
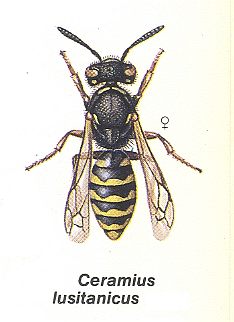
For a further understanding of the evolution of nesting instincts of the masarids (Masaridae), [nesting instincts] still more approaching to the apoine type, the observations of Ferton on the habits of Ceramius tuberculifer Gauss., about which we partly already have spoken earlier, are very instructive. In the Pyreneans (Monlouis) this masarid populates in colonies [aggregations] in hard clayey soil, covered with scarce grassy vegetation, and always in the neighborhood of water. The nearness of water turns out to be necessary to these wasps in order to soften with the help of droplets the hard earth in which they dig. In the second half of July, in which the observations were carried out, the work [of the wasps] was in full swing. Nearby the colony, counting many hundreds of individuals, a small basin of clear water was the place of their constant visiting. The number of individuals bringing-in water[-loads] simultaneously was on average 7-8, but flights-to and flights-from were observed to take place constantly. From the first portions of material obtained from digging out the burrow, an entrance-tube was constructed. It is built in determined order and renewed every time when damaged or destroyed. This tube can resist heavy rains lasting for several hours.
About the meaning and further destiny of this tube the investigator says nothing. Will it, perhaps, merge into the final closure of the nest, or is it, finally, stock-material for the construction of cells?
The cells of C. tuberculifer, lying horizontally in the depth of the burrow, are unified into one whole group. Their outer surface being rough but of regular form, shows individual pieces of cement of which it is made. But on the inside the walls of the cell are polished.
Before preparing provisions, this species deposits an egg in the cell, but the egg is not suspended on a thread as in the nests of Odynerus and Eumenes, but is provisionally deposited on the floor of the cell. The honey-dough having a firm consistency is formed as a pie recalling the shape of a short-necked retort with a widened apical neck. See next Figure.
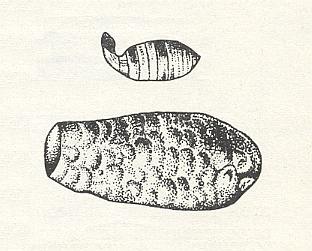
The next Figure depicts a species of the genus Ceramius :
The mother-wasp sticks the egg, laid earlier, onto this projection, with which the just hatched larva starts its feeding. The successive loads of provisions, brought to the cell by the mother, are not mixed up and remain separate from each other, the one following upon the other in a horizontal direction. Having finished provisioning of the cell, the mother closes it with mineral cement. Later, the whole group of constructed and provisioned cells is closed with a common cover of cement.
There exists an indication (Williams, 1927), based on very succinct and incomplete data, according to which the small masarid Euparagia scutellaris Cress. stocks its cell with larvae of snout-beetles. Meanwhile, in the complex natural conditions, it is, without opening up the cells-to-be-provisioned, not possible to hold that precisely the mentioned masarid, and not some other wasp has hunted in the neighborhood after snout-beetle larvae.
Similar assertions have been made for Pseudomasaris, allegedly provisioning their cells with caterpillars. Today it is definitely known that also these masarids do not form an exception, they also provision their cells with honey-food.
In the considered behavioral features of the solitary plaited-winged wasps (Eumenidae and Masaridae) we should see the demonstration of an unusual evolutionary plasticity of them. On the basis of this plasticity, as it was already said earlier, lies the cancellation of certain acts from the chain of serial acts, and the subsequent coupling together of the remaining ones. Such a kind of transformation of instincts, not having demanded essential structural changes, in many cases turned out to be very effective as to their results. Such a simple method of transformation of maternal instincts in wasps is undoubtedly expressed in the acceleration of their changes. This also already follows from the very diversity of maternal instincts in the individual wasp-genera Ammophila, Odynerus, Synagris, and many others, dealt with earlier.
While until now we considered chiefly the rearing-instincts of wasps, it is now necessary to adress our attention to the development of their building-activity. The first stages of it were given earlier. In its simplest form it was expressed in adapting an existing ready-made cavity in behalf of a cell, and especially in the ability to construct an opening in the cells. Working in conditions of insufficient moisture (on the sunny side of clayey slopes, and so on), the wasps had, and, evidently, quickly, to acquire the ability to bring water to the construction site, in order to moisten with it the dry building-material and prepare out of it a plastic mass. This also gave them the possibility to use the latter not only for closing the openings of the cells, but also for covering with it the whole nest cavity, as for instance in Ancistrocerus callosus Thoms. See next Figure.

Figure 2 : Plastic [mouldable] clay, prepared by the wasp Ancistrocerus callosus Thoms., covers, from the inside, the walls of the cells from all sides. (After MALYSHEV, 1966)
As a result, the ability to built roofs of the cells was transformed into the ability to model cells, as this already appeared among odyners (Odynerus laevipes Shuk.) and especially in masarids.
Having now considered apoine behavioral elements in certain vespoid wasps, we will, in the next document turn to sphecoid wasps, and see where, in them, apoine behavioral elements may be found.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LXIV.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII