
Transition from saprophagy to phytophagy. Recent relicts of the Ectophytic saw-fly phase of hymenopterous evolution.
It is very important to clarify to what extent the just (previous document) obtained results are in line with the facts observed in the life and behavior of recent Hymenoptera. We should note that in this regard at first we see, not so much of a similarity, but a difference. Indeed, while the supposed mecopteroid ancestors of the Hymenoptera laid their eggs in cracks of the [woodland] soil [forest floor], that is, essentially on some non-nutritive substrate, and their larvae, as a result, had to live a wandering life and having to find food by themselves, we do not know of such 'geobiontic', soil-dwelling, and wandering larvae in Hymenoptera. Precisely the Hymenoptera (having in mind only the most primitive representatives of them) are "phytobionts", always laying their eggs on some nutritive substrate, and thereby chiefly on seed-plants and only sometimes on ferns and horsetails, which are then eaten by the emerging larvae. In full agreement with this difference the first mentioned larvae (those of the ancestors of the Hymenoptera) are saprophagous, while the secondly mentioned (the primitive Hymenoptera) are phytophagous, feeding exclusively on living plants. But with this great difference existing between the supposed ancestors and their descendants remarkable traits of similarity can nevertheless be observed, which we will now consider.
First we have the question asking in what way primitive phytophagous Hymenoptera could have arisen from their supposed ancestors that fed on disintegrating organic materials, that is, along what way our saprophags turned into phytophags. In order to provide a correct answer to this question it is necessary that we consider the conditions under which this transition took place. As is known, the floor of the damp hot forests at the end of the Carboniferous and the beginning of the Permian was covered not only with disintegrated remains of various fern-like plants and of gymnosperms, but also with a mass of fresh spores and pollen, fallen in plenty from them to the ground and in places forming whole layers. This mass of pollen, being more nutritive than just the tissue of the plants having produced the pollen, and being more easily to ingest with the help of even a weakly developed mouth apparatus, could easily become the chief food of such larvae living in forrest litter, and this food not only as a mass of pollen in a state of disintegration, but even in a fresh condition directly having scattered from the plants.
To convince oneself of the genuine possibility of such a transition, Malyshev placed young larvae of the common scorpion-fly (Panorpa communis L.) in a Koch's dish partly filled with damp soil on which fresh pollen obtained from male pinecones of the Crimea pine-tree were scattered. In another similar experiment, not larvae, but eggs of the same species were placed in a Koch's dish, (these eggs) covered with moss [and also provided with pollen, I assume]. In both cases the young larvae of Panorpa quickly found the pieces of the pollen mass provided for them, settling around them, entering in them their head, and fed on them. And indeed, in this way, we might say, the saprophagous larvae instantly became phytophagous.
Let us now turn to the habits of the most primitive Hymenoptera. In the Triassic deposits of Queensland (Australia) a saw-fly was found, Archexyela crosbyi Riek, and in the lower Jurassic -- Liadoxyela praecok Mart. [Later more fossil Hymenoptera were found in layers deposited in those periods. We shall deal with them later]. These are the oldest fossil Hymenoptera known, and with it they are representatives of precisely the primitive family Xyelidae. See next Figure.
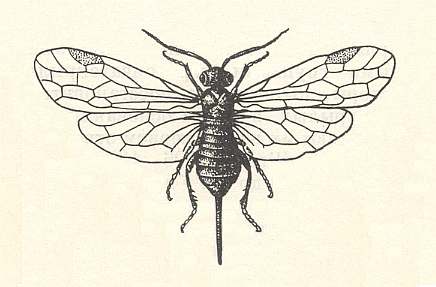
Let it be known to us that the larvae of the recent Xyelidae develop on flower pollen in pine cones. This coincidence in the feeding of the larval states of the Xyelidae on pollen of gymnosperms and that of their supposed mecopteroid ancestors really is a remarkable fact. It throws light on events that took place, we must assume, many millions of years ago, namely about 200 million if we reckon from the beginning of Triassic times up to the present. In addition it is observed that also adults of certain Mecoptera feed on pollen of plants, as feed on flowers [on their pollen] sometimes also adults of Panorpa.
It is known that also the primitive moths of the family Micropterygidae (Lepidoptera), which also originated from the panorpoid complex (Mecopteroids), and, up to today having preserved chewing mouthparts [while all other Lepidoptera have suctorial mouthparts], feed on pollen of flowers. [In all this we should not confuse pollen of flowers and nectar of flowers].
It is further significant that all known larvae of Hymenoptera generally are little mobile and live a more or less settled, if not, a completely sessile life. It is remarkable that decreased mobility being characteristic of the larvae of the recent Hymenoptera appeared, as we saw, and with great definiteness, already in the very early stages of their evolutionary development, that is, already in their mecopteroid ancestors. Having thereby lost in a significant degree, and later also completely, the ability of active moving [around] corresponding to the need for food and self-preservation, the larvae of the Hymenoptera became in need for proper care of them by the indivuals that had generated them. Precisely in connection with this helplessness of the larvae of the Hymenoptera instincts were developed, which are the most clear and complex of all animals. However, the widely distributed and, we might say, steady appearance of decreased mobility of the larvae of the Hymenoptera at the very early stages of their development when the maternal instincts of the given species were not yet properly developed, appears yet unexpected [the appearance of decreased mobility of the larvae appears unexpected because the instincts to care for them had not yet fully developed] and premature, if we do not look at it [= the loss of mobility] as being simply inherited from the past [So only when the loss of mobility is found out to have been inherited from ancestors it is not unexpected or premature].
According to today's conception, when the insects with complete metamorphosis had originated and their adult stages adapted to a life in the open, their larvae continued to live hiddenly as they'd done before in cracks of the ground and of the forest litter. It is clear that in such ecological conditions in the case of saprophytic feeding being characteristic of the larvae they did not need to significantly moving around and therefore remained little mobile with weakly developed organs of locomotion and with a delicate overall integument. On the contrary, another group of primitive larvae having predatory habits (larvae of carabid beetles, Raphidioptera, and others) obtained, in similar early circumstances, other traits -- great mobility, sufficiently strong limbs and a more robust shell. Already basing ourselves on all this, we can judge that the evolution of Raphidioptera as predators must have taken, contrary to the supposition of A. V. Martynov, a totally different course, and therefore the Raphidioptera had no connection with the evolution of the Hymenoptera.
We must also note here the fact that those larvae of recent Hymenoptera that live in the open are found essentially only among saw-flies (Megalodontoidea and Tenthredinoidea [both belonging to the Suborder Symphyta] ). This fact cannot be interpreted as being coincidental or secondary because it is connected with a series of other comparatively primitive features in the organization of the adult saw-flies (chewing mouthparts of the "orthopteroid" type, widely 'sitting' abdomen [= attached to the thorax not through a narrowing or petiole], rich wing-venation, dual trochanters, etc.) as well as of their larvae (a developed head with a pair of single eyes, thoracic and even abdominal legs, etc.). Also the embryological data indicate the more primitive status of the saw-flies as compared with the rest of the groups of the Hymenoptera. It is not without reason that the various authors, trying to set up a genealogic tree of the Hymenoptera, place at the very base of it the saw-flies, or placing them in the first branch of it at the tree's very beginning. From a biological viewpoint such an assessment is correct, and even the only one possible. Here, in the saw-flies, the maternal instinct is generally expressed only as the ability to find the appropriate nutritive substrate for the larvae on which the saw-fly then lays its eggs. This ability is widely distributed among insects, and the corresponding instinct does not need to be investigated further here [It is part of what we had called the p a s s i v e strategy, that is, just an adaptation to something already existing (i.e. the plants on which the larvae may feed)].
In connection with this one should note that feeding of the adults on pollen, of which we spoke earlier, in all probability was the stimulus that encouraged the females of the Xyelidae to lay their eggs on the strobilae [i.e. the pine cones] of pine trees, that is to say, in essence right on the substrate on which they themselves were able to feed. And this was evidently the particular stage from which the development of the Order Hymenoptera began. After the transition from saprophagous soil-dwellers to phytophagous pollen-eaters, the essential, albeit merely external and little substantial, transition from the original phase to further development of the life of the Hymenoptera took place when their primitive larvae began to specialize on eating the juicy and green parts of the plants as do the caterpillars of many Lepidoptera [butterflies and moths]. And then, in connection with this specialization of the larvae of the Hymenoptera (the first pseudo-caterpillars), or even anticipating this specialization, the adults of them began to lay their eggs on precisely that green substrate, that now began to serve as food for the larvae and on which the latter could freely move about.
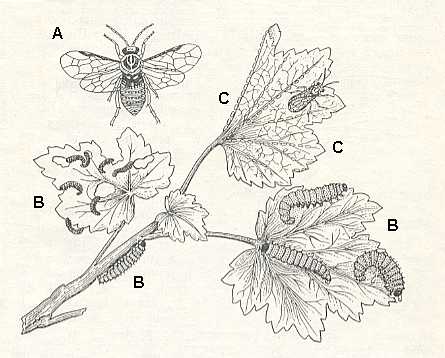
This transition of the primitive saw-flies from laying their eggs on organs that produce pollen to the neighbouring green parts of the plants was realized apparently without special difficulties. At least the larvae of certain primitive saw-flies of the family Pamphiliidae (see next Figure) still continue eating pine needles, while other representatives of the family began to develop already on leaves of trees and bushes of angiosperms, chiefly of the family of rose plants (Rosaceae).

It is remarkable that among the family Pamphiliidae (as also among Argidae and Siricidae) the genera connected with pine trees are characterized by more primitive morphological features than those that are connected with angiosperms.
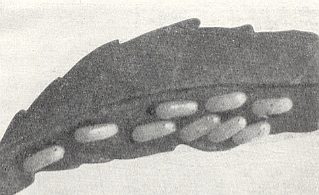
Figure 3 : Eggs of the primitive saw-fly Neurotoma memoralis L. (Pamphiliidae) on the underside of a young leaf of the bean plant Amygdalus nana L.
(After MALYSHEV, 1966)
Later, evidently, in order to obtain a better protection of the eggs from dehydration (bulging of the eggs of saw-flies!) and other detrimental factors the laying of the eggs became preceded by a special operation : injury of the delicate tissues of the plants with the help of the ovipositor (the 'saw'), a phenomenon that is shared by almost all saw-flies. In the simplest case the injury has the character of a light incision, while in more complex cases, for example in the cherry saw-fly or the great birch saw-fly, the skin of the plant (the epidermis of the leaf) is incised such that underneath it a, as it were, deep pocket is formed, in which the egg is put. The opening, which was formed at the place of the sting, is closed by a special secretion (produced by the external genital glands), in this way protecting the wound from infection by fungi. See next Figure.
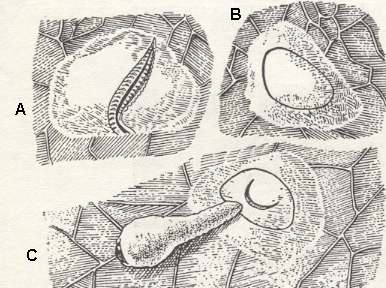
Figure 4 : Laying of the egg and the emergence of the larvae of the cherry saw-fly Caliroa limacina Ret.
A - End of the ovipositor of the saw-fly, incising the epidermis of a leaf.
B - Under the incised epidermis the egg shows through.
C - Emergence of the newly-born larva.
(After SANDERSEN, 1921, from MALYSHEV, 1966)
Nevertheless, the larvae emerging from such eggs, themselves submerged in the tissue of the plant, crawled out into the open and continued to live on the surface of the plants as it is usually still observed today in saw-flies. See next Figure.
They slowly moved, as the need for food demanded it, on the pine needles and leaves, while at times of rest assuming characteristic, diversely curved positions. Some of them, in the course of time, lost also this reduced mobility, becoming almost sessile. It is sufficient to point, for example, to the larva -- looking like a slug -- of the cherry saw-fly (Caliroa limacina R.) or the extremely flattened -- looking like a worm -- larva of the alder flat saw-fly (Platycampus luridiventris Fall.).
HYLJAROV (1949) holds that the habit of living in the open of saw-flies [larvae] is secondary [i.e. a derived condition] which he bases on the fact that in them "also in the case of the larvae living in the open the eggs are laid into the tissue of the plants". We [Malyshev], however, see it the other way around. Precisely the most primitive saw-flies place the eggs not into the plants but on their surface. This, of course, indicates that the ectophytic way of life of the saw-flies is a primary [that is, an original, not derived] feature.
Among recent saw-flies special care of the offspring is shown by the Australian Perga lewisi Westw., as was earlier described by Lewis and later confirmed by Frogat. The female of this saw-fly makes with its ovipositor a double row of incisions into the upper surface of the leaves of the eucalyptus where then a double row of eggs is laid (up to 80). After that she does not leave the eggs but stays above them. Spreading out her legs widely, holding the wings in a half-spread position and with opened jaws, she, from time to time, produces weak buzzing sounds. She in this way watches over the eggs not only until the larvae hatch, but also further, until the larvae begin to feed. Such a kind of "brooding" all laid eggs together, not complexified later by acts of attending to each emerged larva individually, points to the comparatively primitive character of this sort of behavior. Indeed, the described maternal habits of Perga lewisi look very similar to those that are observed from time to time in lower organized insects with incomplete metamorphosis (Hemimetabola), as for example in mole crickets (Gryllotalpidae), some earwigs (Dermaptera) such as Labidura riparia Pall., bugs (Hemiptera), and, indeed, centipedes (Polydesmus). Consequently, the described habits of the females of the "saw-fly of Lewis" can be assessed as being a sort of atavism (a phylogenetic old trait appearing again later).
With respect to the feeding of the adult saw-flies, it is the case that some of them do not feed at all, others visit flowers accessible to them (Umbellifers, Euphorbia, and the like) from which they (also) eat flower pollen. Others, on the other hand, especially from the larger species (Tenthredo, Allantus, and others) are predators, they attack small insects and eat them. They catch and kill their prey with their strong jaws. Being carnivores in adult saw-flies consequently does not have, neither essentially, nor as form, any relationship with the feeding habits of their larvae which independently subsist on green parts of plants.
So we must hold that precisely the sawfly-like forms separated themselves from panorpoids (Mecoptera) as the first representatives of the whole order Hymenoptera. This, presumably, took place at the transition from carboniferous to permian times at the very beginning of the radiation of the panorpoid complex. In this early period of the development of the saw-flies foodplants for their larvae were ferns, horsetails, and gymnosperms. Other foodplants for them did not yet exist. Concerning the vegetation of especially that period it is known that the gymnosperms were represented first of all by seed-ferns (Pteridospermae) -- large trees with strongly incised foliage with seeds (no spores) placed at the edges of their parts in a special cup made up of leaflets. The next group of gymnosperms are the Cordaitae -- trees with a mighty trunk and many branches. Their inflorescences [here, primitive reproductive organs, not true flowers] stood in the axils of leaves and remind us of catkins. Of recent plants they stand closest to gingkho and araucaria, these peculiar "pine trees" of the Southern Hemisphere. With respect to this it is interesting to note that also many larvae of recent saw-flies feed on such primitive plants as ferns, horsetails, pine trees, reeds, willows, birchs, and the like.
The saw-flies, already having early branched off from panorpoid ancestors, radiated [differentiated] such that some of them, the true saw-flies (Tenthredinoidea, or Strophandria), became specialized and did not preserve, as ROSS assumes, clearly archaic forms. Anyway, Ross, as he himself indicates, could not, because of shortage of material, investigate more precisely the interesting family Blasticotomidae which forms an archaic connective link between the true and lower saw-flies. With their anterior thoracic dorsal shield posteriorly deeply incised this family is close to the true saw-flies, but as to their wing-venation close to the lower saw-flies.
BENSON (1938) distinguishes 14 families of phytophagous Hymenoptera (Symphyta), presents a table to identify them and adds a series of essential notes. Pointing to the fact that the group is only little studied especially with respect to its exotic forms, he holds that it is, for example, possible that the Australian complex P e r g i d a e consists of 14 subfamilies, from which some, judging from a series of morphologicval features, turn out to be the most primitive of all saw-flies. Their still little studied larvae either have no abdominal legs (the genus Perga, see next Figure), or with pseudo-pods sometimes even on the first abdominal segment, as in Xyelidae.
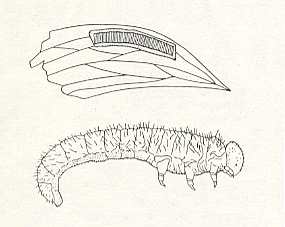
The second group, the lower saw-flies (Xyelidae and Pamphiliidae), preserved, as we saw, some very primitive traits. To this we can add that, for example, Megaxyela langstoni Ross. (from the Xyelidae) has the ability to actively move around in its pupal state. In this stage it crawls to the surface of the soil in order to cast off its pupal skin, and in this way showing the remarkable biological feature of their panorpoid ancestors (active pupa). On the other hand, representatives of the archaic family Xyelidae are characterized by features relating them also to the higher group (consisting) of the horntails [The 'spurious' horntails are the stem-horntails, the Cephidae, while the true horntails (trunk horntails) are the wood-boring Siricidae and allies.], forming with them, as it were, a single whole, the group Orthandria, according to Ross.
In the next document we will deal with the next phase, the Endophytic saw-fly (Cephoid) phase.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXXIII.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI