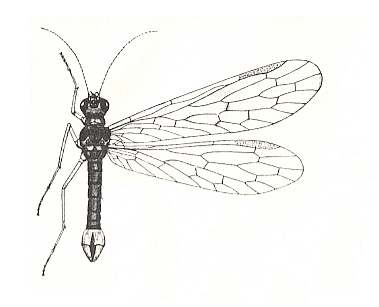
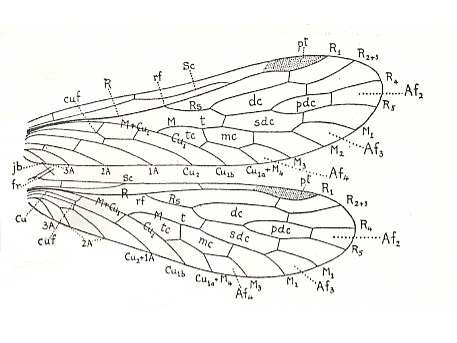
Figure 5 : Wing-venation of Nannochorista dipteroides TILL. (Order Mecoptera, Family Nannochoristidae), Comstock-Needham nomenclature and notation [which is still a generally accepted way of signifying venational details].
Af2, Af3, Af4, second, third, and fourth apical forks. cuf = cubital fork, dc = discoidal cell [not homologous with "discoidal cell" in Diptera], mc = median cell, pdc = post-discoidal cell, pt = pterostigma, rf = radial fork (origin of Rs), sdc = subdiscoidal cell, t = thyridium (medial fork), tc = thyridial cell.
(After TILLYARD, 1917)
If we look to the wing-venation (forewings) in Mecoptera and to that in Diptera, we see that the latter has wings in which the venation is strongly reduced as compared to that in the former. In the Nannochoristidae this originally very rich mecopterous wing-venation has been reduced already so far that we must depart from their venation (concentrating on forewings only) -- in addition to that of some other Mecoptera -- in order to understand and interpret the venation in the Order Diptera. And to understand in turn the venation in Nannochoristidae, we must go back to the more primitive wing-venation as we see it in the large mass of fossil representatives of the Mecoptera.
Having aquainted ourselves a bit with recent Mecoptera (of which the Nannochoristidae -- as has been said -- are the most interesting with respect to the origin and interpretation of dipterous wing-venation), we will now give a number of f o s s i l representatives of the order Mecoptera (most of which are only known by the fore wing). The intention of presenting this series of Figures is to show the degree of (venational) diversity -- but still resting on a more or less (venational) uniform framework -- of Mecoptera especially in Permian times. In all this we assume that venational diversity, respectively venational uniformity, express the diversity, respectively uniformity, of the insects as a whole (in fossils, in the overwhelming majority of cases, the wing-venation of the insects is all we have).
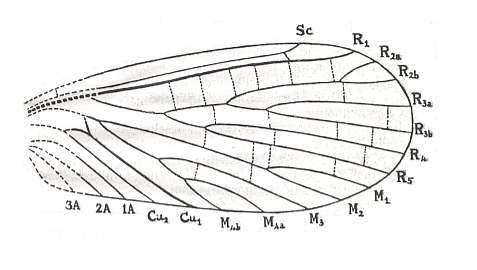
Figure 7 : Wing of Metropator pusillus HANDL. ( Lower Upper Carboniferous (Lower Pottsville), U.S.A.). Length 7.6 mm. Drawn from Specimen No. 35382 in U.S. National Museum, Washington, Department of Paleontology.
(After TILLYARD, 1926)
If the interpretation of the wing-venation of Metropator (Figure 7) is correct, then we can say that Mecoptera were already there in Upper Carboniferous times. This single fossil is the only mecopteron so far found in sediments older than those of the Permian.
CARPENTER (1965) has re-examined the fossil described as Metropator and came to different concusions [but not at all definitive]. See next Figure and and exposition :
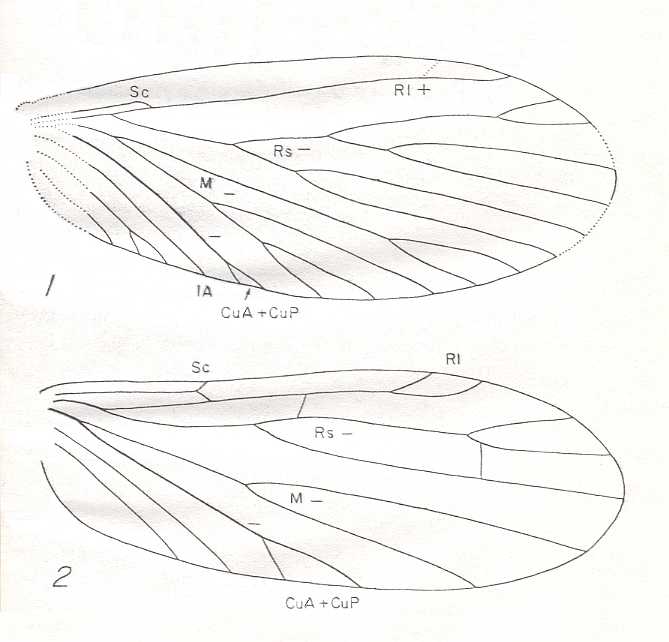
Figure 7a : Miomoptera [according to CARPENTER].
1 - Metropator pusillus HANDLIRSCH. Drawing of holotype, nr. 38731, U.S.N.M. Hindwing [according to CARPENTER]. Length 7 mm. Namurian (lower part of the Upper Carboniferous) of Pennsylvania, U.S.A.
2 - Palaeomantis minuta SELLARDS. Hind wing [according to CARPENTER]. Length 4.3 mm. Lower Permian of Kansas, U.S.A.
Sc = subcosta, R1 = radius, Rs = radial sector, M = media, CuA = anterior cubitus, CuP = posterior cubitus, 1A = first anal vein, + = convex veins, - = concave veins.
(After CARPENTER, 1965)
About Metropator pusillus HANDL. Carpenter, 1965, pp. 177, writes (among other things) the following :
Handlirsch, who originally placed Metropator in the Palaeodictyoptera [one of the most ancient insect orders], believed that the anterior margin of the wing was broken away, the front edge of the wing as preserved being the subcosta [that is, the subcostal vein]. He apparently reached that conclusion because he was unable to discern the subcosta as a submarginal vein. Tillyard in 1926 [see Figure 7, above ], following his examination of the type specimen, concluded that the anterior margin of the wing was actually preserved and that Sc was discernible as a distict vein between R1 and the wing margin. In his description he points out that the subcosta is very faintly indicated, and that he could follow it out only with care by examining the fossil in good oblique light. He also described and figured the cubito-median "Y-vein", this being much more strongly developed than most of the other veins of the wing. His conclusions were that Metropator was a mecopteron, closely related to the Permopanorpidae. He did not discuss the detailed evidence for this, but simply asserted that the mecopterous affinities could readily be seen at once from the figures. His view of the position of Metropator has been generally accepted subsequently, and it is the one presented in the Osnovy Paleontologii (Martynova, 1962).
The drawing included in figure 1 [see Figure 7, above, upper image ] represents my own interpretation of this fossil and shows only those structures which I confidently believe are present. From my studies I am convinced that Tillyard was correct in his conclusion that the costal margin of the wing is actually present in the fossil, but I am also convinced that he was incorrect in his interpretation of the subcostal and cubital areas. The subcosta is discerible near the base of the wing, as noted by Tillyard, but that is the entire length of the vein. It extends only a short distance beyond the origin of Rs. The ammonium chloride preparation brings this vein out clearly enough so that it is visible in photographs. On the other hand, the supposed branch of the media, which Tillyard shows as one arm of the cubital-median "Y-vein" [see Figure 7, above ] and which was an important factor in his conclusions on the affinities of the fossil, cannot be seen -- at least not by any techniques used by me. The cubital vein itself (labelled Cu1 in Tillyard's drawing) is, as mentioned by Tillyard, a distinct one, which stands out more strongly than any of the others excepting R1. It is, however, clearly concave [that is, lying in a groove, and thus not on a ridge] in the fossil. This is important, since the supposedly homologous vein (CuA) in the mecopterous wings is strongly convex. I am convinced, therefore, that the venation of Metropator only superficially resembles that of the Mecoptera and that it does not have the essential features of the mecopterous venation.
I believe the type-specimen of M. pusillus can much more readily be interpreted as a hind wing of a miomopteron [that is, the order Miomoptera, established by Martynov fore a number of small fossil insects apparently related to the Protorthoptera (fossil insects themselves related to the orders centered around grass-hoppers and their relatives) ] In these wings the subcosta is very short [see Figure 7, above, bottom image ], Rs arises close to the base of the wing, and CuA and CuP are completely coalesced, forming a strong concave vein. These are the outstanding features of pusillus. Unfortunately, since the base of the wing is missing in the type of pusillus, the precise relationship between Cu and M cannot be determined. However, there is no reason to assume that M does not join Cu near the basal part of the wing. The venation of pusillus shows more extensive branching than in the miomopteron illustrated in Figure 7, above, bottom image (Palaeomantis minuta SELLARDS), but in other genera of Miomoptera [...] the radial sector has more branches than in Palaeomantis. It seems to me, therefore, that the available evidence, such as it is, indicates that Metropator is more likely a miomopteron than a mecopteron. The occurrence of several genera of Miomoptera in the Carboniferous deposits of Europe supports this probability. The Mecoptera, on the other hand, are otherwise unknown from beds earlier than the Permian, and since these are endopterygote (that is, holometabolous) insects, evidence for their presence in the lowest strata of the Upper Carboniferous should be really convincing before such a conclusion is reached. At present I believe the evidence points to a very different conclusion.
( CARPENTER, F.M., Studies on North American Carboniferous Insects, 4. The genera Metropator, Eubleptus, Hapaloptera, and Hadentomum, published in : Psyche, June 1965, p. 175-190 )
That the taxonomical interpretation of many fossil insects is difficult is evident in this discussion of Carpenter. Indeed, the wing depicted in Figure 7, above, bottom image , and named Palaeomantis minuta SELLARDS, was earlier (1932-33) interpreted by Carpenter as belonging to the order Psocoptera, and belonging to the genus Delopterum. So the taxonomical status of Metropator remains debatable. We must acknowledge the possibility that is is a mecopteron and that consequently holometabolous insect were already present in the Carboniferous.
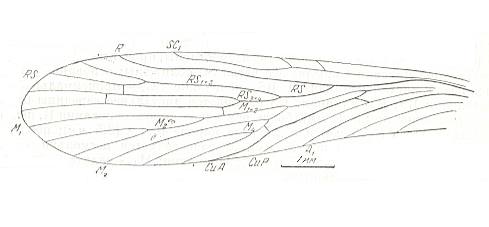
Figure 8 : Fore wing of Mesochorista novojilovi O.MART. (Mecoptera). Coll. PIN No. 673/306. Length of impression 8.5 mm, probable length of whole wing 8.8 mm. Sokolova II. Permian of Basin of Kuznetsk, Siberia.
Description of venation in : Paleozoiskije Nasjekomije Kuznjetskovo Bassjeina [Paleozoic Insects of the Basin of Kuznetsk (Siberia)], ROHDENDORF, et al., 1961.
The wing was found in the Jerynakovskaja suite of the Basin of Kuznetsk (p. 583). The geological age is given (p. 11) as P2, which should then be upper Permian.
(Figure after Martinova, in ROHDENDORF et al., 1961)
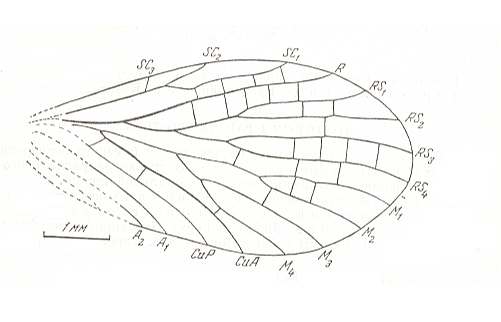
Figure 9 : Fore wing of Tychtopsyche beljanini O.MART. (Mecoptera). Coll. PIN No. 506/101. Sokolova II. Upper Permian of Basin of Kuznetsk, Siberia.
(After Martinova, in RODENDORF et al., 1961)
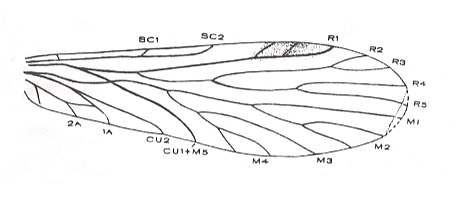
Figure 10 : Fore wing of Lithopanorpa pusilla TILL. Length 4.0 mm. (Mecoptera). Lower Permian of Kansas, U.S.A.
(After CARPENTER, 1930)

Figure 11 : Photograph of (fossil) wing of Lithopanorpa pusilla TILL. No. 5066a, Peabody Museum. (ca. x 21). (Mecoptera). Lower Permian of Kansas, U.S.A.
(After CARPENTER, 1930)
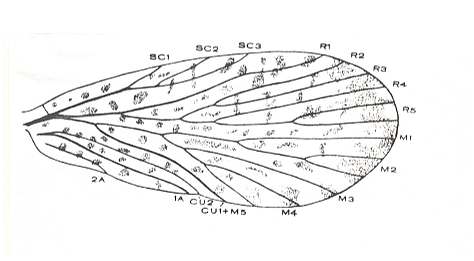
Figure 12 : Fore (?) wing of Agetopanorpa maculata CARP. Length 9.0 mm. Lower Permian of Kansas, U.S.A.
(After CARPENTER, 1930)
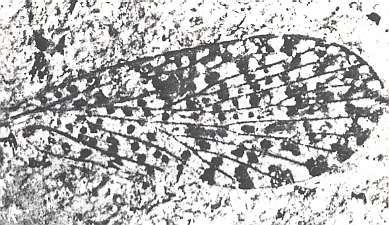
Figure 13 : Photograph of (fossil) wing of Agetopanorpa maculata CARP. (ca. x 13). Lower Permian of Kansas, U.S.A. No. 3037a, Museum of Comparative Zoology.
(After CARPENTER, 1930)
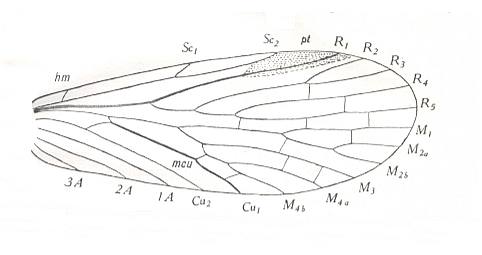
Figure 14 : Forewing of Permochorista sinuata TILL. Order Mecoptera, Suborder Eumecoptera, Family Permochoristidae. Upper Permian of New South Wales, Australia.
(After TILLYARD, 1935)
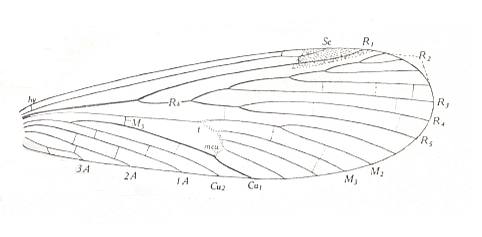
Figure 15 : Forewing of Orthophlebia liassica MANT. Order Mecoptera, Suborder Eumecoptera, Family Orthophlebiidae. Lower Lias of England [the Lias is the oldest division of the Jurassic].
hv = humeral veinlet, M5 = upper arm of cubito-median Y-vein, mcu = medio-cubital cross-vein, t = thyridium [in about the middle of the wing].
[The true primary fork of the Media lies in fact where M5 branches off from the main stem of the Media. It is not always evolutionarily preserved. Therefore we can conveniently call the next furcation point, namely where M4 branches off from the main stem of M, the primary fork of the Media, because this fork is in most cases evolutionarily preseved. This primary fork is called the thyridium (t). Often the venation in the immediate vicinity of the thyridium is weakly chitinized. We can then speak of a thyridial area.]
(Figure after TILLYARD, 1935)
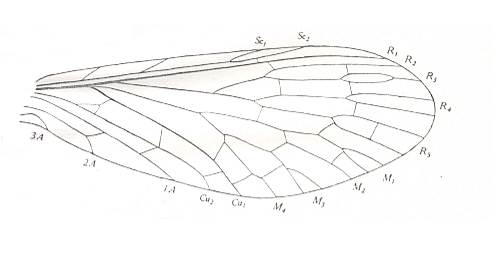
Figure 16 : Forewing of Parabelmontia permiana TILL., Order Mecoptera, Suborder Paramecoptera, Family Belmontiidae. Upper Permian of New South Wales, Australia. (After TILLYARD, 1935)
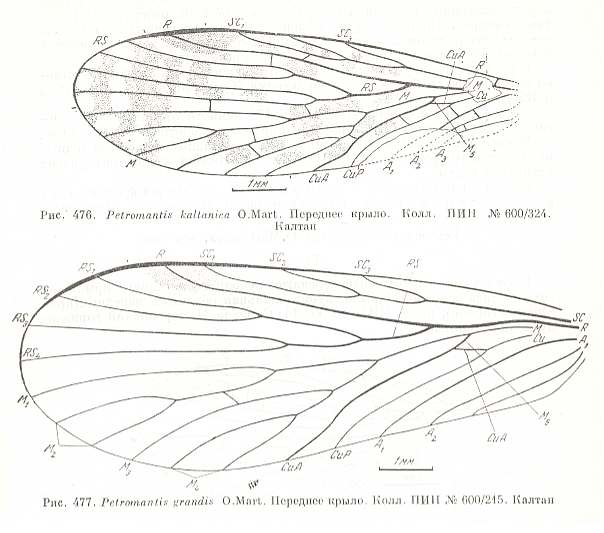
Figure 17 :
Upper image : Forewing of Petromantis kaltanica O.MART. Coll. PIN. No. 600/324. Length of impression 8.2 mm, probable length of whole wing 9 mm. Kaltan, Lower Permian of Basin of Kuznetsk.
Bottom image : Forewing of Petromantis grandis O.MART. Coll. PIN. No. 600/215. Length 10.7 mm. Kaltan, Lower Permian of Basin of Kuznetsk, Siberia.
(After MARTINOVA, in ROHDENDORF et al., 1961)
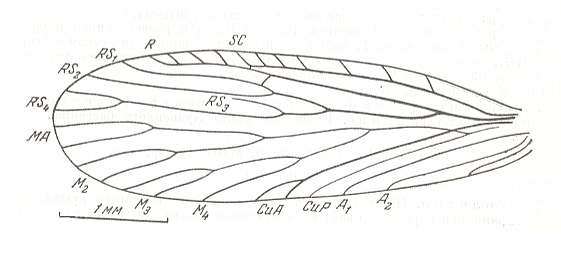
Figure 18 : Forewing of Tomiochorista minuta O.MART. Coll. PIN No. 600/194. Length 4.5 mm. Kaltan, Lower Permian of Basin of Kuznetsk, Siberia.
(After MARTINOVA, in ROHDENDORF et al., 1961)
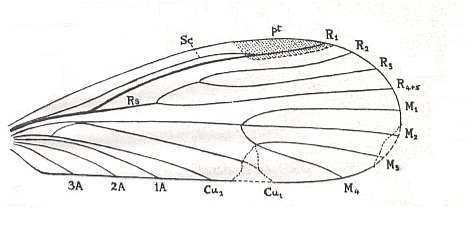
Figure 19 : Wing of Anormochorista oligoclada TILL. Specimen No. 5068 (reverse) in Yale University Collection. Length 5 mm. Lower Permian of Kansas U.S.A.
(After TILLYARD, 1926)
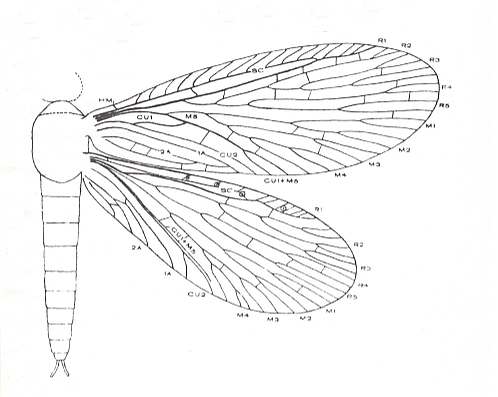
Figure 20 : Platychorista venosa TILL., female.
Length of insect (excluding head) 3.6 mm. Length of forewing 5.6 mm.
Lower Permian of Kansas, U.S.A.
(After CARPENTER, 1930)
From the above Figures it should be clear that the Permian Mecoptera already as such, that is as typical Mecoptera (i.e. still not yet having evolved into Paratrichoptera), show a significant diversity in their (fore) wings (shape and venation), while their venational diversity still rests on a common venational scheme. So this situation is already such that it does not a priori exclude the possibility of the polyphyletic origin of the Diptera. This conclusion will be further confirmed when we study those fossil Mecoptera that are placed into the suborder Paratrichoptera. This will be done in the next document.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XV.