
[Also in these enquiries into the wing-venation and its transformations in Diptera-Nematocera we will consider the subcostal vein (Sc) at most only occasionally. This vein, especially its terminal branches (Sc1, Sc2), is -- in Diptera -- often difficult to discern.]
This superfamily contains three families : Nemopalpidae, Phlebotomidae, Psychodidae.
The venational groundplan of the Psychodidea can be represented by the wing-venation of Nemopalpus zelandiae :
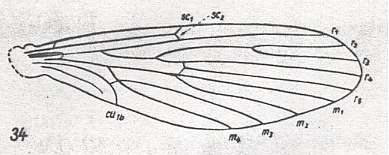
Figure 1 : Wing-venation of Nemopalpus zelandiae ALEX. Order Diptera, Family Nemopalpidae, Superfamily Psychodidea.
(After ALEXANDER, 1927, from HENNIG, 1954)
Radial Sector (Rs) 4-branched. All branches end up in the wing-margin. No capture of R4 by R2+3. M is four-branched, thus Rs and M just as in Tanyderidae. But CuA (= Cu1b) is short and bending toward (and reaching) the wing-margin, and first anal vein apparently present, but weakly developed. Second anal vein absent. Compare with Tanyderidae, Figure 2 in Part XVI . Discoidal cell absent. Wing narrow, not pointed.
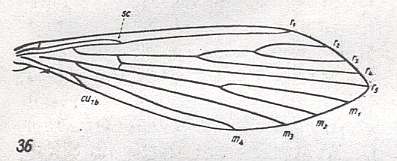
Figure 2 : Wing-venation of Phlebotomus papatasii SCOP. Order Diptera, Family Phlebotomidae, Superfamily Psychodidea.
(After HENNIG, 1954)
Radial Sector (Rs) is 4-branched. But here R2+3 does not branch off from the main stem of Rs but from R4 (= capture of R4 by the vein R2+3, in which, however, R2 does not end up in R1, but simply in the wing-margin.). M is 4-branched. CuA (= Cu1b) even more shortened than it is in Nemopalpus. The cross-vein tb (= mcu) lost as a result of the coalescence of CuA and the extreme basal piece of M4. Discoidal cell absent. Wing narrow and pointed.
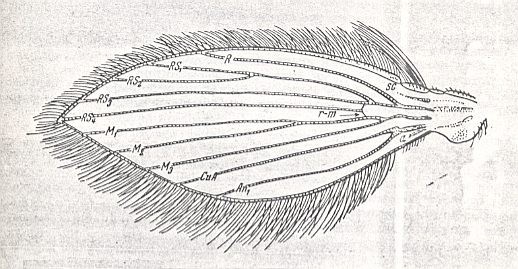
Figure 3 : Wing of Psychoda sp. Order Diptera, Family Psychodidae, Superfamily Psychodidea.
(After ROHDENDORF, 1951)
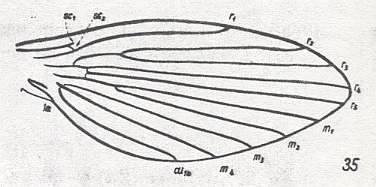
Figure 4 : Wing-venation of Ulomyia fuliginosa MEIG. Order Diptera, Family Psychodidae, Superfamily Psychodidea.
(After HENNIG, 1954)
All bifurcation points markedly shifted toward the wing-base, resulting in the forks themselves being very long, and their stalks short. Wing broad and pointed. Radial Sector (Rs) 4-branched. R2+3 branches off from main stem of Rs. Media 4-branched, depending on interpretation (see Figure 3). CuA (= Cu1b) is not shortened and not bended toward the hindmargin but almost straight. First anal vein very short, depending on interpretation, see Figure 3. Discoidal cell absent.
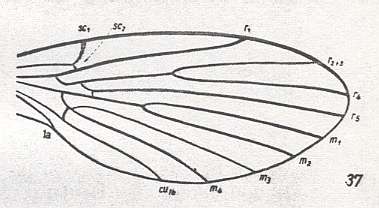
Figure 5 : Wing-venation of Trichomyia urbica CURT. Order Diptera, Family [probably] Psychodidae, Superfamily Psychodidea.
(After HENNIG, 1954)
Also here all bifurcation points markedly shifted toward the wing-base, and thus the forks themselves very long. Radial Sector (Rs) 3-branched. R2+3 branches off from R4, not forked. The venation of Phlebotomus (see Figure 2 ) shows, or suggests, that it is indeed the fork R2-R3 that has disappeared. M is 4-branched. CuA (= Cu1b) is long, almost straight. First anal vein (1a) short. Discoidal cell absent. Wing moderately broad, less pointed than in Ulomyia, Phlebotomus and Psychoda.
The main evolutionary trend in the history of the wing-venation of the Psychodidea is a shift of the main bifurcations toward the wing-base (whereas in the Tipuloids this shift was toward the wing-apex), and a broadenig of the wing.
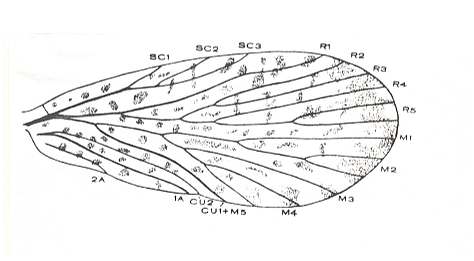
Figure 6 : Fore (?) wing of Agetopanorpa maculata CARP. Length 9.0 mm. Lower Permian of Kansas, U.S.A.
(After CARPENTER, 1930)
By a reduction in size and a slight reduction in the venation such a wing could give rise to one that resembles that of the recent Nemopalpus, see Figure 1 .
Of course, like in the case of the tipuloids, we cannot definitely indicate a particular fossil mecopteron representing a species that has given rise to the first psychodideans. Fossils are sparce as compared to the actual number of corresponding insects that actually lived at the time. Moreover, it seems that the transitions from mecopterons to dipterons were relatively fast, resulting in there having been only few transitional forms.
The superfamily Culicidea includes three families : Dixidae, Chaoboridae, and Culicidae, of which the latter comprises the well-known mosquitoes.
[According to ROHDENDORF, 1964, these families are divided among two superfamilies, the Culicidea (Culicidae, Chaoboridae) and Dixidea (Dixidae). For the present purpose it is perhaps better to unite these three families into a single superfamily. Of the Chaoboridae we have no precise venational data at our disposal, but its seems certain that their wings do not significanly differ from those of the Culicidae.]
The venational groundplan of the wings in the superfamily Culicidea can be represented by the wing-venation in Dixa :
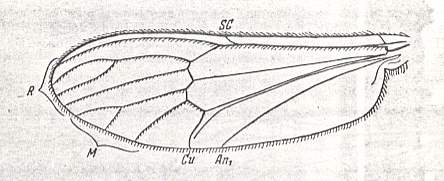
Figure 7 : Wing of Dixa sp. Length 4.5 mm. Order Diptera, Family Dixidae, Superfamily Culicidea.
(After ROHDENDORF, 1951)
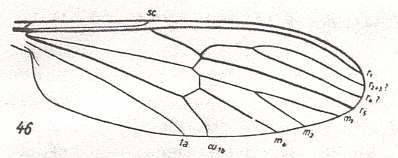
Figure 8 : Wing-venation of Dixa maculata MEIG. Order Diptera, Family Dixidae, Superfamily Culicidea. As usual in HENNIG's figures, CuP (which, for most of its length, is just a fold or groove), is not drawn. But we see it drawn in Figure 7.
(After HENNIG, 1954)
R5 and R4 present. R2+3 branches off from R4 (capture of R4 by R2+3). R2+3 not forked, reaching wing-margin. So the Radial Sector (Rs) is 3-branched. M3 absent. M1+2 forked. So M is 3-branched. Wing moderately elongate.
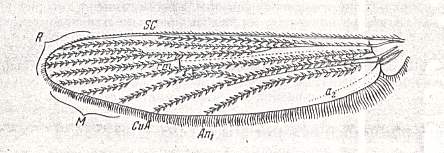
Figure 9 : Wing of Culex pipiens L. Order Diptera, Family Culicidae, Superfamily Culicidea. (After HENDEL, from ROHDENDORF, 1951)
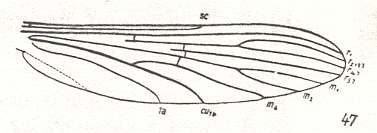
Figure 10 : Wing-venation of Anopheles maculipennis MEIG. Order Diptera, Family Culicidae, Superfamily Culicidea.
(After HENNIG, 1954)
The wing-venation in Anopheles is (formally) identical to that in Culex and Dixa. R4 and R5 present. Capture of R4 by R2+3. The latter not forked. Rs 3-branched. M3 absent. M1-M2 fork present. Wing elongate.
The venation in Culicidea differs from that of the venational groundplan of the Psychodidea (see Figure 1 ) in having both the Radial Sector and the Media 3-branched. But it would be premature to believe simply that the wing-venation in Culicidea actually evolved from that of Nemopalpus in Psychodidae. The Culicidea probably evolved from their 'own' stock of Mecoptera. Which stock cannot be determined.
In the next document we will consider the wing-venation in the superfamily Chironomidea.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XIX.
Back to Evolutionary Part XVII