
The wing-venation in Hymenoptera
We now apply the findings of the previous document to the insect Order Hymenoptera and the venation of their forewings. Although this and the previous document are devoted to the wing-venation of the Hymenoptera-Apocrita, we include here in our derivations -- from the hymenopterous prototype -- also a number of Symphyta, because their venation is important for understanding that of the Apocrita.
The prototypic venation of the forewing of Hymenoptera might be the followng :
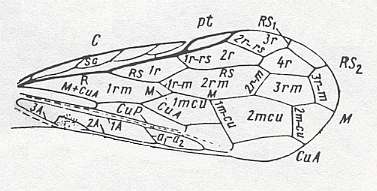
Figure 1 : Prototypic forewing-venation of Hymenoptera, as assumed by RASNITSYN, 1980.
In this forewing (from which all venational patterns in Hymenoptera are derivable) we see the following :
Cross-veins are indicated by lower-case symbols connected by a hyphen, while cells are indicated by lower-case symbols without hyphen. Longitudinal veins are indicated by capital letters, as in M, RS, CuA, 1A etc. pt - pterostigma (area).
See, if necessary, for more details of the prototypic venation of Hymenoptera (and consequently also of Symphyta), the previous document.
Soon, in the evolution of already the Symphyta the up-arching Media suppresses the cross-vein 1r-m and merges for some short distance with the Radial Sector, as the next figure shows :
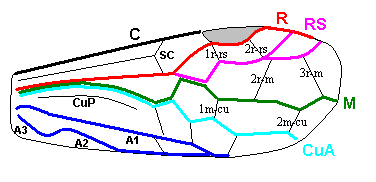
and the venation in this figure might as well be considered to represent the prototypic (fore)wing-venation of the Order Hymenoptera.
The above given Order-Prototype (of forewing) of the Hymenoptera can be seen as itself having been derived from the venational prototype of all insects. If we take the latter to be what is proposed by Comstock and Needham, then we have :
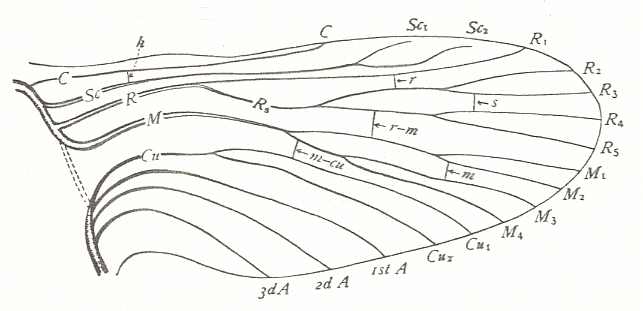
from which then the Hymenopterous Order-Prototype
is derived by (1) proliferation of the SC system, (2) loss of two branches of RS, (3) the Media and the Cubitus becoming unbranched, and (4) transformation of the anal system. Because the many-branched Subcosta of the hymenopterous prototype is not so certain, we globally can say that the latter prototype has been derived from the prototype of all winged insects by reductions of certain branches of the principal veins. The other elements of derivation are the coalescence of the Cubitus (CuA) and the Media, separating (in M and CuA) only near midwing, and the disappearance of the difference (as to look and strength) between longitudinal veins and cross-veins. The course of the main veins in the hymenopterous prototype is not smooth as it is in the prototype of all insects.
Derivations from Order-Prototype
Let us now check the forewing-venation of some fossil and recent Hymenoptera, that is to say, we'll see whether and how it can be derived from the Order-prototypic wing-venation of Figure 1 . We express this derivability by showing that the venation of the wing examined allows us to describe it in terms of the venation of this prototype.
Suborder Symphyta
|
|
Prototypic forewing-venation of Hymenoptera. |
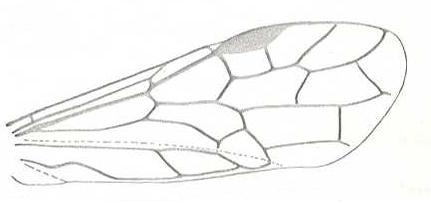 |
Forewing of Macroxyela. Family Xyelidae. Recent. Suborder Symphyta. (Figure 117A of previous previous document) |
Venational derivation (of the just depicted Macroxyela) from the Order-prototype : Subcosta has become 2-branched by reduction. Base of RS shortened, and about perpendicular to R. Two-branched RS still preserved. Pterostigmal area sclerotized. The cross-vein 1r-m suppresed by the up-arching Media. All other named cross-veins of the prototype preserved. Anal area unchanged.
As one can see, in the recent Macroxyela there is still no sign of any differentiation of parts of the forewing : As in the prototype, the veins are more or less evenly distributed over the wing-blade. So in terms of natural selection this primitive feature is not replaced by some more advanced one that might improve flight. Of course, such advanced forms are formed, but they did not replace the primitive forms. And this fact may be an indication that the advanced forms did not evolve from the primitive forms.
|
|
Prototypic forewing-venation of Hymenoptera. |
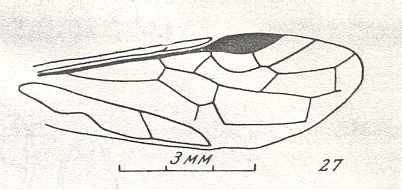 |
Xyelinus majus Rasn. Upper Triassic, Central Asia, Djailjaucho. Family Xyelidae, subfamily Archexyelinae. Suborder Symphyta. (Figure 13 of previous previous document) |
Venational derivation (of the just depicted Xyelinus) from the Order-prototype : Pterostigmal area sclerotized. SC has lost most of its branches. The up-arching Media has suppressed the cross-vein 1r-m and is now merged with RS for a short distance. The cross-vein between the two branches of RS has disappeared. These four changes, with respect to the prototype, apparently have taken place almost immediately after the Order Hymenoptera had been established, or, said differently, the fully proptotypic venation is compatible with the qualitative content of only a few species. On the other hand, the modified form of it (as a result of these four changes, or better, as a result of these four formal differences) is compatible with a great many other species of Hymenoptera especially with many Symphyta. In the other derivations we will not mention these four differences anymore.
The remaining venational characters of the prototype have been preserved (in Xyelinus).
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Ferganoxyela sogdiana Rasn. Upper Triassic, Central Asia, Djailjaucho. Family Xyelidae, subfamily Archexyelinae. Suborder Symphyta. ( Figure 16 of previous previous document) |
Venational derivation (of the just depicted Ferganoxyela) from the Order-prototype : Subcosta disappeared (either atrophied or merged with R). Wing-shape changed from elongated to triangular.
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Shurabisca liassica Rasn. Lias of Central Asia, Shurab. Family Myrmiciidae. Suborder Symphyta. ( Figure 36a of previous previous document) |
Venational derivation (of the just depicted Shurabisca) from the Order-prototype : Subcosta disappeared (either atrophied or merged with R). RS sharply arching up after its separation from RS+M. It has become unbranched. The cross-veins 2r-m and 3r-m disappeared.
Here, already in liassic times (early Jurassic), we see a certain degree of differentiation of parts of the wing-blade. And this is an indication that there are, in Hymenoptera at least, no long-time evolutionary trends finally resulting in some overall progressive state of the wing-venation.
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Karatavites medius Rasn. Upper Jurassic of southern Kazachstan (Michailovka). Family Karatavitidae. Suborder Symphyta. ( Figure 85 of previous previous document) |
Venational derivation (of the just depicted Karatavites) from the Order-prototype : Subcosta disappeared (or merged with R). RS unbranched. Basal half of the wing has become slender. The normally curved A2+A3 is straightened out.
|
|
Prototypic forewing-venation of Hymenoptera. |
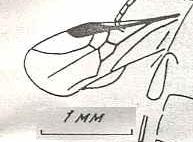 |
Microryssus brachyurus Rasn. Upper Jurassic of southern Kazachstan (Michailovka). Family Paroryssidae. Suborder Symphyta. ( Figure 94 of previous previous document) |
Venational derivation (of the just depicted Microryssus) from the Order-prototype :
Subcosta disappeared (or merged with R). RS unbranched. The cross-vein 2r-rs has disappeared. The cross-vein 3r-m has disappeared, as well as the cross-vein 2m-cu.
Explanation of this derivation : In the venation of Microryssus we see the following features :
From R, just before the pterostigma, RS branches off. It merges for a short part of its course with M, then meets the cross-vein 1r-rs and then heads into the direction of the wing-apex (the cross-vein 2r-rs is absent). The Radial Sector is not branched. M, after having left RS, heads to the apex of the wing. At the point of the separation of M from RS we see a short oblique vein which is the cross-vein 1m-cu. It connects M (in fact M+RS) with CuA.
|
|
Prototypic forewing-venation of Hymenoptera. |
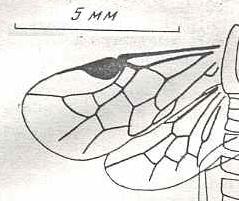 |
Schizocerus konowi Rohwer. Oligocene of Colorado [USA] (Florissant). Family Argidae. Suborder Symphyta. ( Figure 111 of previous previous document) |
Venational derivation (of the just depicted Schizocerus) from the Order-prototype : Subcosta disappeared (or merged with R). The Media arches all the way up to the branching-off point of RS from R, having thus suppressed the basal part of RS, then runs obliquely downward for some part of its course merged with RS, and then, after having separated again from RS continues its course toward the distal part of the wing without reaching the margin. Where it [M] separates from RS the latter runs abruptly up for a very short distance (this section looking like a cross-vein) until it meets the cross-vein 1r-rs (itself connecting RS with the base of the pterostigma), then heads to about the wing-apex while giving off the cross-veins 2r-m and 3r-m.
|
|
Prototypic forewing-venation of Hymenoptera. |
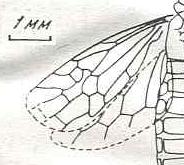 |
Pinicolites graciosus Meunier. Oligocene of Germany (Rott). Family Xyelidae. Suborder Symphyta. ( Figure 115 of previous previous document) |
Venational derivation (of the just depicted Pinicolites) from the Order-prototype : Subcosta still present, 2-branched. Pterostigmal area still unsclerotized. Basal part of RS directed toward the wing-apex (its still primitive condition). The rest of RS, after having gone up and down, proceeds toward the wing-apex and splits up into two short branches. The trunk M+CuA only splits up into M and CuA well beyond mid-wing. The Media soon gives off a very short cross-vein, 1r-m, to RS (which is, as a very primitive condition, also the case in the prototype), then continues its course toward about the wing-apex.
|
|
Prototypic forewing-venation of Hymenoptera. |
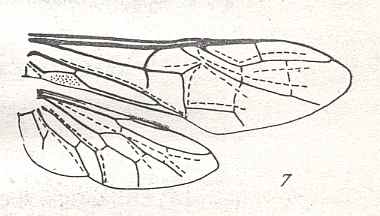 |
Cimbex Oliv., male. Family Cimbicidae. Recent. Suborder Symphyta. ( Figure 119 of previous previous document) |
Venational derivation (of the just depicted Cimbex) from the Order-prototype : Subcosta disappeared (or merged with R). The Media suppresses the origin of the Radial Sector, and the segment of the latter, between the Media and the place where 1r-rs is supposed to be, has disappeared. Cross-veins 1r-rs and 2r-rs still present, but the former (supposed to be) strongly modified. The Radial Sector has become unbranched.
|
|
Prototypic forewing-venation of Hymenoptera. |
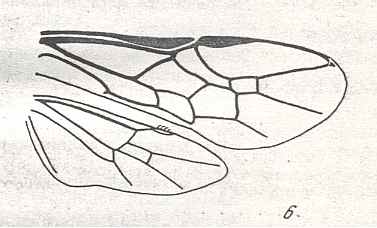 |
Wings of Acordulecera Say., Family Pergidae. Recent. Suborder Symphyta. ( Figure 118 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Acordulecera) from the Order-prototype : Forewing : Subcosta disappeared (or merged with R). The costal-subcostal-radial area highly specialized. RS unbranched, curving up to the end of R well beyond the pterostigma. The cross-vein 2r-rs wanting. The cross-vein 1r-rs looking like the basal part of RS because the part of RS between M and 1r-rs is lacking. The up-arching Media almost completely suppresses the base of the Radial Sector. First anal vein present. Anal area transformed as compared with prototype.
|
|
Prototypic forewing-venation of Hymenoptera. |
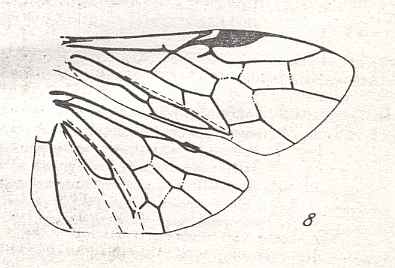 |
Wings of Neodiprion Rohw., Family Diprionidae. Recent. Suborder Symphyta. ( Figure 120 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Neodiprion) from the Order-prototype : Forewing : Subcosta vestigial. Pterostigmal area highly specialized. Maybe SC plays a part in it. RS unbranched. Section of RS between M and 1r-rs partly atrophied (see also Acordulecera in previous Figure). Cross-vein 2r-rs wanting. Up-arching Media suppressing basal part of RS. Anal area slightly transformed.
|
|
Prototypic forewing-venation of Hymenoptera. |
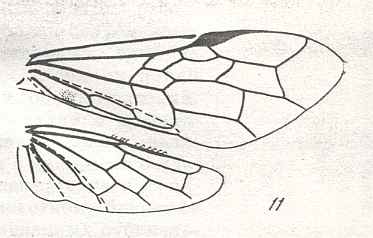 |
Wings of Megalodontes Latr., Family Megalodontidae. Recent. Suborder Symphyta. ( Figure 121 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Megalodontes) from the Order-prototype : Forewing : Subcosta wanting. RS unbranched, curving up to R. Cross-veins 1r-rs and 2r-rs both present. Basal part of RS running obliquely backwards. Radio-Medial T-vein and Radio-Medial Triangle well developed. Anal area transformed.
|
|
Prototypic forewing-venation of Hymenoptera. |
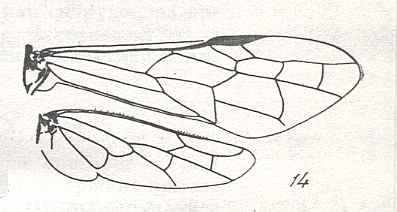 |
Wings of Syrista Knw., Family Cephidae. Recent. Suborder Symphyta. ( Figure 122 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Syrista) from the Order-prototype : Forewing : Subcosta wanting. RS unbranched. Cross-veins 1r-rs and 2r-rs both present. Basal part of RS very short, therefore Radio-Medial T-vein not developed. Nevertheless a clear triangle is formed by R, M, and M+CuA. Anal area strongly transformed. First anal vein well developed. Apparently what is 2A+3A as one straight vein running closely and parallel along the posterior wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
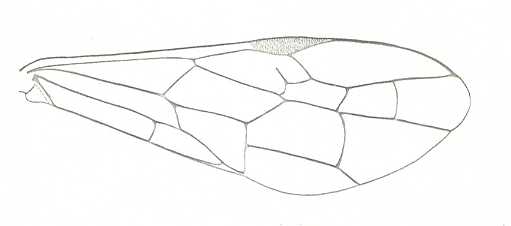 |
Forewing of Janus abbreviatus , Family Cephidae. Recent. Suborder Symphyta. ( Figure 122A of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Janus) from the Order-prototype : Subcosta wanting. RS unbranched. Cross-vein 1r-rs partly reduced, 2r-rs present. Basal part of RS very short, therefore Radio-Medial T-vein not developed. Nevertheless a clear triangle is formed by R, M, and M+CuA. Anal area strongly transformed. First anal vein well developed. Apparently what is 2A+3A as one straight vein coinciding with the posterior wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
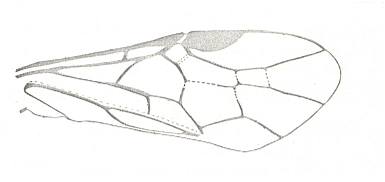 |
Forewing of Pteronidea ribesii, Family ? Cephidae. Recent. Suborder Symphyta. ( Figure 122B of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Pteronidea) from the Order-prototype : Subcosta specialized. Except for terminal branch, apparently coalesced with R. RS unbranched. Cross-vein 1r-rs present as a strong vein. The cross-vein 2r-rs wanting. Basal part of RS suppressed by up-arching Media. Part of RS between Media and 1r-rs more or less atrophied. First anal vein present. Anal area transformed.
|
|
Prototypic forewing-venation of Hymenoptera. |
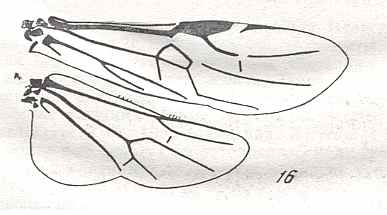 |
Wings of Orussus F., Family Orussidae. Recent. Suborder Symphyta. ( Figure 123 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Orussus) from the Order-prototype : Forewing : Venation markedly reduced. Subcosta absent or coalesced with R. RS unbranched, not reaching wing-margin. Connection between Media and Radial Sector completely lost, except for a vestige of 2r-m. The cross-vein 2r-rs present in the form of a continuation of RS. Anal area transformed.
|
|
Prototypic forewing-venation of Hymenoptera. |
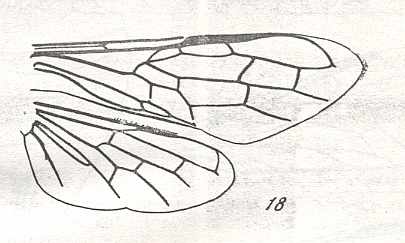 |
Wings of Urocerus Geoffr., Family Siricidae. Recent. Suborder Symphyta. ( Figure 124 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Urocerus) from the Order-prototype : Forewing : Subcosta 2-branched. Radius after elongated pterostigma curves down to meet RS. The latter unbranched, not reaching wing-margin. Cross-veins 1r-rs and 2r-rs present. The basal part of RS runs obliquely backwards, and then, after having touched the up-arching Media, the Radial Sector sharply turns around and heads toward the apical margin of the wing without reaching it. Anal area not transformed.
|
|
Prototypic forewing-venation of Hymenoptera. |
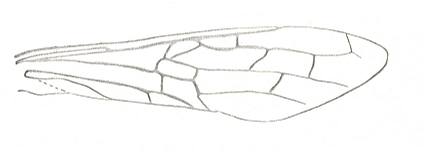 |
Forewing of Sirex, Family Siricidae. Recent. Suborder Symphyta. ( Figure 124A of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Sirex) from the Order-prototype : Subcosta has become unbranched. Radius straight, running well beyond the pterostigmal area, and then curving down to meet RS (see Urocerus (previous Figure)). Pterostigma not developed. Base of RS directed obliquely backwards, merging with M for a very short section, and then RS continues its course toward the wing apex which it doesn't reach. Radio-Medial T-vein moderately developed. Radio-Medial Triangle clearly developed. Cross-veins 1r-rs and 2r-rs present. First anal vein present. Anal area little changed. The wing has become very elongate.
|
|
Prototypic forewing-venation of Hymenoptera. |
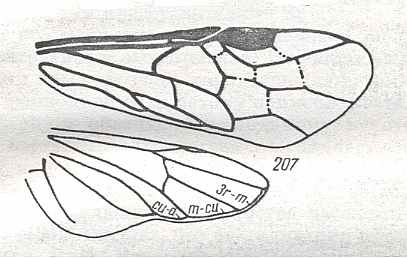 |
Wings of Caliroa Costa. male. Family Tenthredinidae. Recent. Suborder Symphyta. ( Figure 128 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Caliroa) from the Order-prototype : Forewing : Subcosta merged with R except its end. Radius running well beyond the pterostigmal area and meeting RS at the wing-margin. Pterostigma well developed. Base of RS suppressed by up-arching Media. Cross-veins 1r-rs and 2r-rs present. First anal vein present. Anal area little changed. The wing has become moderately elongate.
|
|
Prototypic forewing-venation of Hymenoptera. |
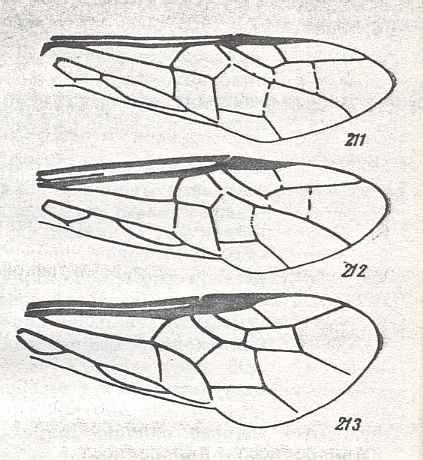 |
Forewing of Cimbex Oliv. female. Family Cimbicidae. Recent. Suborder Symphyta.
Forewing of Zaraea Leach. Family Cimbicidae. Recent. Suborder Symphyta.
Forewing of Corynis Thunb. Family Cimbicidae. Recent. Suborder Symphyta.
|
Venational derivation (of the just depicted Cimbex, Zaraea, Corynis) from the Order-prototype : General forewing-characteristics of these three genera : Subcosta absent or vestigial. Pterostigma moderately developed. Base of RS suppressed by up-arching Media. Cross-veins 1r-rs and 2r-rs present. Secrion of RS between Media and 1r-rs missing. Cross-veins 2r-m and 3r-m present. First anal vein present. Anal area changed. The wings have become moderately elongate.
|
|
Prototypic forewing-venation of Hymenoptera. |
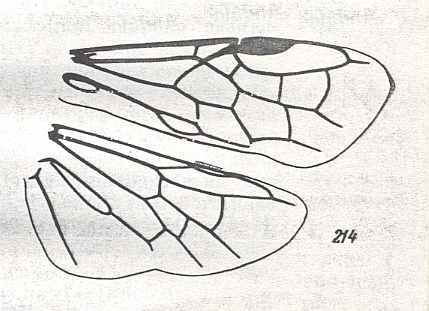 |
Wings of Arge Schrnk. Family Argidae. Recent. Suborder Symphyta. ( Figure 130 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Arge) from the Order-prototype :
Forewing : Subcosta vestigial, but its end-branch preserved. Radius running well beyond the pterostigmal area, bending down to meet RS. Pterostigma well developed. Base of RS suppressed by up-arching Media. From this point the Radius curves up to the pterostigma and beyond. After leaving M, the Radial Sector goes steeply up (looking like a cross-vein) until its meets the cross-veins 1r-rs, then proceedes into the direction of the wing tip, meeting with R and not reaching the wing-margin. The cross-vein 2r-rs has vanished. The cross-veins 2r-m and 3r-m present. First anal vein present. Anal area changed.
In Eriocampa (family Tenthredinidae) ( Figure 131 ) the forwing venation is the same, except that the anal area is virtually unchanged as compared to the Order-prototype. Another instance of independent development.
|
|
Prototypic forewing-venation of Hymenoptera. |
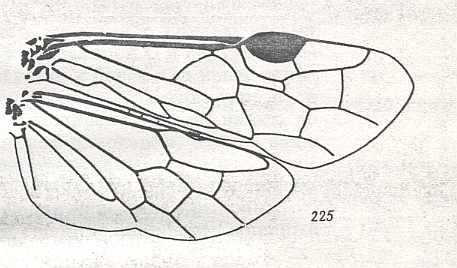 |
Wings of Blasticotoma Klug. Family Blasticotomidae. Recent. Suborder Symphyta. ( Figure 132 of previous previous document) (scrolling may be necessary) |
Venational derivation (of the just depicted Blasticotoma) from the Order-prototype : Forewing : Subcosta vestigial. Pterostigma well developed. Base of RS almost completely suppressed by up-arching Media. The section of RS between M and 1r-rs missing. The latter is fluently connected with the rest of RS that runs past the cross-vein 2r-rs towards the apex of the wing but well before it curves up to meet the anterior wing-margin. Although no Radio-Medial T-vein is developed, the up-arching part of the Media and the triangular area closed by it are conspicuous. Anal area unchanged.
Suborder Apocrita
|
|
Prototypic forewing-venation of Hymenoptera. |
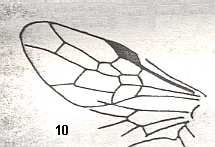 |
Wing(s) of Asiephialtites oviventer, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 3.5 mm. ( Figure 9 of previous document) |
Venational derivation (of the just depicted Asiephialtites) from the Order-prototype : Forewing : Subcosta absent. Pterostigma well developed. Radio-Medial Y-vein developed. Origin of RS shifted toward the pterostigma, and thus taking the place of the cross-vein 1r-rs [this could mean that in fact the base of RS has been lost]. Cross-vein 2r-rs present. RS unbranched. Cross-veins 2r-m and 3r-m both present. RS, M, and CuA reaching the wing-margin. The basal half of the wing is markedly narrowed and the anal area and veins strongly reduced, if not totally vanished. Here it is especially clear that, unlike as in most wings of Diptera, the forewing failed to produce a widened anal area, this area being, on the contrary, reduced. In these Hymenoptera the latter is in fact functionally present in the form of the small hindwing attached by hooks on the forewing. The reduction of the anal area of the forewing is especially evident in Asiephialtites ventricosus ( Figure 10 of previous document ).
|
|
Prototypic forewing-venation of Hymenoptera. |
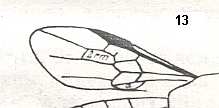 |
Wing of Parephialtites reductus, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 4.4 mm. ( Figure 11 of previous document) |
Venational derivation (of the just depicted Parephialtites) from the Order-prototype :
Subcosta absent. Pterostigma well developed. Radio-Medial Y-vein developed. Origin of RS well before pterostigma. Vestige of the cross-vein 1r-rs present. Cross-vein 2r-rs present. Cross-vein 2r-m present. Cross-vein 3r-m absent. RS unbranched, reaching wing-margin. Media and CuA not reaching wing-margin. First anal vein present.
Here, in Apocrita, we see, already in upper jurassic times, a differentiation of parts of the wing-blade, so characteristic of recent Apocrita (see, for exampe, Figure 90, previous document , Figure 93B, prev doc , and Megachile below ).
|
|
Prototypic forewing-venation of Hymenoptera. |
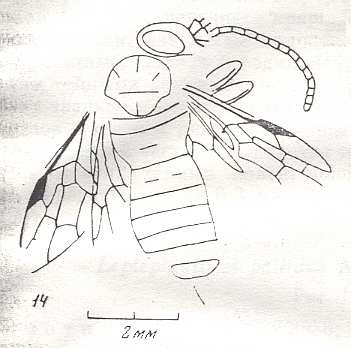 |
Mesephialtites paurocerus, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 4 mm. ( Figure 12 of previous document) |
Venational derivation (of the just depicted Mesephialtites) from the Order-prototype : Subcosta absent. Pterostigma well developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. Origin of RS shifted towards the base of the pterostigma. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-veins 2r-m and 3r-m both present. Media and CuA reaching wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
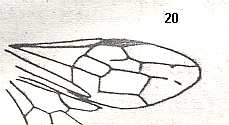 |
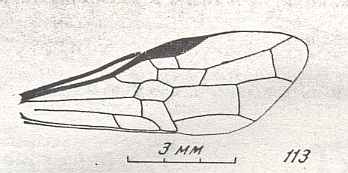
Wing(s) of Leptephialtites tenuicornis, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 8.1 mm. ( Figure 15 of previous document) |
Venational derivation (of the just depicted Leptephialtites) from the Order-prototype : Forewing : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. Origin of RS shifted towards the base of the pterostigma. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Wing(s) of Leptephialtites angustus, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 3.6 mm. ( Figure 17 of previous document) |
Venational derivation (of the just depicted Leptephialtites) from the Order-prototype : Forewing : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. The Triangle is, however, rather short. Origin of RS well before the base of the pterostigma. The cross-vein 1r-rs only present as a vestige. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-veins 2r-m and 3r-m both present.
|
|
Prototypic forewing-venation of Hymenoptera. |
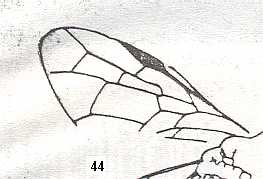 |
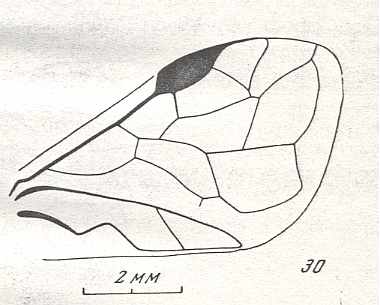
Forewing of Micrephialtites minor, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 3.5 mm. ( Figure 30 of previous document) |
Venational derivation (of the just depicted Micrephialtites) from the Order-prototype : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. The Triangle is, however, very short. Origin of RS well before the base of the pterostigma. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-veins 2r-m and 3r-m both present. RS, M, and CuA reach the wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
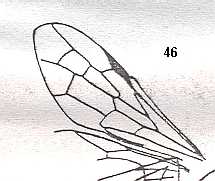 |
Wing(s) of Karataviola micrura, Family Ephialtitidae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 7.5 mm. ( Figure 32 of previous document) |
Venational derivation (of the just depicted Karataviola) from the Order-prototype :
Forewing : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. The Triangle is long as a result of the very oblique position of the base of RS. Origin of RS well before the base of the pterostigma. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-veins 2r-m and 3r-m both present. RS, M, and CuA reach the wing-margin. Anal area reduced.
A forewing with similar venation we see in the much smaller (wing-length 2.5 mm) Cleistogaster curtis (Upper Jurassic) of the family Megalyridae ( Figure 38 of previous document ).
|
|
Prototypic forewing-venation of Hymenoptera. |
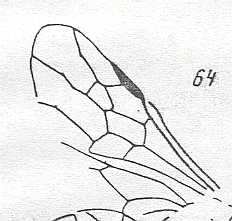 |
Wing(s) of Mesaulacinus stenocerus, Family Megalyridae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 4 mm. ( Figure 45 of previous document) |
Venational derivation (of the just depicted Mesaulacinus) from the Order-prototype : Forewing : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle moderately developed. The Triangle is long. Origin of RS well before the base of the pterostigma. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-vein 2r-m present. Cross-vein 3r-m absent. Cross-vein 1m-cu present. Cross-vein 2m-cu absent. RS, M, and CuA reach the wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
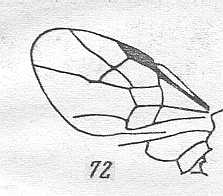 |
Wing(s) of Brachycleistogaster nana, Family Megalyridae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 1.8 mm. ( Figure 53 of previous document) |
Venational derivation (of the just depicted Brachycleistogaster) from the Order-prototype :
Forewing : Subcosta absent. Pterostigma well developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. The Triangle is broad. Origin of RS close to the base of the pterostigma. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-vein 2r-m present. Cross-vein 3r-m absent. Cross-vein 1m-cu present. Cross-vein 2m-cu absent. RS, M, and CuA reach the wing-margin (CuA almost reaches it). Anal area reduced, but the first anal vein is present. Forewing more or less triangular. Despite its small size (1.8 mm) this wing has a fairly complete venation.
Brachycleistogaster pallipes (Upper Jurassic) having a similar venation, and having the same size, has the Media and CuA by far not reaching the wing-margin ( Figure 55 of previous document).
|
|
Prototypic forewing-venation of Hymenoptera. |
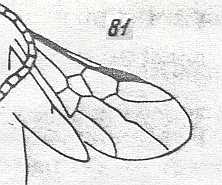 |
Wing(s) of Microcleistogaster parvula, Family Megalyridae. Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 1.5 mm. ( Figure 57 of previous document) |
Venational derivation (of the just depicted Microcleistogaster) from the Order-prototype : Forewing : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle not really developed. Origin of RS close to the base of the pterostigma, perpendicular to R. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-margin. Cross-veins 2r-m and 3r-m absent. Cross-vein 1m-cu (short, oblique) present, taking of from RS+M, and 2m-cu absent. Cross-vein 1cu-a present. RS, M, and CuA reach the wing-margin. The first anal vein is present. Also here, despite its small size (1.5 mm) this wing has a fairly complete venation.
|
|
Prototypic forewing-venation of Hymenoptera. |
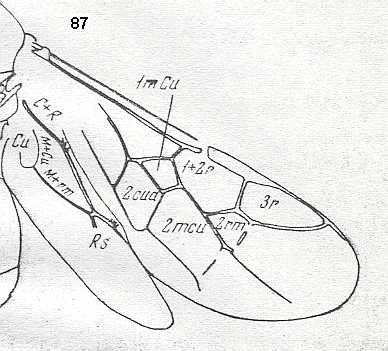 |
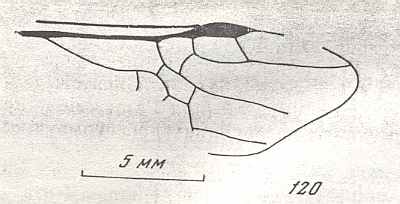
Wings of Maimetsha arctica, Family Maimetshidae (Superfamily Ceraphronoidea). Upper Cretaceous of Taimyr. Suborder Apocrita. Length of wing 1.7 mm. ( Figure 58 of previous document) |
Venational derivation (of the just depicted Maimetsha) from the Order-prototype : Forewing : Subcosta absent. Pterostigma not clearly developed. Radio-Medial T-vein and Radio-Medial Triangle not really developed. Origin of RS close to the base of pterostigmal area, perpendicular to R. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching R at the wing-margin. Cross-vein 2r-m absent or vestigial. Cross-vein 3r-m absent. Cross-vein 1m-cu (short, oblique) present, taking off from RS+M (as in Microcleistogaster [previous Figure] ). Cross-vein 2m-cu apparently vestigial. M and CuA do not reach the wing-margin. The first anal vein is present. Also here, despite its small size (1.7 mm) this wing has a fairly complete venation.
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Elasmomorpha melpomene, Family Stigmaphronidae (Superfamily Ceraphronoidea). Upper Cretaceous of Taimyr. Suborder Apocrita. Length of body 1.1 mm. ( Figure 60 of previous document) |
Venational derivation (of the just depicted Elasmomorpha) from the Order-prototype : Forewing : Costo-Radial complex present as one sclerotized region (including pterostigma). Apart from the unbranched Radial Sector, and the mentioned complex, no other veins present. (Fore)wings fairly elongated.
|
|
Prototypic forewing-venation of Hymenoptera. |
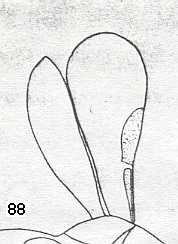 |
Wings of Stigmaphron orphne, Family Stigmaphronidae (Superfamily Ceraphronoidea). Upper Cretaceous of Taimyr. Suborder Apocrita. Length of wing 0.5--0.6 mm. ( Figure 59 of previous document) |
Venational derivation (of the just depicted Stigmaphron) from the Order-prototype : Forewing : Costo-Radial complex present as one sclerotized region (including pterostigma). Apart from this, no other veins present. (Fore)wings fairly elongated. This extremely minute forewing still possesses venational elements.
|
|
Prototypic forewing-venation of Hymenoptera. |
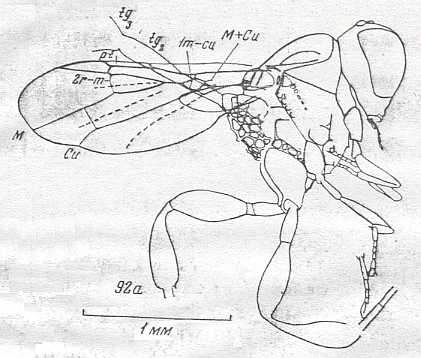 |
Cretevania minor, Family Cretevaniidae (Superfamily Evanioidea). Upper Cretaceous of Taimyr. Suborder Apocrita. Length of wing 1.7 mm. ( Figure 61 of previous document) [The long narrow structure (drawn as two lines) extending from the thorax obliquely upwards (partly lying under the wing) is the [beginning of the]abdomen.] |
Venational derivation (of the just depicted Cretevania) from the Order-prototype :
Forewing : Shape unusual for hymenoptera (and thus also for order-prototype). It is more characteristic of Diptera (So this shape is d e r i v e d with respect to that of the order-prototype) : Posterior margin broadly curved almost up to the base, [this margin being] not even (it is a little concave at the region of the end of Cu).
More or less recognizable hymenopterous venational elements in this form : M+Cu splits into Cu and M. The latter arches up and merges with RS for some distance before it separates again from RS.
Wing-blade in its distal half with sharp convex longitudinal folds between RS and M, M and Cu, and after Cu (the position of the veins themselves is concave). Distally, the wing-membrane is clearly crimped. Costal field wide, pterostigma narrow and very long. Origin of RS is well before the pterostigma, that is, it lies at a distance from the pterostigma equal to the latter's length. First section of RS vertical [i.e. perpendicular to R ], very short. M+Cu reduced except for its end. Cross-vein 1m-cu takes off from M+RS.
So also this extremely minute forewing still possesses venational elements.
With respect to the order-prototype, the venation of the forewing of Cretevania is reduced in the following way : Subcosta absent, RS unbranched, cross-vein 1r-rs absent (2r-rs present). Cross-vein 3r-m absent (2r-m present). All anal veins absent.
( Hindwing extremely short (almost 3 times shorter than the forewing), lacking all veins, except C, and lacking a jugal lobe.)
|
|
Prototypic forewing-venation of Hymenoptera. |
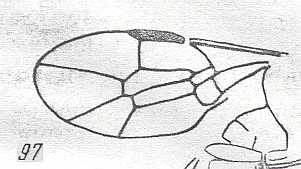 |
Forewing of Anomopterella stenocera, Family Anomopterellidae, ? superfamily Evanioidea . Upper Jurassic of Karatau. Suborder Apocrita. Length of wing 2 mm. ( Figure 63 of previous document) |
Venational derivation (of the just depicted Anomopterella) from the Order-prototype : Subcosta absent. Pterostigma fairly well developed. Radio-Medial T-vein well developed, the base of RS forming the chief part of it, that is, the section of Media taking part in it is very short. Origin of RS close to the base of the pterostigma, directed obliquely backwards. The cross-vein 1r-rs absent. Cross-vein 2r-rs present. RS unbranched, reaching wing-apex. So the position of RS is almost in the middle (median line) of the wing. Cross-vein 2r-m present, and 3r-m absent. Cross-vein 1m-cu present, taking off from M. The cross-vein 2m-cu present. Cross-vein 1cu-a present. RS, M, and CuA reach the wing-margin. The first anal vein is present. Anal area apparently narrow. Wing short and broad, but, apparently with a narrow base. Also here, despite its small size (2 mm) this wing has a fairly complete venation.
|
|
Prototypic forewing-venation of Hymenoptera. |
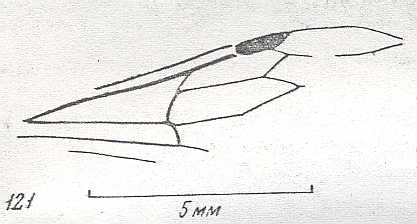 |
Forewing (fragm.) of Pompilopterus ciliatus, ? Family Pomplilidae, superfamily Pompiloidea . Lower-lower Cretaceous of Zabaikalj. Suborder Apocrita. Length of wing 8-8.5 mm. ( Figure 74 of previous document) |
Venational derivation (of the just depicted Pompilopterus) from the Order-prototype : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. Origin of RS well before the base of the pterostigma, directed obliquely backwards. The cross-vein 1r-rs absent. Cross-vein 2r-rs present as an oblique vein. At about the level of the pterostigma RS separates from RS+M, meets 2r-rs and then proceeds arch-wise into the direction of the end of the anterior wing-margin.
|
|
Prototypic forewing-venation of Hymenoptera. |
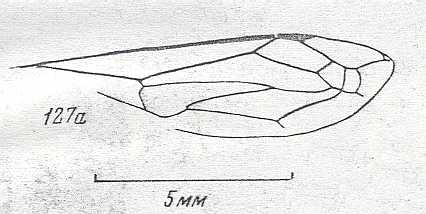 |
Forewing of Curiosivespa curiosa, Family Eumenidae, superfamily Vespoidea . Lower-upper Cretaceous of southern Kazachstan. Suborder Apocrita. Length of wing about 10 mm. ( Figure 78 of previous document) |
Venational derivation (of the just depicted Curiosivespa) from the Order-prototype : This wing has a venation that is very hard if at all to derive from the order-prototype.
Subcosta absent. Pterostigma extremely small and narow, in fact hardly developed. Radio-Medial Y-vein and Radio-Medial Triangle developed. It looks as if the Radio-Medial Y-vein has pulled the Radius downward, resulting in a large triangular area in the anterior part of the wing, an area without veins. Origin of RS well before midwing and very short, directed obliquely forward. The course of the (branched?) Radius is difficult to establish, not because of poor preservation in the fossil, but because of problems of interpretation. RS, directly after its origin merges with M for a long distance, after which M proceeds backwards while the free RS proceeds to the apical wing-margin (which it does not reach).
It might be that the curious venation of this wing is not derivable from the order-prototype at all and represents instead a separate line of hymenopterous evolution. That this wing is not some individual anomaly is shown by the fact that yet another species is found at the same site. See Figure 79 of previous document .
|
|
Prototypic forewing-venation of Hymenoptera. |
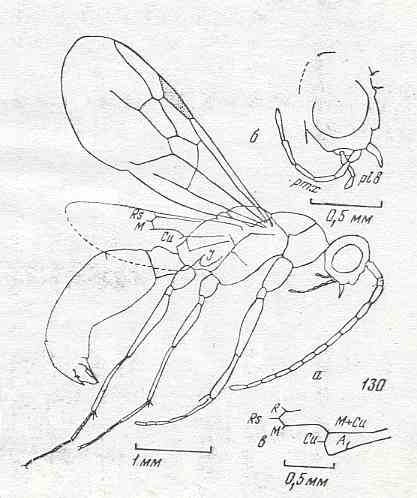 |
Palaeomyrmex zherichini, Family Formicidae, superfamily Formicoidea (ants) . Lower-upper Cretaceous of Taimyr. Suborder Apocrita. Length of wing 3.6 mm. ( Figure 80 of previous document) |
Venational derivation (of the just depicted Palaeomyrmex) from the Order-prototype : Forewing : Subcosta absent. Pterostigma moderately developed. Basal section of RS very short, perpendicular to R. Cross-vein 1r-rs absent, 2r-rs present. RS unbranched. It reaches the wing-margin. The cross-vein 2r-m present, and 3r-m absent. M, CuA, and the first anal vein do not reach the wing-margin, resulting in the postero-apical part of the wing to be without veins. The cross-vein 1m-cu is present, taking off from M. The cross-vein 1cu-a and the up-arching Media look like a single vein.
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Forewing of Trichobaissodes antennatus, Family Baissodidae. Lower-lower Cretaceous of Zabaikalj. Suborder Apocrita. Length of wing 7 mm. ( Figure 82 of previous document) |
Venational derivation (of the just depicted Trichobaissodes) from the Order-prototype : Subcosta absent. Pterostigma moderately developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. Cross-vein 1r-rs absent, 2r-rs present, markedly oblique. RS unbranched. It reaches the wing-margin. The cross-veins 2r-m and 3r-m both present as curved veins. The course of RS in this fossil especially emphasizes the common condition in hymenopterous wings : Parts of the original longitudinal veins follow transverse courses.
|
|
Prototypic forewing-venation of Hymenoptera. |
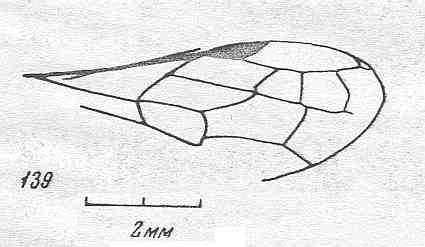 |
Forewing of Vitimosphex incompletus, ? Family Baissodidae. Lower-lower Cretaceous of Zabaikalj. Suborder Apocrita. Length of wing 6.2 mm. ( Figure 83 of previous document) |
Venational derivation (of the just depicted Vitimosphex) from the Order-prototype : Subcosta absent. Pterostigma well developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. Cross-vein 1r-rs absent, 2r-rs present, markedly oblique (looks like continuation of RS). RS unbranched, it meets with the Radius and apical wing-margin at its end. The cross-veins 2r-m and 3r-m both present. Also here the course of RS in this fossil emphasizes the common condition in hymenopterous wings : Parts of the original longitudinal veins follow transverse courses. Cross-veins 1m-cu and 2m-cu both present. Cross-vein 1cu-a present. First anal vein present.
|
|
Prototypic forewing-venation of Hymenoptera. |
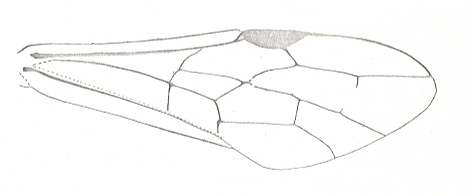 |
Forewing of Odontaulacus editus, Superfamily Evanioidea. Recent. Suborder Apocrita. ( Figure 83A of previous document) |
Venational derivation (of the just depicted Odontaulacus) from the Order-prototype :
Subcosta absent or fused with R except for its very end. Pterostigma well developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. Cross-vein 1r-rs absent, 2r-rs present. RS unbranched, it reaches the apical wing-margin. The cross-veins 2r-m and 3r-m both absent, one of them left as a vestige. Cross-veins 1m-cu and 2m-cu both present. Cross-vein 1cu-a present. First anal vein present. Anal area narrow. 2A and 3A absent. Media, CuA, and 1A all reach the wing-margin.
As we can see, this type of venation was very common already in Jurassic times. So, for instance, in Leptephialtites tenuicornis (Ephialtitidae) ( Figure 15 of previous document ). In the recent Aulacinus fusiger (Fig 83B prev doc) the cross-veins 2r-m and 3r-m are still present as in many Jurassic forms (see for the latter, for instance Karataviola micrura , Fig 32 prev doc).
|
|
Prototypic forewing-venation of Hymenoptera. |
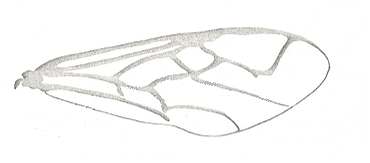 |
Forewing of Evania appendigaster, Family Evaniidae, Superfamily Evanioidea. Recent. Suborder Apocrita. Body-length about 9 mm. ( Figure 83C of previous document) |
Venational derivation (of the just depicted Evania) from the Order-prototype :
Subcosta absent or fused with R. Pterostigma well developed. Origin and base of Radial Sector shifted to the pterostigma (i.e. RS originates from the pterostigma). Radio-Medial T-vein well developed, but the 'arms' of the " T ", i.e. its top section, very oblique, almost parallel with the anterior wing-margin, resulting in the Radio-Medial Triangle to be very narrow. The base of RS is at the location of the vanished cross-vein 1r-rs. Cross-vein 2r-rs present as a thick vein. RS unbranched and sharply curved towards the anterior wing-margin, meeting R. The cross-veins 2r-m and 3r-m both absent, one of them left as a small vestige. Cross-vein 1m-cu present, and 2m-cu absent. Cross-vein 1cu-a present. First anal vein present. Anal area narrow. 2A and 3A absent. Media and CuA reach the wing-margin. 1A almost reaches it.
Very remarkable here is the shift of the origin and base of RS to the pterostigma, and the sharp turn of RS toward the anterior wing-margin. The same we see in the recent Acanthinevania princips ( Fig 83D prev doc). In the latter form the free Media and CuA are more or less reduced.
|
|
Prototypic forewing-venation of Hymenoptera. |
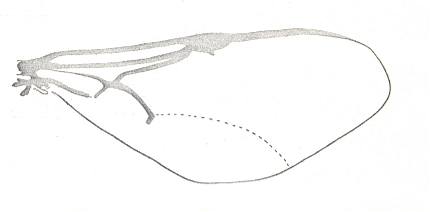 |
Forewing of Semaeodogaster barticensis, Family Evaniidae, Superfamily Evanioidea. Recent. Suborder Apocrita. ( Figure 83E of previous document) |
Venational derivation (of the just depicted Semaeodogaster) from the Order-prototype :
Subcosta absent or fused with R. Pterostigma moderately developed. Its position is at about the middle of the anterior wing-margin. Origin and base of Radial Sector shifted to the pterostigma (i.e. RS originates from the pterostigma), but RS+M absent, like the rest of RS. So there is no Radio-Medial T-vein, and also no Radio-Medial Y-vein. The base of RS is at the location of the vanished cross-vein 1r-rs. Cross-vein 2r-rs present only as a very short vestige. Free Media totally absent. Of the free CuA only its proximal third is developed, the rest of it is no more than a trace. Only the proximal part of 1A is developed. Cross-vein 1cu-a present. Anal area very narrow, 2A and 3A absent. The wing has a more or less triangular shape.
It is clear that we have here a case of intense venational reduction. More than the distal half of the wing is almost devoid of veins.
|
|
Prototypic forewing-venation of Hymenoptera. |
 |
Forewing of Hyptia, Family Evaniidae, Superfamily Evanioidea. Recent. Suborder Apocrita. ( Figure 83F of previous document) |
Venational derivation (of the just depicted Hyptia) from the Order-prototype :
Subcosta absent or fused with R. Pterostigma little developed. Its position is just after the middle of the anterior wing-margin. Base of Radial Sector absent. RS+M and M absent. The rest of RS is present in a vanishing state, by far not reaching wing-margin. While there is no trace of the (free) Media, CuA+M is present, as well as the proximal part of the free CuA. 1A almost completely reduced. 2A and 3A absent. Anal area very narrow. The wing has a more or less triangular shape.
It is clear that also here we have a case of intense venational reduction. Evidently this form cannot be derived from the previous one (Semaeodogaster), neither the other way around.
|
|
Prototypic forewing-venation of Hymenoptera. |
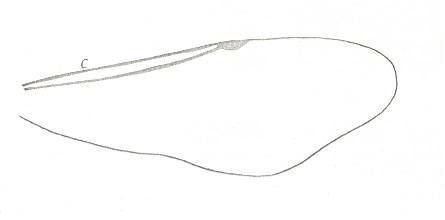 |
Forewing of Evaniellus, Family Evaniidae, Superfamily Evanioidea. Recent. Suborder Apocrita. ( Figure 83G of previous document) |
Venational derivation (of the just depicted Evaniellus) from the Order-prototype : Venation extremely reduced. Only the Costa (C), Radius, and a small pterostigma are present. The wing is more or less elongate.
|
|
Prototypic forewing-venation of Hymenoptera. |
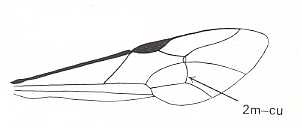 |
Forewing of an ichneumon-fly. Family Ichneumonidae. Superfamily Ichneumonoidea. Recent. Suborder Apocrita. ( Figure 84 of previous document) |
Venational derivation (of the just depicted ichneumon-fly) from the Order-prototype :
Venation moderately reduced. Radius merged with Costa. Pterostigma well developed, its position far before wing-apex. Media, separating off from CuA+M (as is usual in Hymenoptera), suppresses the origin and base of RS at the point between the end of Costa and the beginning of the pterostigma. As a consequence of this suppression, no Radio-Medial Y- or T-vein developed. Nevertheless, a Radio-Medial Triangle is well developed. RS+M absent. Of RS only the section between the cross-vein 2r-rs and its end at the anterior wing-margin is preserved. The cross-veins 2r-m and 3r-m have approached each other until they almost coincide. The cross-vein 2m-cu is present, while 1m-cu has become part of the free Media which reaches the wing-margin. Free CuA and 1A present and reaching the wing-margin. Anal area very narrow. 2A and 3A absent.
The above venational interpretation is such that the veins that are present in the wing, as well as those that have vanished, are assumed to be correctly identified as the corresponding veins in the hymenopterous venational prototype. It is, however, not excluded that this interpretation is wrong. Perhaps here too great an evolutionary transformation of the prototypic venation into that of the present wing is assumed to have taken place. This assumed evolutionary transformation includes the loss of RS+M and the extreme mutual approaching of the cross-veins 2r-m and 3r-m.
Nevertheless, there are enough elements in the venation that guarantee it to be a truly hymenopterous venation. So it must be derivable from the hymenopterous prototypic venation in the way we have suggested or in some other way. And indeed, the venation of another ichneumon-fly (see its derivation in the previous document ) reveals that RS+M has indeed vanished (because a remnant of it is still present), and that the alleged cross-vein 2r-m is in fact a section of RS, that is to say that the assumed approaching of two cross-veins has not taken place.
|
|
Prototypic forewing-venation of Hymenoptera. |
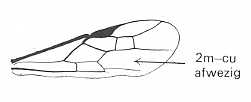 |
Forewing of a braconid fly. Family Braconidae. Superfamily Ichneumonoidea. Recent. Suborder Apocrita. ( Figure 85 of previous document) |
Venational derivation (of the just depicted braconid fly) from the Order-prototype : Radius merged with Costa. Pterostigma well developed. Media, separating off from CuA+M (as is usual in Hymenoptera), does not suppresses the origin and base of RS. The base of the latter branches off from R at the point between the end of Costa and the beginning of the pterostigma, and is directed obliquely backwards. As a consequence the Radio-Medial T-vein is well developed, so also the Radio-Medial Triangle. RS+M present. From it the free RS goes steeply upward (looking like a cross-vein) until the cross-vein 2r-rs and then heads to the wing-margin while giving off the cross-vein 2r-m. The cross-vein 1m-cu present, while 2m-cu is absent. CuA does not reach the wing-margin. Cross-vein 1cu-a present. The vein 1A present, not reaching the wing-margin. Anal area very narrow. 2A and 3A absent.
The derivation of the venation of another ichneumon-fly and another braconid (with the help of a Jurassic form) was given in the previous document. To see it, click HERE .
|
|
Prototypic forewing-venation of Hymenoptera. |
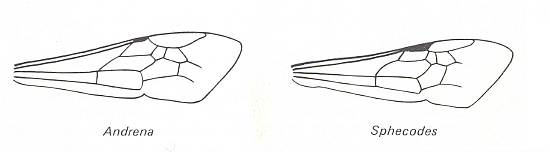 |
Left image : Forewing of Andrena. Family Andrenidae. Superfamily Apoidea (bees). Recent. Suborder Apocrita. Right image : Forewing of Sphecodes. Family Halictidae. Superfamily Apoidea. Recent. Suborder Apocrita.( Figure 88 of previous document) |
Venational derivation (of the just depicted bees) from the Order-prototype :
Radius not merged with Costa. Subcosta absent. Pterostigma hardly developed in Andrena, while fairly developed in Sphecodes. Radio-Medial T-vein and Radio-Medial Triangle developed (especially in Andrena). After RS+M, the Radial Sector goes up crossvein-like until it meets the cross-vein 2r-rs and then proceeds to the anterior wing-margin. The cross-veins 2r-m and 3r-m both present. Media, CuA, and 1A do not reach the wing-margin (that is, as a result of either the reduction of their end-sections or the expansion of the wing-membrane). The cross-veins 1m-cu and 2m-cu present. Cross-vein 1cu-a present. Anal area very narrow. 2A and 3A absent. (Fore)wing elongated, apex pointed.
It is remarkable that these wings, wings that is, that are possessed by individuals that belong to the highest Hymenoptera (Aculeata-Apoidea), still have a more or less complete wing-venation as compared to the hymenopterous prototype.
|
|
Prototypic forewing-venation of Hymenoptera. |
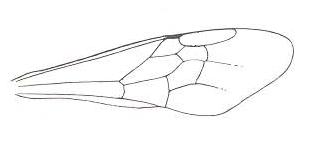 |
Forewing of Megachile. Family Megachilidae. Superfamily Apoidea (bees). Recent. Suborder Apocrita. ( Figure 89 of previous document) |
Venational derivation (of the just depicted Megachile) from the Order-prototype :
Radius not merged with Costa. Subcosta absent. Pterostigma hardly developed. Radio-Medial T-vein and Radio-Medial Triangle very well developed. After RS+M, the Radial Sector goes up crossvein-like until it meets the cross-vein 2r-rs and then proceeds to the anterior wing-margin, while strongly curving up to it, ending up far before the wing-apex. The cross-vein 2r-m is present, while 3r-m is absent. Media and CuA do not reach the wing-margin (that is, as a result of either the reduction of their end-sections or the expansion of the wing-membrane). The cross-veins 1m-cu and 2m-cu are present. Cross-vein 1cu-a present. Anal area very narrow. 2A and 3A absent. (Fore)wing elongated, apex not pointed.
As a result of the early ending of the veins RS, M, and CuA, and of the presence of the large Radio-Medial Triangle, the venation of the wing is concentrated in an oblique band in the middle of the wing. This is quite an internal differentiation of parts of the wing as compared to the order-prototype.
|
|
Prototypic forewing-venation of Hymenoptera. |
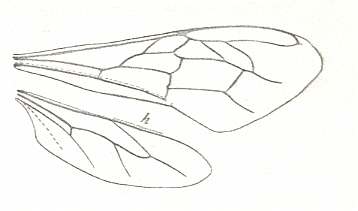 |
Wings of Apis. Family Apidae. Superfamily Apoidea (bees). Recent. Suborder Apocrita. ( Figure 93C of previous document) |
Venational derivation (of the just depicted Apis) from the Order-prototype : Forewing : Radius not merged with Costa. Subcosta absent. Pterostigma virtually not developed. Radio-Medial T-vein and Radio-Medial Triangle well developed. After RS+M, the Radial Sector goes up until it meets the cross-vein 2r-rs and then proceeds to the distal part of the anterior wing-margin where it unites with the down-bending Radius. The cross-veins 2r-m and 3r-m both present as more or less curved long veins. Media and CuA do not reach the wing-margin. The cross-veins 1m-cu and 2m-cu are present. Cross-vein 1cu-a present, shifted basad. Vein 1A present, almost reaching wing-margin. Anal area very narrow. 2A and 3A absent. (Fore)wing elongated, apex moderately pointed.
This concludes our survey of hymenopterous (fore)wings, and the investigation of in what way their venation can be formally derived from the hymenopterous venational prototype.
Noëtic evolutionary theory and the wing-venation in the Order Hymenoptera
We have now, in the last three Parts, considered extensively the wing-venation of fossil and recent Hymenoptera. Although we have not considered and interpreted the venation of all known recent Hymenoptera, we have done so with quite a lot of them, that is we have considered examples of all the major groups of Hymenoptera. And in this survey we have seen that all wing-venations considered in it can indeed be formally derived from the assumed hymenopterous venational prototype because they turned out to allow us to describe them in terms of this prototype. We have not met with any unsurmountable difficulty in so deriving them. Nowhere have we been forced to assume the appearance of truly new veins, new with respect to the hymenopterous venational prototype.
There are two fossil wings of alleged Apocrita -- see Curiosivespa above -- of which the wing-venation might not be derivable from the hymenopterous prototype. But, because in both cases only a forewing is known they might perhaps not belong to Hymenoptera at all.
Of course the assumed hymenopterous prototype was based (by Rasnitsyn, 1969, 1980) on existing fossil and recent material, and all we have done is merely to check whether this is indeed the right prototype. Our survey proves it to be so. And because it especially complies with the fossil evidence (i.e. the assumed prototype is not only based on primitive recent forms, but on fossils as well), the prototype is, as far as our knowledge stands today (that is about 1980 -- but that is, for the time being, recent enough) a true prototype. And although it is to be expected that all hymenopterous wings, fossil and recent, taken together, cannot form a single (branched) sequence in which all hymenopterous venations can be derived sequentially from each other -- and then indirectly and ultimately from the hymenopterous prototype as the first member of such a sequence, and thus form a continuous phylogenetic (branched) sequence, -- they can at least formally all be derived directly from the prototype. And this precisely was our theoretical and formal demand : The wing-venation of any given hymenopterous species must be such that it not only complies with the comstock-needham scheme of principal wing-veins, but also that it is formally derivable from the hymenopterous venational prototype. This demand is meant to be satisfied in the Implicate Order when strategies are formed there in a timeless fashion. When this demand is actually satisfied in the Implicate Order, and indeed it is, the result, upon projection into the Explicate Order, is compatible with the general empirical findings (for example the fact of the existence of a certain derivational order in Hymenoptera), and it is also compatible with the expected fact (also based on the general experience in taxonomy) that, because of existing specialization-crossings, many species or groups cannot be derived from others.
Basic to all this is the fact that in the Explicate Order, that is, by whatever factors active in that Order, transformation of (living) forms involving not just mere slight changes, but involving complex and sophisticated adaptations, cannot be accomplished, that is, cannot be accomplished without introducing (in current theory) teleological elements (i.e. that the goal acts as a cause). And teleological elements are indeed alien to the Explicate Order. Explanation of the origin and development of such adaptations by random genetic mutations and natural selection looks like being not teleologic. But different and simultaneous selection pressures (pressures for the improvement of several different organs, structures, or behavior) will undoubtedly interfere with each other, rendering a continued undisturbed sequence of incremental additions, subtractions, and perfections, leading to a particular sophisticated adaptation (as we see so many of them in the living world) of the organism, to be impossible. And, moreover, Darwins theory presupposes continued perfection (improvement) of organisms or their parts, giving them a higher chance of survival, and thus becoming relatively more numerous and thus being selected. But we know that, generally, primitive forms do as well in life as do their 'advanced' cousins. Of all recent groups the primitive forms of them are still living today. But if one nevertheless sticks to perfection of organisms as being the main trait of evolution, we still have the mentioned interferences caused by the simultaneous presence of various different and competing selection pressures, and also have the expected fact that often certain directions, initially taken by the selection process, will, subsequently, change into some other direction and head to something else as a result of the action of the many abiotic and biotic factors in the environment. And these interferences and changes of direction must -- if one insists on the correctness of Darwinian theory -- somehow be neutralized, neutralized, that is, by some other natural factors. But this precisely presupposes t e l e o l o g y coming into play in natural events : A long continued series of precisely the right incremental evolutionary changes, eventually and definitely leading to the 'intended' sophisticated adaptation or life-form.
And because -- as is generally accepted -- teleology cannot play any role in the physico-chemical space-time world, sophisticated adaptations in organisms cannot be 'made' ('manufactured') in and by this world. And the reader already knows of course that that precisely was the reason for us to bring in the Implicate Order which is an immaterial but objective domain of Reality (of Being) that is in constant interaction (projection/injection) with the space-time world, the Explicate Order.
And all this is also precisely why we maintain that organic evolution, seen as a process of continued transformation of organic -- but still material -- entities [from one (adaptive) type into another], is impossible in the Explicate Order. And this means that organic evolution, as we 'experience' it (through theory), is in fact not such a series of material transformations, but nothing more than a sequence of appearances of new forms and new adaptations, and we wish to explain just this sequence. The material universe cannot, however, all by itself cause and sustain such a successive appearance of ever new organic forms. And this is indeed where the idea of the Implicate Order comes in.
Empirically, we know that there exists, in the realm of fossil and recent organisms, a certain degree of derivability of given forms from other forms. That's why we can do taxonomy and, especially, phylogenetic systematics. However, after all the efforts of evolutionary theory, one has not been able to produce a true and continuous phylogenetic tree of all organisms. The origin of many, including large, groups of organisms remains totally unclear because of the spectre of specialization-crossing and of the presence in today's fauna and flora of so-called primitive forms. To explain the latter phenomenon, one assumes this being the result of their continued persistence (already since very early times) whithout having ever been subjected to serious changes, while in fact natural selection should have had them replaced by their 'better' off-shoots already long ago. In other cases a given group may be in such a high degree idiosyncratic that its origin cannot be found. And this brings us back to our Hymenoptera (which form such an idiosyncratic group among the insects).
Their various strategies are, first of all, part of the noëtic strucure of the Implicate Order, that is, they are themselves noëtic entities (which only upon projection appear as organismic species). And they qualitatively happen to stand in some (branched) derivational order which is reflected in the Explicate Order in the form of organisms (organismic species) that display a certain degree of derivability of one from the other. So the actual sequence of appearance of organisms in the Explicate Order seems to show derivational order, making us to suppose an organic evolution to have taken place in that space-time world. But the actual sequence of appearance is a sequence, not of production in the Explicate Order, but of projection from the Implicate Order. And the projection of a given strategy depends on existing existential conditions in the Explicate Order. It does not exhaustively depend on the presence of previous organisms (i.e. previously projected strategies), 'ancestors', in the Explicate Order, although this does play a role, because earlier organisms take part in the constitution of existential conditions of other organisms.
Initially, as a result of the incessant injections (from the Explicate into the Implicate Order) the inorganic existential conditions become 'known' to the Implicate Order. Inorganic patterns (physical and chemical conditions) in the Explicate Order that are more or less stable, including all inorganic substances (in the metaphysical sense), will be injected (as 'copies' of their qualitative content) into the Implicate Order, and they will together settle there as noëtic patterns of actual or potential existential conditions, that is, qualitative patterns that can, in the form of their material counterparts in the Explicate Order, sustain the existence there of organic species. So these inorganic existential conditions, as they are noëtically present in the Implicate Order, 'cause' other noëtic elements to come together and 'crystallize' in them, i.e. to form intercalated new patterns, like the many different types of snow-crystals that are formed in the various existential conditions of the atmosphere, conditions, there, of humidity and temperature, in which water-molecules can come together to form intricate crystals. And when such incoming noëtic elements indeed crystallize to form 'crystals', they have, in the Implicate Order, become strategies, strategies, that is, to exist in the Explicate Order. And to exist in that Order is the ultimate form of existence : It is not only 'insistere' (to stay put in the Implicate Order), but 'exsistere' (to appear out in the Explicate Order). Every immaterial form ontologically aspires to exist in the highest degree, and that is to exist in the Explicate Order, because that brings with it its ontological completion. And depending on the conditions actually present in that Order, such strategies are actual or potential strategies. And, as has been explained earlier, a given strategy in the present sense is a noëtic prescription of how for this prescription itself (which is as such not more than just a noëtic entity) to exist and persist in the Explicate Order.
And of course, when, by the phenomenon of projection, organisms are being added (or others replaced) to the Explicate Order, the existential conditions change there and become more diverse and complex : Ecological conditions are developing in the Explicate Order, which themselves may form existential conditions for still other organisms not yet appeared. This is what takes place in the Explicate Order.
In the Implicate Order, of course, there is no such gradual diversification and complexification of the pattern of existential conditions, because this Order is timeless. When we say, while speaking about the Implicate Order, that in and by the 'initially' only inorganic existential conditions organic patterns crystallize (i.e. form strategies) in these conditions (which are in the Implicate Order their noëtic environment) and 'become' elements of existential conditions themselves that are 'added' to the 'already existing' set of inorganic conditions, while they themselves in turn let other crystals be formed as well, that are 'then' also added to the set of existential conditions 'so far formed' -- this is only meant in terms of analyzed complexity, not a complexity that is constructed by putting together parts, but a complexity that c o n s i s t s of (virtual) parts, without these parts having been there first. And this is what we also mean by "formed in the Implicate Order". That is to say, in so expressing states of affairs in the Implicate Order, we in fact analyze its complexity -- look to the virtual parts that make it up, and describe it as if it were successively built up by adding and substituting more and more parts.
Back to insects.
Each insect species (and each organic species for that matter) is the result of a projection, from the Implicate into the Explicate Order, of a strategy, itself formed in the Implicate Order. The strategy -- as a noëtic description -- has materialized. Such a strategy contains, first of all, elements that are not only necessary elements of this particular strategy, but also of those of a larger or smaller group of related strategies (existing in the Explicate Order as organismic species). Some such elements are even common, and necessary, to all strategies.
In the insect group Pterygota (primarely winged insects) the possession of functional wings is of course such a common and necessary element (of the materialized strategy). But what about the wing-venation? The presence of veins at all is also, of course, such a necessary element of the strategies of all individuals and species of the Pterygota, because they strengthen the wing-membrane without making it too heavy for flight. So all these are among the common and necessary elements of all the strategies of the Pterygota.
But what about the actual pattern of wing-veins? At first we thought that any specific venational pattern, other than a homogeneous distribution of veins over the wing-membrane, is entirely functionless, that is, it is only a formal feature. This might indeed be true, but there are some indications that this is not so : First, the many cases of high elaborateness (subtle structure) of many existing venational patterns, and, second, the persistence of a particular venational ground-work, not only such a ground-work for all insects, but also the existence of special ground-works each of them typical to each insect Order of the Pterygota. While the first of these indications (the high degree of elaborateness) is the most important one to bear witness of the functional status of the wing-venation, the second may just indicate the common descent of all Pterygota, by having inherited some non-functional feature (an thus indifferent to natural selection). Nevertheless the tenacious persistence of the mentioned venational ground-works might indicate that they are in some way shielded off from change because of their functional importance.
Indeed, we have seen that all venational patterns are based on a single scheme of principal veins, the comstock-needham scheme. And in addition we know that every Order of winged insects has its own specific venational prototype (itself still complying with the comstock-needham scheme) from which all existing venations of representatives of that Order can formally be derived.
In the Order Hymenoptera we have shown this in particular. And results like this, obtained from observations in the Explicate Order, have forced us to assume that in the noëtic formation (constitution) of a strategy, of what later is to become a species of the Pterygota, the wing-venation must be such as to adhere to the comstock-needham scheme, and to be derivable from the venational prototype of the insect Order concerned. We have not demanded that all wing-venations in such an insect Order must be derivable successively from each other, but only, and formally so, from the order-prototype. So when a given strategy of a pterygote insect is noëtically constituted in the Implicate Order (as described above) the particular wing-venation is a necessary but more or less commonly shared ingredient of that strategy, because it determines the particular fligh-regime of the insect, and this flight-regime co-determines how that insect precisely exists (i.e. finds itself) in its environment.
So if some given venational pattern satisfies the mentioned demands, and, in addition, does not in any way interfere with other elements of the strategy, it will as such automatically be added to this strategy. In this 'stage' the strategy and also the venational pattern are still purely noëtic, that is, immaterial. But upon projection we get a pterygote species of which the individuals possess the mentioned venational pattern. This pattern is more or less constant, and is in the Explicate Order only subjected to relatively minor changes, as described earlier. So although the venations existing in a given insect Order can all be derived from the order-prototype, and a number of them can even be derived from each other, they have not actually originated from each other but are, each one of them, in the Implicate Order assigned to the respective strategies.
But what about especially the h y m e n o p t e r o u s venational prototype? Although it does comply with the comstock-needham scheme of principal wing-veins of all insects, there exists a great morphological gap between that scheme and the hymenopterous prototype. In the fossil record, as well as in the wings of still existing primitive (generalized) holometabolic or other primitive insects, one cannot find transitional venational patterns that mediate between the wings of these primitive insects and those of the hymenopterous prototype (or of living or fossil primitive hymenoptera for that matter). So although this prototype can itself formally be derived from the general scheme of principal veins, it seems to have actually appeared suddenly, without direct morphological precursors.
For standard evolutionary theory this is really a problem, and when it indeed cannot be solved by it (as it is so unsolvable in many other cases as well) there is something wrong with this theory. For our noëtic theory of evolution, on the other hand, the morphological uniqueness of hymenopterous wing-venation is not a problem at all : It is part of some commonly-shared strategy-aspect, commonly shared by all Hymenoptera (that is, all Hymenoptera have a venation that can be derived from that prototype), and as such constituted in the Implicate Order.
Conclusion of Hymenoptera
In all the documents on Hymenoptera preceding the last three on fossil Hymenoptera, we have treated the origin and subsequent evolution of this Order in terms of the development of instincts. Indeed, the evolution of instincts, eventually resulting in the highest forms of instincts of providing for the young, is the chief characteristic of this Order. To provide for the young is the main activity of the adult insect (that is, the female, while the male only plays a role in the fertilization of the female, which activity is, indeed, also part of "providing for the young"). So the chief part of what a given hymenopteron essentially is, is its instinct-driven activity to provide for the young, and any theory of their phylogeny must mainly be based on the development of these instincts. And this we have done by following Malyshev's book (1966) on hymenopterous evolution.
In the three Parts concluding our treatment of the Order Hymenoptera, and dealing with the wing-venation of fossil and recent Hymenoptera, we have inspected these venations not chiefly in order to reveal the phylogeny of the Order but inspected them from a structural point of view.
In the next document we will return to the Order Diptera. While having in earlier documents most of our attention devoted to the evolution of them (mainly of the suborder Nematocera), by basing ourselves on their ecological setting, and also by studying the wing-venation of fossil and recent Diptera, we will now concentrate on their wing-venation again but now purely from a structural point of view as we have just done for Hymenoptera. A venational prototype of Diptera has been established by Hennig (1954, 1969), and we will check whether and how most of the venational patterns found in Diptera can indeed be formally derived from this prototype.
While we have seen that in Hymenoptera the wings and their venation are very uniform indeed (also in all fossil representatives of them), we will see that in Diptera there is much more venational diversity. Of course the dipterous venational prototype is very different from that of Hymenoptera (but still complying with the comstock-needham scheme of principal veins), but this cannot be the reason for the mentioned venational diversity in Diptera. The reason for this diversity must be sought in their being real flyers. They have attained the full "dipterigia", that is, they did not, as the Hymenoptera have done, merely decrease the size of the hindwings and functionally integrate them with the forewings (as if they now constituted the expanded anal or jugal area of them), forming one functional whole, that is, an intermediate stage of dipterigia, - but they had reduced the hindwings completely and had them turned into "halteres" that probably assist in positioning the body of the insect during flight.
And also in Diptera the various wing-venations cannot in all cases be derived from each other, but they must be derivable from the order-prototype. If that is so, then also here can we hold that the different venations in Diptera did not evolve from each other, but were assigned to the respective strategies in the Implicate Order.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXIX.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII