
As was shown above, the new, and comparatively more perfect work-order of wasps, having originated in complexificated circumstances during its establisment, still was encumbered by an essential insufficiency, namely the fact that to the place of the timely prepared nest had to be brought a prey taken in the random conditions of the hunt, and moreover being sufficiently large. As for this difficulty a way out was found. This could be accomplished thanks to the fact that the construction of the nest before supplying provisions, itself served as a base for further, and indeed for the core transformation of the providing-instinct. The fact is that precisely now the possibility was there to place a stock of provisions in the timely prepared cell, not in one definitive amount, as this took place until now, but in several, that is, supplying in measures. First, instead of the cell being provided with one rather big prey, two were brought in [one after the other] that were, however, of smaller size. See next two Figures.

In all this, the amount of labour of transportation of the prey is decreased, especially when preys are not at hand but must be taken from more or less distant places.
The work-order, in all this, now became a little different [from what it was before], and it may be expressed, using our symbols [For the meaning of the symbols, see HERE (previous document) ] thus :
( preparing a nest, catching and paralyzing a prey, transporting the prey to and into the nest, oviposition on that prey, catching and paralyzing a second prey, transporting it to and into the [same] nest [and also the same cell], closure of the nest) (explanation follows)
In order to understand this consecutivity of acts, let us try to base things on the same principle as before (previous document). Let us suppose, consequently, that also in the present case cancellation of individual links from the ladder of serial actions took place followed by unification of the remaining links without transposition in time. Then we get [where "0" = zero] :
first series (B + A1 + C1 + D1 + 0) + second series (0 + A2 + C2 + 0 + E2) =
B + A1 + C1 + D1 + A2 + C2 + E2.
[ Originally, the individual series each belong to some one individual nest (or cell) built by the same individual wasp :
A, B, C, etc. originally belong to the first nest.
A1, B1, C1, etc. originally belong to the second nest.
A2, B2, C2, etc. originally belong to the third nest.
A3, B3, C3, etc. originally belong to the fourth nest.
etc.
The "first series", (B + A1 + C1 + D1 + 0), as here signified just above, thus is in fact a combination of the original first series and the original second series, while the "second series", (0 + A2 + C2 + 0 + E2), as here signified, belongs to the original third series.]
Let us now, to explain further, turn to the true activity. The digger wasp Ammophila urnaria Gress., according to observations by PECKHAM, 1898, usually places two (sometimes one) caterpillars into the cell, whereby it temporarily closes the nest two times -- first after having prepared the cell, and then after oviposition onto the first prey (on the second prey no egg is laid). Consequently, the complete work to provide one larva may in this case be written as :
(0 + B + 0 + 0 + E) + (A1 + B1 + C1 + D1 + E1) + (A2 + B2 + C2 + 0 + E2).
(Where E is the temporary closure of the nest (applying a cap), as is also E1, while E2 is the definitive closure of the nest. Further, B1 is, as B, a building (digging) activity, here, however, not the building of the nest (as is B), but the removal of the temporary cap of it. And the same goes for B2.)
The temporary closure (E and E1) of the cell and the removal of this temporary cap (B1 and B2) together make zero, and therefore, having subtracted E, E1, B1, B2, as elements that cancel each other, we will obtain the results of the work of Ammophila urnaria :
which is what we sought for (see above ).
Apparently, the present method (way) of complexification of instincts (by way of cancellation followed by combination) also was taken further, proportionally to the growth of the number of preys in a cell in connection with the decrease of their size. At least there exists an indication, although insufficiently precise, that Ammophila yarrowii Gress. and A. polita Gress., bringing into a cell several (4-6) caterpillars, each time close the entrance of the nest. In the same way behave the small individuals of A. procera Dahlb., and partially also A. campestris Latr.
Temporary closure of the nest during providing is somtimes also done in other digger wasps. However this may be, the temporary closure and opening of the nest may, in certain circumstances, become superfluous and then totally disappear. Thus, A. holosericea L., which also provides for several (up to 5) preys, already does not temporarily close the nest. The cancellation of these superfluous acts, easing the work of the wasp, cooperated to further increase of the number of preys in a cell [per cell] with, of course, a corresponding decrease of their size. Moreover, the uniform repetition of a given act may create a certain inertia, which works in the same direction [i.e. contributes to the persistent cancellation of superfluous acts].
Now, this repetition, we have here, a repetition, that is, without substantial change of actions carried out at the previous moment, undoubtedly also was of great significance to the working-out of maternal instincts in wasps and bees. The most characteristic examples of such "mass-providing" of each cell with a number of similar [among each other] preys is encountered in the majority of solitary wasps (especially of the families Crabronidae, Oxybelidae, Trypoxylonidae, Mellinidae, Nyssonidae, Pemphredonidae (see next Figure), and also of the plaited-winged wasps (Eumenidae)).

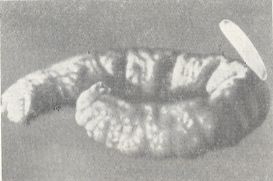
Figure 3 : Nest of Psenulus fuscipennis Dahlb. Pz. The cells are filled with aphids.
1 - a just emerged larva of the wasp. 2 - separation wall. (After MALYSHEV, 1966)
Taking into account the indissoluble consecutivity -- in such a provision -- of two acts, namely the hunt (A) and the bringing-in of the prey (C) into the cell, the previous series may be given by :
or, in a more general form :
(indices as to what series the individual acts belong, now omitted)
where (AC)n signifies the hunting acts corresponding to the number of preys (n) to be prepared after oviposition (D). So the formula means : building of the nest, catching and bringing-in the first prey, oviposition onto this first prey, repeated catching and bringin-in of additional preys [into the same cell] (without oviposition), closure of the nest.
In solitary wasps, in an increased rate preparing for several preys in one single cell, sometimes also another complexification is observed. It consists of the fact that the egg is deposited, not onto the first prey that has been brought in, but onto one of the next preys or even onto the last one, which [latter] may be espressed as :
In this case, where we have to do with a high rate of provision, in these wasps not interrupted by another activity (temporary closure and opening of the cell, periods of awaiting), the act of oviposition looses its connection with the first-brought-in prey, and now comes only at the completion of the provision such that the closure of the cell follows directly upon oviposition. So do behave, for example, the American species of Sceliphron and Podium. For a Palaearctic representative of the genus Sceliphron, see next Figure.
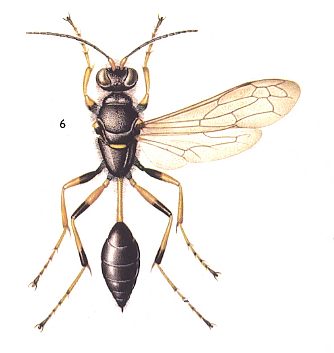
Figure 4 : Sceliphron destillatorium. 15-30 mm. (Europe, Central-Asia, Iran). Family Sphecidae.
(After SEVERA, in Thieme's Insektengids voor West- en Midden-Europa, 1977)
Parallel to the increase of the number of preys to be prepared in a cell, the qualitative selection of the provision also changed. If we survey the preys of he wasps of the present group [these wasps] bringing into each cell several preys, then we are struck by the unusually wide qualitative content of the [collection of] preys, far surpassing all what is observed in the previous Phases of the evolution of wasps [That is, the range of possible preys (prey-types) is much larger than it is in the previous Phases, where specialization with respect to prey-type is much more the rule].
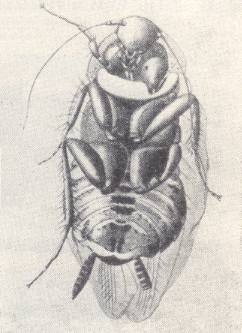
Figure 5 : Egg of Tachysphex lativalvis Thoms. deposited transversely on the thorax of the cockroach Ectobius lividus. (After GRANDI, 1929, in MALYSHEV, 1966)
Other species (Tachysphex mantiraptor Fert., T. julliani Kohl.) hunt for young praying mantises (mantids), that is, for representatives of an insect Order (Mantoptera) close to that of the cockroaches (Order Blattoptera) [in IMMS' Texbook of General Entomology, 1977, the cockroaches and the mantids are placed in one and the same insect Order, the Dictyoptera]. In one cell are placed 3-16 paralyzed mantids, and the egg is deposited upon one of the last of them or upon the last mantid having brought into the cell. The long body of the mantid does not completely fit into the cell of Tachysphex and projects from it out into the nest channel. Such non-correspondence of the sizes of the cell and the prey tells us, apparently, that the building instinct here in its development lagged behind the selection of prey, and that this cell once contained another prey having a normal size such as cockroaches. For mantids also hunts Stizus ruficornis F. (Stizidae).

Figure 6 : Content of a cell of the wasp Dinetus pictus F.
Left : Five paralyzed bugs (Hemiptera) Reduviolus ferus L. with an egg of the wasp on one of them -- onto the middle one.
Right : The bugs, taken out of the cell.
(After MALYSHEV, 1966)
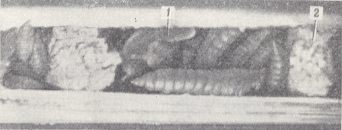
Figure 7 : Nest of Spilomena troglodites Lind. made in a stalk. In the cells are provisions consisting of paralyzed thrips.
1 - egg of wasp. 2 - dividing wall. (After MALYSHEV, 1966)
As to earwigs (Dermaptera), stick- and leaf-insects (Phasmoptera [Phasmida]), termites (Isoptera), and some others,-- they have, apparently, not been encountered on the paths of evolution of solitary wasps, and therefore are also not encountered in them today as preys. On the other hand, spiders are taken as preys also in the present Phase, but are represented by smaller forms, and especially [by] cross spiders (Epeiridae).

Figure 8 : "Cuckoo's spit" -- a foamy mass, formed on a stalk of an euphorbia by nymphs of the cicada Aphrophora sp. (After MALYSHEV, 1966)
In order to pull out such an, apparently, strange, not directly visible, prey from the foamy mass surrounding it, the wasp has to penetrate into the foam not only with its sting, but also with its legs.
Similar things tell us FERTON,1901, and GRANDI,1926, about Gorytes. According to REINHARD, 1925, Hoplisus costalis Gress. adheres its egg to the thorax of a cicada (also of the Membracidae) only with one end, but which one [head or tail end] is not indicated.
On the 2nd of August, 1949, the author [Malyshev] succeeded in finding a nest of Harpactus [subgenus of Gorytes] tumidus in the Chopersky nature reserve. In the just provided [with prey] cell there were 11 cicadas present -- Jassidae -- of which 5 adults and 6 nymphs. The egg of the Gorytes was present on precisely that adult cicada that had been placed in the very depth of the cell, and brought in, we should think, as the first prey. See next Figure.

Figure 9 : Egg of the gorytes Harpactus tumidus Pz. on the cicada-jassid (Jassidae), taken from the cell of the wasp. (After MALYSHEV, 1966)
It had a length of 1.5 mm and rested along the ventral side of the cicada, beside its left legs. The egg's wider end was directed anteriorly, to the head of the cicada, and its narrower end was directed tailward. By reason of the fact that in other wasps (in Bembex'es (Sphecoidea) and Stizus'es (Stizidae)) the narrower end of the egg corresponds to the head-end of their larva, we can assume the same relationship holding also here (Harpactus) [That is, in the case of Harpactus the narrow end of the egg would also correspond to the head-end of the larva in that egg, and that, consequently, this head-end is directed toward the posterior end of the cicada].
But most remarkable in the present Phase is the wasps hunting for adult insects with complete metamorphosis (Holometabola) -- flies (see next Figure), midges, beetles (next next Figure), butterflies, bees, wasps, ants, terebrants [= parasitic wasps], and others.
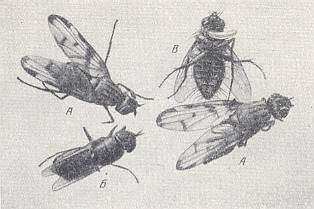

Figure 10 : Content of a cell of the wasp Crabro chrysostomus Lep.
A - Two fruit-flies Otites formosa Pz.
6 - One stratiomyid fly Eulalia hydroleon L.
B - One tachinid [fly, as larva endoparasitic] Dexiomorpha sp. with an egg of the wasp.
Right image - Fruitfly. The egg of the wasp is visible. It is adhered with its head-end toward the throat (of the prey).
(After MALYSHEV, 1966)

Figure 11 : Content of a cell of Cerceris quinquefasciatus consisting of 29 small leaf-eating beetles (Chrysomelidae) and weevils (Curculionidae). (After MALYSHEV, 1966)
The characteristic attachment ('devotion') of these or those wasps to one fairly determined [specific] prey, is striking. Here, specialization within the limits of the species usually goes together with a significant variability within the limits of the genus : Different species of one and the same genus of wasps not seldom select a prey very different from the preys of the other species of its genus, and typical to themselves. Here, however, not only individual species, but also whole genera, and even groups of related genera, often show an attachment to preys of one determined group (for instance, to orthoptera), and this, not in connection with mere local [and more or less the same] conditions [i.e. this attachment to preys belonging to one determined group, is not some local (and thus locally caused) phenomenon], but, the other way around, in [connection with] different conditions even in different continents [i.e. despite different conditions and even different continents, the mentiond phenomenon prevails]. Thus, the Tripoxylonidae, and Sceliphron's everywhere hunt for small spiders, and the Ammophila's [everywhere] for bald caterpillars.
The notification (SMIRNOV, 1915) that Ammophila (Eremochares) dives Brullé selects grasshoppers (and not caterpillars or pseudo-caterpillars [larvae of saw-flies] ) is, apparently, incorrect. This also is the opinion of BERLAND, 1925.
The second possible evolutionary path along which was realized the change in prey selection consisted of the fact that one and the same wasp began to carry to its cell very different [from each other] preys. Thus it is known that Palarus flavipes F. simultaneously collects different genera of terebrants, waps, and bees (Ichneumon, Tiphia, Myzine, Mutilla, Scolia, Cerceris, Philanthus, Lyrops, Crocia, Sphecodes, Ammobates, Andrena, Apis). There are, however, no indications that later these wasps might specialize on some one of the preys taken initially.
As we will see later, this is still not the limit of possible selection of prey by the solitary wasps. We get the impression that the instinct of the insect, having reached a certain high level of development, enters, as it were, a special pliable condition [of it], easing further possibility of its evolution.
Summarizing what has been expounded, we see, on the one hand, together with the great effort needed to drag [it] over the ground, the very large but single [one and only] prey, being many times (15 times in Psammophila hirsuta Kby. and Ammophila sabulosa L., according to FABRE) heavier than the wasp having attacked it, and on the other hand [we see] whole tens of small forms [as prey] (aphids, small caterpillars, ants), which are carried to the cell by air with unsurpassible speed and ease. In this way the instinct has coped with the weight of the prey. As a result, the preparation of provisions, [evolutionarily] earlier being simple, consisting of one single act, now has become to consist of many acts [repeatedly catching and bringing in (small) prey]. In this state the provisioning indeed is found to be in all higher wasps and in all bees nesting independently.
In the present[ly under discussion] Phase of [hymenopterous] evolution, in addition to the paralyzation of the prey up to this or that measure, and also to the overpowering of it with its sting or jaws, from time to time maiming of the prey is observed. From its prey the wasp bites off parts of one or several legs, and sometimes also the antennae, which [operation] increases the manageability of the prey and eases the placing of it in the cell. In this way processes, for example, Notogonia pomiliformes Pz. crickets, Sphex maxillosus F. -- katydids, Sphex albisectus Lep. -- grasshoppers. Sometimes of the prey, on that side of it where the egg of the wasp (Crabro, Oxybelus) is laid, the wasp turns out the wing [of the prey], preventing, apparently, the prey from falling over, and with it the egg, onto the bottom of the cell.
Regarding the "chewing" of the prey [by the wasp] at the region of its head or thorax, also this operation is sometimes observed in the present Phase, and in certain cases this act acquired here a special significance. Thus, in the American species of Diodontus the chewing of the prey (aphid), apparently, totally replaced paralyzing it with the sting.
With all this we conclude our exposition of the Third Vespine (Crabroid) Phase of hymenopterous evolution.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LI.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX