
Organic Evolution in terms of the Implicate and Explicate Orders.
Part LXIV
Hymenoptera (wasps, bees, ants) (Sequel)
The evolutionary diversification in the Order Hymenoptera in terms of Strategies (Sequel).
Evolutionary establisment of the Bees (Apoidea)
Evolution of the Sphecoid Wasps and the question of the (evolutionary) establishment of the true bees
Sphecoid wasps are, as to their morphological features, more specialized than vespoid wasps are. We can even say that in virtue of the structure of their thorax they are distinguished from all other Hymenoptera and come only close to true bees, as was already noted earlier.
Handlirsh (1906-1908) assumes that from both families, together named Sphegiformia, that is, bees (Apidae) and digger-wasps (Sphegidae) [also called Sphecidae], the first, evidently, originated from one not yet highly specialized member of the latter. The fact that in this series still kidney-shaped eyes are sometimes found, but not plaited fore-wings (for example in Philanthus and Trypoxylon), points to the origin of sphecoid wasps from scolia-like ancestors, or from lower not yet plaited-winged vespoid wasps, in which the first alternative is taken by the author to be the more probable. For good orientation, we, in the next three Figures, give examples of the wasp families Scoliidae, Sphecidae, and Vespidae, and in one more Figure a representative of the true bees (Apoidea).
For a wasp of the genus Scolia, see next Figure.

Figure 1 : Scolia maculata, Family Scoliidae, Dagger-wasps, Suborder Aculeata. 20-40 mm.
(After SEVERA, in ZAHRADNIK, Thieme's insektengids)
For a typical representative of the sphecoid wasps, see next Figure.

Figure 2 : Ammophila sabulosa, Family Sphecidae, Digger-wasps, Suborder Aculeata. 16-28 mm.
(After SEVERA, in ZAHRADNIK, Thieme's insektengids)
For a typical (social) plaited-winged wasp see next Figure.

Figure 3 : Polistes nimpha, Family Vespidae, Social Plaited-winged wasps, Suborder Aculeata. 12-12.5 mm.
(After SEVERA, in ZAHRADNIK, Thieme's insektengids)
For a typical (solitary) (true) bee, see next Figure.

Figure 4 : Halictus quadricinctus, Superfamily Apoidea, Family Halictidae. Social bees, Suborder Aculeata. 15-16 mm.
(After SEVERA, in ZAHRADNIK, Thieme's insektengids)
Thus, according to Handlirsch, on the basis of morphological data, we must depart from the fact that the vespoid wasps (to which also the scoliids belong) are more ancient than the sphecoids, and that the sphecoid wasps, and with them, consequently, also the true bees, had their beginnings from them. From the biological viewpoint these relationships are fully confirmed.
Indeed, in the first typically vespine phase ( Primary-vespine (pompiloid) Phase), in which hunting after a prey still took place before the [evolutionary advent of the] ability of building nests, vespoid wasps (especially scoliids and pompilids) were in great development, whereas sphecoid wasps were still weakly represented as if they were mere exceptions.
In the next Secondary-vespine (sphecoid) Phase, in which, on the contrary, construction of the cell preceded the hunt after prey -- also still one prey for one larva each -- we find vespoids (only pompilids) as well as sphecoid wasps. However, all of them are represented here by only a small number of forms, because at the very base of this phase of development elements were included for its subsequent rapid and fundamental transformation.
In the two subsequent phases, in the 3rd (crabroid) and 4th (bembecoid), in which into the earlier prepared cell were already brought in, not one, but several preys, either massively by the accelerated method (3rd phase, Third vespine (crabroid) Phase ), or, on the contrary, decelerated (4th phase, Fourth vespine (bembecoid) Phase ), we see only sphecoid wasps, represented here in unusual diversity and in an abundance of forms and habits. The evolutionary line of development of the vespoids is here as it were lost, and this means that we must find its traces. And such traces there undoubtedly were. This is already evident from the fact that they anew and clearly appeared in the fifth phase ( Fifth vespine (moneduloid) Phase ), again next to the sphecoid wasps in that remarkable form of having the egg deposited on the wall of the still empty cell and the prey brought in later according to need.
In this phase the sphecoid wasps reached, apparently, the limits of their development, whereas the vespoids came out in new forms as the very wasps -- Diploptera (plaited-winged wasps). Only as an anomaly or atavism sometimes appear, among these "true" wasps, habits of a lower order, characteristic of the 4th or even of the 3rd phase, as for instance the bringing-in into the cell of the first prey (caterpillar) before laying an egg onto it, about which we spoke earlier. [In what follows, these evolutionary phases (1st, 2nd, 3rd, 4th, and 5th) are often referred to, and the reader may consult the documents treating them by means of the above LINKS (Primary, Secondary, Third, Fourth, and Fifth vespine phases)]
The clarification of the pathways and conditions of development of the instincts of wasps that lay their egg in the still empty cell (5th phase), showed that the instincts of this type could only originate from the ground of earlier formed instincts of provisioning of the cell with several preys and then the bringing-in of prey into the cell in a slowed-down rate. There is no basis for connecting the high-level instincts of the 5th (moneduloid) phase directly with the primitive instincts of [those] wasps that prepare only one prey for each cell (1st and 2nd phases). Therefore we must conclude that also vespoid wasps, before going into the 5th (moneduloid) phase in the form of already true plaited-winged wasps, had to go through precisely those two previous phases that were seen in sphecoid wasps -- the 3rd (crabroid) and the 4th (bembecoid).
We must take into account that the life-history of a multitude of diverse wasps still remains unstudied, and this not only is the case with individual species and genera of them, but with whole tribes [tribi] and even families, as for instance the vespoid wasps of the family Fedtschenkiidae, of which the life-history [way of life] is completely unknown. We may therefore rightfully expect that many important gaps in our knowledge will be eventually filled up here. But also now we may already guess among which groups we must look for the missing links, and by which habits, generally, they should distinguish themselves.
Thus, the sphecoid wasps went, in the development of their instincts, together or parallel with the vespoids, through the successive series of phases beginning with the primary-vespine (pompiloid) phase and all the way to the 5th (moneduloid) phase. But further than this point, to the 6th phase ( Newly vespine (vespoid) Phase ), the sphecoid wasps apparently did not go. Meanwhile the vespoids passed over into that 6th newly vespine phase and here subsequently reached the highest development of vespine instincts -- the social way of life. In addition, they gave also rise to apoine habits of the vespoid type (masarids). At the base of these two biological features of vespoid wasps there was that very one moment -- the transition of the vespoid wasps, in the beginning of the newly vespine phase, from the original habit of feeding the larvae with a whole prey to bringing it to them in a chewed form.
Now we have the question of where to find those forms of sphecoid wasps which, following the example of certain vespoid wasps, also had to pass through the 6th (newly vespine) phase of development and making here their own the habit of feeding the larvae with a chewed prey. After all, precisely this was the basic condition for the origin of apoine life among vespoid wasps.
From the circle of sphecoid wasps, standing in the previous, 5th, vespine (moneduloid) phase, we know representatives of, all in all, two closely related families -- Stizidae and Bembecidae. But they, with their prey being coated with hard chitin (adult cicadas, grass-bugs, grasshoppers, flies), ended up, as might be said, in a dead-end line and could not develop further. Anyway, data on the habits of many bembecoid wasps are still very insufficient or lacking completely. Thus, almost nothing is known about the life of the peculiar Australian Exeirini, standing close to the Stizini. This tribe is represented by, all in all, one genus with only one known species Exeirus lateritius Shuch., hunting for cicadas.
It is therefore really remarkable that sphecoid wasps of the genus Microbembex also carry sometimes a prey not in one piece, but in individual chunks, now and then, it is true, too hard for them to be fed to the larvae, -- in the form of individual legs, skins shed in molting, and so on. See next Figure.
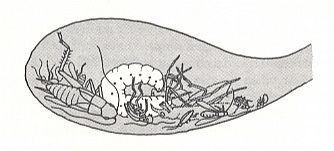
Figure 5 : A larva of Microbembex monodonta (Superfamily Sphecoidea, Fossorial wasps, Suborder Aculeata) in the nest cell, feeding on a variety of dead arthropods (drawn from a photograph). (After EVANS and EBERHARD, 1973)
One gets the impression that, having reached the highest stage of development, the sphecoid wasps also made an attempt to pass over to feeding the larvae with a reduced (i.e. bitten into pieces) prey, but this attempt turned out to be unsuccessful for them : they did not have a delicate prey suitable to being chewed, after which precisely the vespoid wasps hunt, having made their own a new method -- offering the larva a chewed prey.
Thus, based on the available biological data we may tentatively say that of all sphecoid wasps the highest stage of development was reached by the wasps belonging to the genus Microbembex. It is therefore interesting to analyse their habits more closely. First of all we note that the very name "Microbembex" is biologically not so appropriate. Already in the flight of the microbembexes there is no such intense striving with which bembexes [that is, representatives of the genus Bembex] hunt for their fast-flying prey -- horse-flies, tachinid flies, bee-flies, and so on. On the contrary, according to observations of Hartman, 1905, M. monodonta Say. can be seen quietly flying between trees, with its flight reminding of a dragonfly, stubbornly keeping one height, about a foot above the ground. It further turned out that microbembex does not hunt for anything specific but only collects or picks up from the ground objects deemed useful to it. But what are these objects? Stoehr, 1917, observing in Canada the work of M. monodonta, very lively describes the amazement of the investigator after having seen that instead of the usual prey as it is in wasps, M. monodonta carries dead bodies of the most diverse insects and even their parts -- legs, shed skins, and the like. Also the married couple Rau, 1918, having, in the state of Kansas, dug out burrows of M. monodonta, found, as to their astonishment, in the cells of the wasp two different beetles, two cicadas, one spider, and one ant. "All these six prey-objects were dead, dry, and hard. Anyway, we hardly can call this collection "prey", because this is just a collection of dead remains gathered by the wasp."
Searching the surface of the ground for motionlessly lying insects, their parts, and in case also spiders, the wasp, apparently, without special selection, collects totally dried-out remains of them as well as those that have retained some degree of juiciness and freshness. When the investigator throwed at the wasp close to its nest a freshly-killed grasshopper, she also picked it up, as she after it also picked up a dry hover-fly thrown at her.
If we take into acount the fact that M. monodonta is active in the hot period of the year in dry sandy places, it becomes clear that in its searches, as direct observations indicate, it encounters mainly already totally dried-out dead bodies of insects especially of those that are common in those places. Naturally not excluded is the possibility of finding in these conditions not only still completely fresh, recently died insects, but also those that were recently paralyzed by other wasps and temporarily left behind on the ground or totally abandoned, as it not seldom occurs in ammophilas hunting for caterpillars or in pompilids hunting for spiders. In precisely this context, apparently, we must understand the bringing-in to the nests by different individuals of M. monodonta of five slender colored caterpillars, reported by Hartman. In connection with this we must note that, according to observations of another investigator (Stoehr), microbembex carries sometimes totally dried-up "mummies" of caterpillars of Geometridae (Carpets, Waves, Pugs, etc.).
Hartman, the first one to observe the behavior of these wasps, supposes that they also may attack living insects. He bases this on the fact that he once had the occasion to see a struggle between a microbembex and a large fierce ant, Pogonomyrmex apache Wheel. But this argument is inconvincing, first of all because in this case it is not known who attacked who first, and secondly, in the struggle the wasp was defeated -- an event altogether little probable in wasps hunting for their prey.
Thus we get the impression that in the microbembexes the hunting habits of wasps were totally atrophied, and only in the form of a rudiment of these habits still the instinct of collecting dead bodies of formerly being preys, or of [collecting] what is in a certain degree similar to that prey and its parts, was preserved. The picked-up remains of this prey Microbembex monodonta carries to its burrow which terminates in, all in all, one single cell -- a simple widening at the end of the burrow.
But what happens in the cells of the microbembex? The strange habit of these wasps outside the nest is still more puzzling inside it, in the cell. One of the mentioned investigators of the biology of M. monodonta, Stoehr, openly declares that with respect to feeding the larva of microbembex he is left with total unclarity and even holds that it is impossible to understand for what, to the wasp, serves its dry mummified prey. Digging out by him five nests of microbembex did not provide an answer to the question. Thus, in one nest were found the remains of, all in all, three ants (of them one "very big"), and in another nest -- pieces of 4-5 ants and one beetle. By what then did develop the larvae in these nests?! About this the investigator left it to judge by his readers! Also Hartman did not report anything about the egg and larva of Microbembex, and only supposes that a fresh juicy caterpillar (which was referred to, -- C.M), placed together with old dry insects, must be indeed a nice bite for the growing larva. To this we can add that also the Rau's failed to find in the by them opened to be provisioned cell of M. monodonta neither an egg nor a larva of the wasp.
Parker, 1917, on the other hand, studying in the state of Ohio during six weeks the nesting behavior of M. monodonta, gave a series of essential data about the relationship between the sexes, the digging of burrows, the finding of nests, and so on, but chiefly clarifying the question not in those respects that are considered right now. According to Parker, microbembex, having digged a burrow to a depth of 8-12 cm, lays an egg in its only one posterior widening -- the brood chamber (cell). The egg is firmly fixed in a vertical position in the sand at the bottom side in the depth of the cell. It develops in 2-3 days, and in this time no food is brought into the cell. The wasp feeds its larva with dead insects, which it collects at the surface of the sand instead of catching and paralyzing with its sting living insects as do other wasps. Therefore, the food collected is very diverse, but mayflies and midges (Chironomidae) are taken most, as a result of their abundance at the work place.
It is not a single time observed that the wasp attacked living insects. She eagerly took mayflies just killed and thrown to her by the investigator at the entrance of the nest, but always was frightened when the wounded mayfly could still move its legs or wings, and did not touch it. Having taken a dead insect, the microbembex bent its abdomen and, as it seemed, made an attempt to sting it. But that the wasp also did with dry insects. Thus, the reflex of stinging the prey was still preseved in microbembex but had lost its significance. It was noted that the wasp does not provision the cell massively, but provisions the nest bit by bit during at least two or even several days.
The investigator concentrated his effords to feeding larvae of microbembex in artificial conditions. The obtained data were, according to his own opinion, far from satisfactory. But nevertheless some larvae, taken from cells, in various developmental stages, managed to reach that stage in which they prepare a cocoon, and two of them did make it even from egg to fully-grown larva. These larvae were held each of them in a separate test-tube, closed with a plug. They were fed with very delicate [soft] freshly-killed insects, mainly mayflies and midges. Rearing of the larvae took 5-6 days, apparently faster than in natural conditions, because, as the investigator assumes, in the artificial conditions they were given more delicate food.
Unfortunately, about the very technique of offering food to the young, especially to the newly-born larvae (hatching from the egg, placed not on the prey!), the investigator is silent. From such experiments we may only draw the conclusion saying that although the wasp itself had lost the hunting instincts, her larva still had preserved the ability to eat delicate freshly-killed prey, if, of course, this food was given to it in an appropriate way.
But how will feeding of the larva take place in that case in which it, in its cell, is brought totally dried-up and hard prey? In other words, on what does the larva of the microbembex feed, when the mother-wasp gives it legs of grasshoppers, heads of ants, skeletons of snout-beetles, thoracic armour of mutillids, and similar objects, in which hardly the needle of the entomologist goes in? This question is not asked by Parker. But for us it is very interesting. It would be useful to carry out experiments directly contrasting with that of Parker. Instead of freshly-slain soft prey, after which the microbembex itself does not hunt and which its larva hardly could have in natural conditions, it would be interesting throwing to the microbembex at the entrance of its nest precisely that what the wasp itself eagerly collects, but what is, as might be held, totally unsuited to feed her larvae with. We already had pointed to the cells of Microbembex monodonta with a similar content.
In the absence of corresponding experiments and direct observations of the nutrition of the larvae in such cells, we can only make one guess : When the mother-wasp carries in her legs precisely that what cannot serve as food for her larva, then she must bring it food in another way, inside herself, and giving it off through the mouth. But what then is this food? Hartman once observed that the female M. monodonta worked, alternately with her jaws and snout, the thorax of a dead hover-fly, and assumes that here the wasp undoubtedly fed on the sap of the fly. But to this is added that the majority of solitary wasps sucks nectar of flowers. In the other case a dry mutillid (antwasp) was partitioned by the wasp into two parts, both of them carried separately. Stoehr also observed a microbembex having bitten-up a dry grasshopper, and in this way having reduced its size, that is, making it more portable, easy for transport. He also holds that this wasp is not carnivorous and has to feed itself with nectar. And thus, a mixture of nectar and saliva of the wasp, and in cases perhaps also [mixed] with sap of some not yet totally dried-out finding, -- this then is all what the microbembex may bring to the mouth of its larva. Systematic investigations in nature should have to clarify this.
In concluding the analysis of available data on the nesting instincts of M. monodonta we must still recall the observations of Claude-Joseph (see Primary-apoine Phase of the Vespoid Type) in Chile on another species of microbembex, M. sulfurea Spin. These observations also have an interrupted character, whereby the author of them does not refer to any of his predecessors having studied the ways of M. monodonta, these ways then, evidently, remaining unknown to him.
It turns out that M. sulfurea Spin., also populating in sandy places, constructs in its burrow not one, but several cells, which, from the viewpoint of the author of these lines, is, with respect to bembecoid wasps, more primitive. The egg of M. sulfurea "is placed in the depth of the cell, fixed upright on one of its poles among the sand particles". At the side of the newly-born larva, in one of the two observed cases, "as a first prey, there was a paralyzed harvest spider (Galeodes variegata). The other larva had a paralyzed fly and a lady beetle". Indeed, was this prey really not dead but paralyzed, and paralyzed precisely by whom -- this is not asked by the investigator. In two months, when the nesting of Microbembex sulfurea was already completely concluded, a dozen cells were dug out. In the analyzed remains almost all main orders of insects were represented -- Diptera, Hymenoptera, dragonflies, cicadas, beetles (of them lady beetles, leaf-eaters, carabids, lamellicornids, and gyrinids), and one harvest spider. We cannot hold that the wasp indeed had hunted for such diverse preys, living in the most different conditions, including water! Clearly, M. sulfurea, following the example of its northern cousin, also collects only dead bodies of insects, now and then already totally dry, and in cases perhaps also alien paralyzed prey. Accordingly, the data on M. sulfurea Spin. do not give us something essentially new, but only a certain confirmation of what we have understood from the study of the habits of M. monodonta.
Thus, we may be confident to say that wasps of the genus Microbembex indeed have reached the highest developmental phase among sphecoid wasps. This is expressed in a number of their biological features. To these first of all belong their loss of the hunting instincts and the replacement of them by the instinct of collectors [instead of hunters]. This remarkable trait makes Microbembex similar to the masarids (Masaridae) and brings them close to the sphecoid bees. With the loss of hunting instincts was connected also the loss of the instinct to sting the prey, of which was preserved but an unuseful rudiment -- only a mere reflex. Then follow the deposition of the egg in the still empty cell, step by step feeding of the larva, the bringing-in into the cell of dead bodies of the most diverse insects, and sometimes also of spiders, for a great part already totally dried-out, and not only [bringing them in] as whole bodies, but also in individual chunks, totally having lost any nutritive significance. It is very well possible that this way of provisioning, representing a rudiment of once provisioning the cell with paralyzed prey, is now, in these new conditions, a prerequisite, stimulating the wasp to feed the larva with precisely that what she herself eats and which may be given off again out of the mouth to the larva, that is, that very moment in the evolution of sphecoid wasps at which they switched over to apoine habits and from which the sphecoid bees might have taken their origin.
It is interesting to clarify more closely the conditions in which the loss of hunting instincts took place in Microbembex. This could probably not have happened from the [back]ground of the use by them of a prey of other wasps and laying their own egg onto it. Such habits are known for a number of various sting-carrying Hymenoptera, including one American stiz -- Stizus unicinctus Say. This wasp penetrates into the nests of the sphex Priononyx thomae F., destroys the egg of the host-wasp deposited onto its prey (grasshoppers of the genus Melanoplus), and lays its own. The author of the present lines thinks that Stizoides tridentatus F, living in the USSR, has a similar relationship with Sphex maxillosus F. This wasp [stizoides] was encountered a number of times by the author (in the Chopersky nature reserve and at the shores of the Sula) precisely there where the mentioned sphex was nesting in colonies, and at the same time. But this is not a phenomenon of cleptoparasitism, but of inquilinism, not connected with the robbing of an alien prey and hiding it in a self-constructed nest.
Such relationships clearly point to a certain instability appearing in the hunting instincts of bembecoid wasps. Indeed, these instincts in the change of selection of prey and with that prey's extraordinary diversity, might become atrophied and transformed into the habit of collecting alien or totally prepared and dead prey, as it is observed in Microbembex.
In a final evaluation, taking into account the parallellism in the evolution of sphecoid and vespoid wasps, and also the fact that in the latter the habits of the apoine type originated only at their highest phase of their solitary life, it is not difficult to conclude that the nesting instincts of the true bees might have the same beginning, although direct concrete indications for it we do not yet possess.
In the maintained a step by step provisioning in a number of plaited-winged wasps of chewed [into pieces] food, carried in the mouth, the mother-wasp places the food directly into the mouth of the larva or toward its mouth onto its body. But such a way of feeding [the larva] is, with respect to sphecoid wasps, very doubtful. A for this moment preparatory feeding of the larva with chunks of a chewed prey did never occur in them. They only were in a position to carry inside themselves liquid or semi-liquid food, which they collected on flowers or swallowed from the crop of preys, and which they themselves ate. But this food, without special methods of offering could easily contaminate the body of the larva and kill it. In addition, if feeding from mouth to mouth once occurred in sphecoid wasps, it would sooner or later have led them to "trophallaxis" -- mutual feeding of larva and mother-wasp, in a strong degree conditioning the bloom of social life in plaited-winged wasps. However, trophallaxis does not only not occur in sphecoid wasps, but also not in sphecoid bees. Clearly, in that apoine line of evolution there were no conditions for its development, that is, in other words, feeding of the larva by the mother from mouth to mouth never has occurred here.
In this, apparently, obscure position we should remember the puzzling habits of microbembexes, bringing to their larvae dry chitinous remains totally unsuited for feeding, but perhaps useful to serve as substrate or some sort of "kitchenware" for liquid food, which the mother-wasp may pour out onto those remains from her mouth in the step by step provisioning of her larvae. A number of authors, having written about these strange habits did not make any statements as to their biological significance. Our guess is nevertheless in nothing improbable. The content of "prey" in these remarkable wasps has reached a limiting diversity. It is therefore natural that they could bring-in into the cell also an abundance of proper food, collected by them on flowers or sucked out of the body of preys, and give it off from the mouth. Such an abundance of food, able to be carried inside the organism of the mother-wasp, being of liquid or semi-liquid consistency, could be placed in the cell in the given conditions only onto secondary objects earlier or on the way brought into the cell in the form of dry skeleton-formations.
Because the bembecoid wasps of the genus Microbembex, bringing into the cell dry insects, feed the larvae with the "progressive method", from day to day, also the offering of honey-food, able to be given off from the mouth onto these remains, had to be realized, then, in a step by step fashion.
About the further development of such kinds of instincts as in the typical apoines [bees] we can, so far, only put forward vague guesses.
In the next document we will, still following MALYSHEV, 1966, discuss the origin of the Primary-apoine Phase of the Sphecoid Type, (this type) being the result of the transition from the sphecoid wasps to the sphecoid bees as to their instincts.
[This Phase is the last one that is discussed by Malyshev, that is, with it the exposition of the evolution of Hymenoptera as to their instincts is then concluded. The transition from solitary bees to social bees is not discussed. It probably has much in common with that same transition in wasps.]
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LXV.
Back to Homepage
Back to Contents
Back to Evolutionary Part XIV
Back to Evolutionary Part XV
Back to Evolutionary Part XVI
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XIX
Back to Evolutionary Part XX
Back to Evolutionary Part XXI
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XL
Back to Evolutionary Part XLI
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part L
Back to Evolutionary Part LI
Back to Evolutionary Part LII
Back to Evolutionary Part LIII
Back to Evolutionary Part LIV
Back to Evolutionary Part LV
Back to Evolutionary Part LVI
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LIX
Back to Evolutionary Part LXa
Back to Evolutionary Part LXb
Back to Evolutionary Part LXc
Back to Evolutionary Part LXI
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII