
Organic Evolution in terms of the Implicate and Explicate Orders.
Part LIII
Hymenoptera (wasps, bees, ants) (Sequel)
The evolutionary diversification in the Order Hymenoptera in terms of Strategies (Sequel).
Newly vespine (vespoid) phase
We here again present a picture of Eumenidae, which play an important part in the present Phase of evolution, and, together with some other wasps, eventually lead to social wasps.
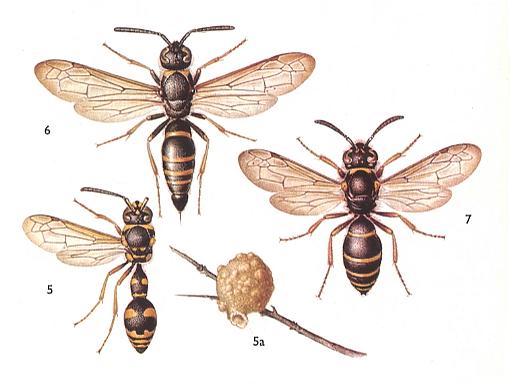
Figure 1 : Eumenidae.
5 : Eumenes pomiformis . 10-16 mm. 5a : nest.
6 : Ancystrocerus antilope . 12-15 mm. [ Ancystrocerus might (also) be a subgenus of the cosmopolitic genus Odynerus including a number of subgenera]
7 : Oplomerus spinipes . 9-12 mm.
(After SEVERA, in ZAHRADNIK, Thieme's insektengids voor West- en Midden-Europa, 1977)
REMARK on Eumenidae and other wasps intermediate between solitary and social wasps. The details of this Remark are taken from other sources (other than from MALYSHEV, 1966).
In the transition from solitary wasps to social wasps the family Eumenidae plays a significant role. The Eumenidae are solitary pleated-winged wasps. The family contains several genera, among which Eumenes and Odynerus. About the latter we might add the following : The 'old' genus Odynerus today is interpreted as representing a complex of several genera. It contains about 3000 known species, and is distributed all over the world.
The Eumenidae (Eumenes, Odynerus, Synagris, Zethus) are related to the Vespidae. The latter contains three subfamilies, viz., the Stenogastrinae, the Polistinae, and the Vespinae. The latter two are true social wasps. About the Stenogastrinae EVANS and EBERHARD, 1973, p. 159, 161, say the following :
The Stenogastrinae (Stenogaster and Parischnogaster), although classified in the family Vespidae along with the social wasps, have so far proven to be subsocial wasps, in many respects intermediate between the solitary Eumenids and the social Vespids. IWATA has described them as "rather dull and depressed [I take this to refer to their color and shape] wasps that live mostly in gloomy and wet habitats especially along mountain torrents and cascades." As in the Eumenids Synagris and Zethus, a single female [of Stenogastrinae] builds a cluster of cells, either of mud or of mud mixed with vegetable material, and places an egg loose in the bottom of each cell (rather than glued to the cell wall as in social vespids). The eggs of the Stenogastrinae are intermediate in average size between those of solitary wasps and the relatively small ones produced by social wasps. A food ball of viscid liquid, perhaps regurgitated from the mother's gut, is placed in the cell before the egg hatches. Later the larva is like a young social wasp in that it is progressively fed masticated insects and spiders. But unlike most social species the mother seals the nest of the mature larva and no cocoon is spun. The female generally attends several cells of various ages simultaneously, and is evidently sometimes still present when her first offspring emerge. Sometimes two or more females are found on the same cluster of cells, but nothing is known of their origins and activities. There is some evidence that offspring females sometimes remain near the parental cells and reuse them or build adjacent new ones. Unfortunately these oriental wasps have not received the study they deserve.
(End of Remark, continuing with Malyshev)
The slowed-down method of nursing of the larva, providing it with provisions day by day, implies in the Eumenidae the same insufficiency as was also seen in certain digger wasps, the bembexes. It decelarated, as we saw, all reproductive activity of the wasp. Under its pressure the wasp, instead of 1-2 days or even a few hours, now spends 2-3 weeks and more in nursing only one larva. Spending so much time is possible only in a very favorable climate, especially there where the activity of the wasp is possible the whole year around, as for instance in tropical Africa.
Precisely in such conditions, in the wasp Synagris cornuta L., related to the odynerus, about one month goes by from the moment of oviposition up until the closure of the nest. In some cases, however, this wasp may nurse one single larva during more than three months (ROUBAUD, 1910). It is unsurprising that also in these optimal conditions S. cornuta is capable of rearing only 4-5, maximally 6, descendants. Such slowed-down nursing of larvae, one after the other, supposes a long life of the mother-wasp herself. Unfortunately, we do not have direct data about that.
Individual elements of the new instincts are, still in their initial stages, encountered in various representatives of the Eumenidae. But an integration (combination) of [a number of] these elements was needed before the beautiful effect could ensue.
Feeding one larva from day to day with the ease of catching and transporting small provisions afforded the wasp much extra time, that was spent by it in an unexpected way : Instead of nursing one larva after another, which is generally observed in solitary species, the wasp Odynerus tropicalis Sauss. sets itself to nurse the second larva, before having finished provisioning the first. The very construction of the cells proceeds here, apparently, consecutively, but the care for several (at least two) larvae coincides in time [That is, the larvae are not reared one after the other, but reared at the same time]. This also formed the beginning of simultaneous nursing of several larvae by one individual -- the first step down the path of development of the new structure of vespine life.
A fundamental hindrance for the evolved new instinct was the old, totally primitive construction of the cells (as in O. tropicalis) in the form of short, nearby galleries, dug out into the thickness of earth walls. But we can see that the building instinct in Eumenidae freed itself form the limiting properies of the substrate, and the cells obtained a free shape (in the form of a glass, cup, and the like, as, for instance in Synagris cornuta L.) independent of the shape of the cavity taken by these cells. See next Figure.

Figure 2 : The wasp Synagris cornuta L., at its nest, modelled from white clay.
(After BEQUAERT, 1918, in MALYSHEV, 1966)
In addition to the perfection of the shape of the cell we see in Eumenidae also an evolution of the [use of] building material : Instead of the little perspective heavy mineral substance (earth, clay) being originally used, an unusual light plant material came into use -- "wasp paper", having beautifully guaranteed, in this respect, the bloom of vespine life. It is remarkable that true "wasp paper" with its grey color is already encountered in the odynerus -- Odynerus (Symmorphus) murarius L., although the paper is used here only for a special purpose -- for closing the entrance to the finished nest. See next Figure.

Figure 3 : The wasp Odynerus murarius L., closes its nest (in a log) with "wasp paper".
(After MALYSHEV, 1911, in MALYSHEV, 1966)
We just saw the destruction of the original consecutiveness in the work of the wasp, expressed by the simultaneous provisioning of two cells (O. tropicalis). But among Eumenidae also another destruction of the same elementary consecutiveness in the series of acts is observed. This second destruction relates to the building activity of the wasps and is expressed in the partial [= still incomplete] construction of cells, or, more precisely said, in the interruption of a whole, until now, building act and insertion into it of the following act, namely oviposition. Thus, the Brazilian wasp Zethus lobulus Sauss., related to the Eumenidae, after having built its free cell only half way, oviposits in it, and then finishes construction and passes over to provision the cell.
All these individual changes of the nursing and building activity of the wasps later [in evolution] began to combine with each other and became the basis for the fundamental change of the maternal instincts of the pleated-winged wasps. Such a transformation took place when to the indicated basic stages still another, and evidently, the most important, element was added -- the bringing in (into the cell) of prey in a chewed condition. It is interesting that also this moment is encountered in Eumenidae in a fairly determined form. Thus, the above mentioned African wasp Synagris cornuta (L.) feeds its larvae not with whole caterpillars, but with preliminarily chewed ones.
Such a deep-seated change of habit of the wasp could originate only after a development of a basic instinct of the previous phase, namely oviposition in a still empty cell. In this case it is not possible to suppose a preliminary preparation of a chewed prey, onto which later an egg is laid. Such a kind of food stocks we do not know of in cells of wasps -- yes, they are hardly possible. From here it is clear that another preparational step in the new method of provisioning cells was the highly developed instinct of slowed-down provisioning in step with the needs of the larva. It is therefore not surprising that precisely in S. cornuta the method of slowed-down provisioning coincides als with the feeding of the larva with chewed food. For this also was fortunate the nature of the prey of such wasps : minute caterpillars (of Hesperidae) with delicate integument. We should also give attention to the relative sizes of the wasp and its prey. About this we do not possess precise data, but, apparently Synagris cornuta and relatives select, as compared to their own size, the smallest prey of all other solitary wasps. While S. colida L. and S. sicheliana Sauss. prepare more than 50 small caterpillars (for a large part also Hesperidae) for each larva, Synagris cornuta must bring them in in still larger numbers. To this one might conclude judging the size of the piece of kneaded caterpillar, offered by it to the [virtually] fully-grown larva. See next Figure.
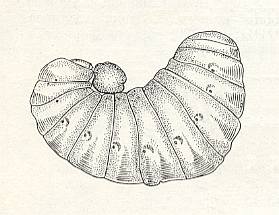
Figure 4 : Almost fully-grown larva of Synagris cornuta L., with a piece of provisions from a chewed caterpillar. (After ROUBAUD, 1910, in MALYSHEV, 1966)
So far, indeed, these wasps have evolved away from the original stage where only one prey was needed to feed one larva.
The point of departure of this act is interesting. The tearing up of the prey for their own feeding has, in large saw-flies, as was told, a deviating nature and has no relation with the feeding of the wasps [themselves]. The "sucking" of saps of the prey at the spot of the sting of the ovipositor, distributed among terebrants [parasitic hymenoptera] is, evidently, their specific trait and is also not characteristic of [true] wasps (aside from some Bethylidae). In the same way also "maiming" of the prey (amputation of limbs, interfering with transportation and placing of the prey in too narrow a cell) stands in no direct connection with feeding neither of larvae nor of adult wasps. The source of [the habit of] feeding the larvae with chewed bits of prey, lies, presumably, in another supplementary operation of the wasps -- in "malaxation" ("chewing") of the prey, already spoken of above. Although initially malaxation of the prey was not connected with inflicting wounds and the sucking of saps, with the delicateness of the integument of small caterpillars injury could easily lead to licking-up by the wasp of saps oozing out. It is interesting that a similar kind of tearing up by the jaws of the wasp of a delicate region (caudal projection) of the body of the prey gave in Odynerus nidulator Sauss. rise to special habits and led to supplementary feeding of the wasp itself. Similar things are also observed in O. nobilis Sauss.
We saw that Eumenidae did not evolve along the line of hunting for a diversity of prey, as did bembex-formed wasps, but concentrated on collecting mainly small caterpillars. Supplementing, by "chewing", the light paralyzation of caterpillars, they formed the beginning of the habit of feeding the larvae with chewed bits of prey. But, oviposition in a still empty cell and slowed-down provisioning (progressive provisioning) were, as told, necessary prerequisites for this. Indeed, preparation of chewed bits of food could not be done en masse, but only gradually, in step with the growth and need of the larva. Progressive nursing of larvae with chewed bits of prey, and being given to them in a mouth-to-mouth fashion, finally led to the social wasps and became among them a wide-spread almost exclusive application.
When the prey (small caterpillars and the like) was brought still in one piece to the larva, the orientation of the cell was of no importance neither to the larva nor to the wasp. The position of the cell not only could be horizontal, but could also be with the opening upward, as is observed in Eumenes pomiformes F., and others. The position of the cell did not change the way of suspending the egg namely from the ceiling of the cell. Things changed when the food became offered as chewed bits. In a standing cell (as is often the case in eumenids) the larva, holding itself from the ceiling [and the entrance of the cell more or less obliquely directed upwards], would have its head downwardly directed, that is, toward the side opposite of the place from where food is offered to it directly into the mouth. This difficulty was removed by the fact that later [in evolution] the wasps began to orient their cells in a turned-over fashion, with the bottom upward, from where their larvae were suspended head-down, into the direction of the entrance of the cell. Thanks to this new orientation, debris, left over from feeding, automatically falls out of the cell. Matters were perfected by the fact that the social wasps totally "forgot" about sealing the cells built by them. The act [of the adult wasps] of the making of a cap of the cell was completely atrophied in them (i.e. it was cancelled) and passed over to their larvae that, upon conclusion of feeding, and having reached a fully-grown condition, themselves seal the cell when making a cocoon.
The transition to feeding the larvae with chewed bits of food itself further led to a fundamental change in the selection of prey. Instead of small caterpillars the wasps now began to chew various insects, especially those that are abundantly around, such as houseflies and even hornets (Vespa crabro L.), and honeybees. Having grabbed such a prey, the wasp quickly bites off robust uneatable parts (wings, legs, head), makes, at the same place, the rest into a cutlet, which it brings to the nest and gives to the larva. See next Figure.

Figure 5 : The hornet Vespa crabro L., having its head bent toward the cell, in order to feed the larva in it [going from cell to cell]. (After MALYSHEV, 1966)
We must assume, however, that just emerged, and also very young larvae, are not able to take such food [Only when the larva is older it can eat these bits of food]. Therefore, the wasp began to feed them with its saliva or with the sap of the prey, which it itself had licked up before, and which now must be given from the mouth of the adult wasp into the minute mouth of the newly-born larva. This is not a crude putting-in of a bit of food into the throat, as do birds, but a very delicate procedure, more likened to "licking up of nectar".
Subsequently, the instinct of feeding the larva with chewed bits of food, changed such that the wasp-mother began to accept all potential nutrients, even prepared hamburger meat, and also fish flesh, for feeding the larva. It is interesting that the adult wasps, even in these circumstances, did not become themselves carnivorous, but retained their original vegetarianism -- nectar of flowers and other sugar-containing substances. In these new conditions, with constant contact between adults on the one hand, and their larvae on the other, a special method of feeding developed in wasps -- trophallaxis, or mutual exchange of food between adults and larvae. The latter, before or after taking food, give off saliva which is eagerly swallowed by the feeding individuals. This kind of relationship not only guaranteed and regulates, but also stimulates the feeding of the larvae such that now we cannot speak anymore of any slowed-down provisioning [because now feeding the larvae (by the adult) is more active and direct], although the feeding of the larvae continues from day to day.
As to the role of trophallaxis in sociality, EVANS and EBERHARD, 1973, p. 138/9, say : " Trophallaxis should not be considered in any sense a "prerequisite" for insect sociality, as the earlier workers implied. For some social insects -- the stingless meliponine bees -- manage their social lives without any larva-adult interaction, sealing the egg and a food supply in a waxen cell that remains closed until the young bee matures." [We should add : Trophallaxis is not indispensable (not a sine qua non) to sociality, but it could have played an important role in its origin (and then, later, be secondarily abolished in certain groups.]
As soon as the nursing of the larvae from day to day with chewed bits of food combined with simultaneous construction of a number of cells and with the simultaneous nursing in them of a number of larvae, the reproduction of the wasps, in the beginning having been slowed-down, increased rapidly. Matters went faster still when, to isolate the larvae -- living now in free, turned upside down, and unified in combs, cells, -- from unfavorable external influences, the wasps began to built around the nest something like a climatic insulation -- a special insulating shell made of several concentric layers of "wasp paper" with a multitude of individual air cavities between these layers. As a result, from a single wasp-mother, initially hardly able to raise one larva in the course of several weeks, now, in the course of a season a whole tribe or generation develops, counting thousands of individuals, organized in one society, easily aroused, well armed, and able to furiously protect their prosperity. See next Figure.

Figure 6 : The hornet Vespa crabro L. guarding the entrance to the "commonly possessed" nest. (After MALYSHEV, 1966)
It is remarkable that this whole complex and deep-seated evolution of the way of life and behavior of the pleated-winged wasps [Eumenidae, Vespidae] did not involve deep changes in their morphology. Now it is already clear that the social wasps do not form a special systematic group sharply distinguished by structural features from solitarily living forms. Together with this, no polymorphism of individuals in a wasp society was developed, apart from some indications of it, observed in the highest societies of tropical Polybiinae, and [apart from] the fact that all female individuals in the society, with exclusion of the one fertile "tsaritsa" [queen], have lost the ability of reproduction and have changed in this way into "workers", or auxiliary females serving the society. [So only in this fashion we can speak of polymorphism in a wasp society].
In this way, the evolution of the pleated-winged wasps in the present Phase had affected basically only their instincts, and along this path, from, apparently, unassailable difficulties, present in the beginning, has led them out into a wide space [of opportunities], where vespine life received the clearest and most complex expression.
Sociality in wasps has probably originated a number of times independently : Apart from having originated from the family Eumenidae (a family of pleated-winged wasps), it has also originated from certain members of sphecoids (digger wasps, with the Sphecidae as the most important family) :
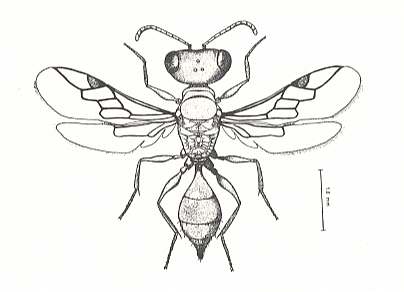
Figure 7 : Microstigmus comes, a minute sphecid wasp occurring in tropical rain forests, that may represent the closest approach to sociality by members of its family [Sphecidae]. (The bar on the right measures 1. 0 mm) (After MATTHEWS, 1968, in EVANS and EBERHARD, 1973)
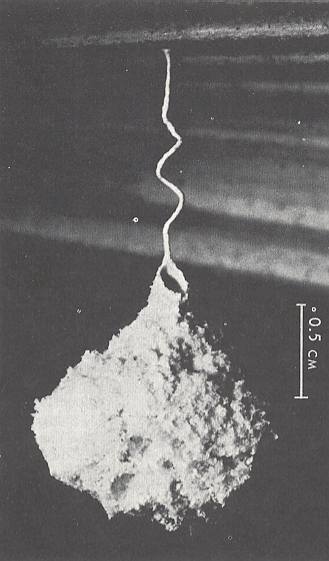
Figure 8 : A nest of Microstigmus comes suspended by a coiled pedicel from the underside of a Crysopila frond. The nest is built of fibers scraped from the frond.
(After a photograph by RETTENMEYER, from MATTHEWS, 1968, in EVANS and EBERHARD, 1973)
Discussing the subsequent path of development of the so-called social structure of vespine life is beyond the limits of the present work [of MALYSHEV], the more so because precisely with respect to this question fundamental investigations are published in the scientific literature.
We here deem it only possible to point to the final result of vespine evolution, a result, having at its base a whole series of characteristic instincts having gone a long way of development, and [these instincts] having been revealed in a certain group of solitary wasps only in their final phase of development. These instincts [already encountered in the mentioned solitary wasps] are characteristic also of social wasps, and from the basis of those instincts precisely also originated their new "social" traits of vespine life.
With this we conclude the Newly Vespine (vespoid) Phase of hymenopterous evolution.
Following MALYSHEV's account, we just have arrived at the Social Wasps (family Vespidae). Because the structure and further evolution of societies of wasps is fairly extensively discussed in the literature, MALYSHEV did not deem it necessary to dwell longer at the social wasps (his book is only about the main evolutionary paths followed by Hymenoptera). We, however, are concerned, not only with the main course of (hymenopterous) evolution, but also with all details in so far as they might challenge our general interpretation of evolution in terms of the Implicate and Explicate Orders of Reality (See [for instance] at the end of Part LI of the present Series). And indeed, sociality in wasps, its origin and structure, must eventually come to be described in terms of our noëtic theory in order to test, amend, or supplement this theory (The same goes for sociality in ants and bees, and also in termites (Order Isoptera)). So, having until now followed MALYSHEV in his series of evolutionary Phases, we, in the next document will insert an additional Phase -- the Ultimate Vespine (social) Phase of hymenopterous evolution. Although in the present document the origin of sociality is already more or less expounded (following MALYSHEV), we will present some ideas about it obtained from other sources. And to understand the main features of vespine social life it is very instructive to study it in the wasp Polistes (pleated-winged wasps, family Vespidae, subfamily Polistinae), which we will do in the next document.
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LIV.
Back to Homepage
Back to Contents
Back to Evolutionary Part XIV
Back to Evolutionary Part XV
Back to Evolutionary Part XVI
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XIX
Back to Evolutionary Part XX
Back to Evolutionary Part XXI
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XL
Back to Evolutionary Part XLI
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part L
Back to Evolutionary Part LI
Back to Evolutionary Part LII