
In the previous document we once more concluded that random genetic mutations followed by natural selection of certain mutants, although at times occurring, cannot constitute the chief driving force of organic evolution. We found out that the genetic mutations that drive evolution must be non-random. But from this result we are not willing to conclude that there must be some transcendent deity directing and controlling organic evolution. Also non-random changes must have natural causes. But if we really want to account for the emergence of true novelty (which does happen all the time in evolution) we, as long as we remain in a purely physical context -- that is, Reality unfolded along space and time dimensions --, must invoke randomness : From an existing diversity of elements, a diversity having resulted from random variation, certain elements are -- depending on their fitness and stability -- selected (to continue their existence) while others are weeded out. But we had found out that in the case of organic evolution randomness cannot be the chief factor of the emergence of novelty. From this we must conclude that the mentioned context -- the purely physical context -- is too narrow, that is, our conception of Reality is too narrow. And indeed, to be able to account for the emergence of novelty, especially of functional novelty in organic evolution, that is, of the development of 'smart' adaptations in organisms invading new environments, we had presented our theory of Reality as being a dynamic between two Orders : (1) The Explicate Order of Reality in which things and processes are stretched out ('unfolded', 'explicated') along space and time dimensions, and (2) the Implicate Order in which things and processes are folded back ('enfolded', 'implicated') along those dimensions, resulting in them to be mere immaterial or mathematical entities in which space and time only appear as numerical parameters. And because of their immaterial nature we call them noëtic or thought-like entities. And while their material counterparts in the Explicate Order can under certain specific conditions physically, and especially, chemically, react with each other, giving rise to determined reaction-products, the noëtic entities in the Implicate Order can, depending on their respective qualitative contents, noëtically react with each other resulting in determined noëtic reaction products. These noëtic reactions and their noëtic products can, under certain conditions, be projected from the Implicate Order into the Explicate Order where they can be observed as material physico-chemical processes and their material products because they are now unfolded along space and time dimensions.
Precisely along these lines we will now describe the evolution of muscoid flies from protorhyphoid flies, but, unfortunately, compelled by necessity, in general terms only.
As in all organic evolution, also in this evolution the successive invasions of new ecological niches (and with them of new zones) determine its course. This demands a further exposition of the potential ecological niche of a given species of organism. The representatives of that given species live in the actual ecological niche of that species. They are completely adapted to this niche by having 'internalized' certain key features of the environment. All this applies to at least all animal species. Here we confine ourselves to species of Diptera, and moreover only to those belonging to precisely the families that are involved in the evolution of muscoid flies from (jurassic) protorhyphoid flies.
What then is the potential ecological niche of a given species of fly?
First of all we must realize that such a species is a definite qualitative content. In the Explicate Order this content is represented by a number of individual instances, that is, here-and-now existing individual flies (adults or larvae). The existence of these individual instances means that the mentioned qualitative content is unfolded along space and time dimensions, resulting in that qualitative content to be observable in the form of existing material entities -- individuals -- each being spatially separated from one another, and each having its internal and external parts also spatially separated from one another. But these material entities can be " injected " (from the Explicate Order) into the Implicate Order. They then are folded back along the space and time dimensions (they are enfolded or implicated). These dimensions now only appear as numerical parameters, and the result is a definite and purely qualitative content which we call a "noëtic entity" residing in the Implicate Order.
The potential ecological niche of the given organic species is also a qualitative content (in the Explicate Order represented by existing individual instances of it). It is a qualitative content because it is definite. And it is definite because it is precisely determined by something which is itself a definite qualitative content, namely the given organic species whose potential ecological niche we are now discussing. And in addition to this it is co-determined by some general features of the environment.
But how is this potential ecological niche so determined (and thus precisely delimited)?
For this we must consider the existing individual characters or features of the species, the potential ecological niche of which we are now considering. If these features change, the species generally is transformed into a new species and is adapted to a new ecological niche. Any given character (for example a feature of the wing-venation, a feature of the antennae, or some feature of the digestive system [of the larva or of the adult] ), can, evolutionarily, change only in certain definite ways or in a certain definite direction. For example, such a change, that is, such a transformation of the present state of a given feature into a new state of it, cannot be such that it is an exact evolutionary reversal to an earlier state of it, that is, a transformation back to a state-it-was-in already earlier in the history of the organism carrying this feature. Also those transformations that are incompatible with the present morphology or physiology of the organism, that is, transformations of which the proper initial state happens to be absent, cannot take place. And, of course, all transformations necessarily resulting in an unstable or unfit organism cannot take place, or, when they do so, their carriers are weeded out by natural selection. Further, the individual features of a given species are connected with each other in such a way that an evolutionary change in just a single feature, while all other features remain what they were, does not generally take place in organic evolution. So in one evolutionary step of the species actually taking place, a whole system of existing features of that species changes in such a way that the change in one given individual feature is coherent with the changes of all other features of that species. All this, consequently, determines how the representatives of the species will look like after one evolutionary step. It has, according to its new, but still only potential, qualitative content, become a (potentially) new species. And this new qualitative content -- still in its state of potential, that is, not actual, qualitative content -- in turn determines (not by causing, but just by qualitatively implying) in what environment the (new) species -- if it represented this new content -- can live, or, more precisely, in what new ecological niche it can live. The new, but still only potential, qualitative content thus connects as it were with elements of the existing overall environment, that is, it demands such elements. And if these demanded environmental elements actually turn out to be present in the existing environment (in the broad sense) we have finally found the sought-for potential ecological niche of our species. And indeed, every individual instance of this new niche represents its definite qualitative content. That content is definite because that content is precisely determined and delimited. It is so delimited by the potential form and structure of the original species (how its representatives would look like after one single [feasible] evolutionary step), which potential form and structure in turn was determined by the actual form and structure as it was present in the representatives of that species. And because it (i.e. the potential ecological niche) is, accordingly, delimited and definite, it is a genuine qualitative content (in the Explicate Order [still] existing in the form of individual instances), and can, and will, as such be injected into the Implicate Order (that is, a copy of it is so injected) where it then exists in the form of a noëtic entity, enfolded along space and time dimensions. The potential form-and-structure ( = not the actual, but potential qualitative content) of the original species -- in fact a (not yet existing) form that has acquired the appropriate adaptations to the potential ecological niche (which does exist) -- will not be injected into the Implicate Order because it is not (yet) an existing unfolded qualitative content (So there is nothing to inject). (Let us make this more clear by expressing it a little differently : The potential ecological niche is not some niche that does not yet exist. It is potential only with respect to the species under discussion, that is, it is not yet the ecological niche of that species. But all the environmental elements together constituting the pattern which we later call "ecological niche" are actually present (in the Explicate Order), therefore it can and will -- as a determined pattern -- be injected into the Implicate Order. On the other hand, the potential form-and-structure of the original species, that is, the new species it can become, does not yet exist (in the Explicate Order), and consequently cannot be injected into the Implicate Order.). So what is injected from the Explicate Order into the Implicate Order is : (1) the qualitative content of the original species, (2) the qualitative content of the actual ecological niche of that species, and (3) the qualitative content of the potential ecological niche of that species.
These injections result in the presence in the Implicate Order of their respective noëtic counterparts. And so we now can point to one of these noëtic entities, namely the one which in the Explicate Order exists as the potential ecological niche of our species, and this noëtic entity can noëtically react with another noëtic entity, namely the injected qualitative content of the original species. The (timeless) product of this reaction is, of course, also a noëtic entity. It is a new qualitative content having resulted from the noëtic integration of the two mentioned noëtic reactants : (1) the qualitative content of the original species and (2) the qualitative content of its potential niche. This new qualitative content, that is, the product of the noëtic reaction, is the qualitative content of the new species that has acquired the appropriate adaptations to the potential ecological niche of the original species. So far this new species still does not exist.
However, if now in the Explicate Order there exists a number of individuals of the original species that have access to at least one of the existing individual instances of the potential ecological niche of that species, the mentioned new qualitative content, that was noëtically generated in the Implicate Order, will project into the Explicate Order, that is, it will unfold itself along space and time dimensions (as they were specified in it in the form of numerical parameters), resulting in individual instances of the new species. In the Explicate Order this 'event' of unfolding (of the noëtic reaction [stages] and of the [final] noëtic product) will be 'observed' as the successful invasion by members of the original species of a new ecological niche by way of developing the appropriate adaptations (resulting in this original species to become a new species). In Part LXIXb of the present Series on Evolution we will further elaborate on these ideas, and add many details.
Let us now apply all this to our flies more closely.
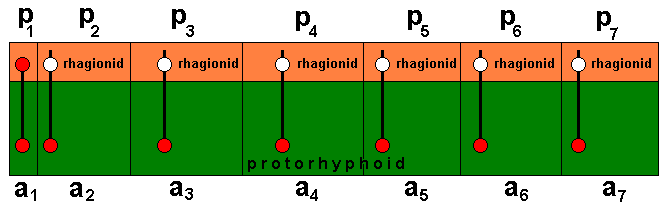
Diagram 1 : Actual (green) and potential (orange) ecological zones and niches of the Protorhyphidae.
a1, a2, a3, a4, a5, a6, a7, (green) --- different actual ecological niches of species of Protorhyphidae. Together they form the actual ecological zone of the family.
p1, p2, p3, p4, p5, p6, p7, (orange) --- different potential ecological niches of these species of Protorhyphidae. Together they form the potential ecological zone of the family. After invasion it becomes the actual ecological zone of the family Rhagionidae. This zone will then consist of actual ecological niches of corresponding species of Rhagionidae. The rhagionid (red) at the left end is supposed to be such that it cannot evolutionarily develop further (in the direction of muscoid flies). The other rhagionids can, and will, evolutionarily develop further : When representatives of them have successfully invaded the corresponding potential ecological niches they turn into empidoids, which is again one step closer to the evolutionary production of muscoid flies (see previous document).
Let us analyse this further in terms of the Implicate / Explicate Orders and noëtic reactions on the basis of considering just one of the potential / actual pairs of ecological niches : a3 harboring, say, the species Archirhyphus asiaticus ROHD. (Family Protorhyphidae [Jurassic] ), and p3, its potential ecological niche :
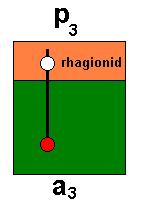
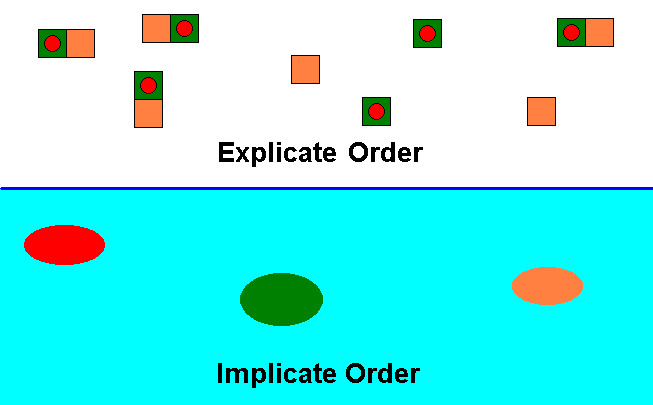
Diagram 2 : The species Archirhyphus asiaticus ROHD. (Family Protorhyphidae [Jurassic] ) (red), its actual ecological niche (green), and its potential ecological niche (orange), as they are present in the Explicate and Implicate Orders of Reality.
In the Explicate Order the qualitative content of the species Archirhyphus asiaticus was represented by a number of individual instances (red), which here were individual flies ( The family Protorhyphidae lived in Jurassic times). Also the species' actual ecological niche (which is the environmental extension of the species' qualitative content) is represented by a number of individual instances (green). And the same applies to the species' potential ecological niche (orange). The diagram expresses the fact that not all individual instances of the species have access (through just one evolutionary step) to one or another individual instance of the species' potential ecological niche.
In the Implicate Order the species' qualitative content, the species' actual ecological niche, and the species' potential ecological niche, are also present, but here only as the noëtic counterparts (of their unfolded, material, versions in the Explicate Order), implying that they are not represented by individual instances. They are present there as immaterial, not-individualized timeless structures.
These two Orders stand to each other in a dynamic relationship : injection and projection of their contents. See next diagram :
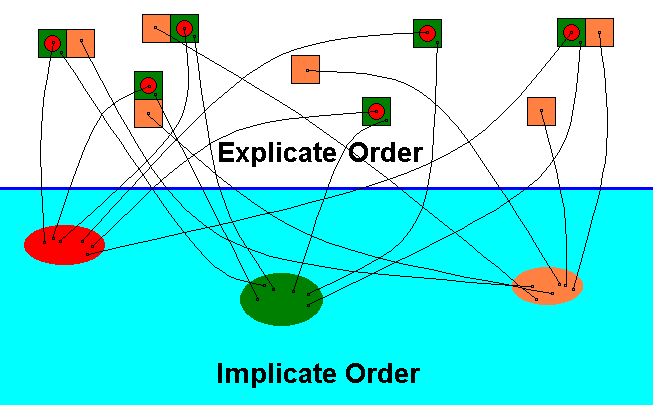
Diagram 3 : The species Archirhyphus asiaticus ROHD. (Family Protorhyphidae [Jurassic] ) (red), its actual ecological niche (green), and its potential ecological niche (orange), as they are present in the Explicate and Implicate Orders of Reality.
Dynamic relationship between the Explicate and Implicate Orders by way of injection (always of copies) of the individual instances of a given existing (in the Explicate Order) qualitative content from the Explicate Order into the Implicate Order, resulting in the single noëtic counterpart of the collective of these instances :
Collective of individual instances of the qualitative content of the species Archirhyphus asiaticus (red) ==> single noëtic structure (red).
Collective of individual instances of the qualitative content of the actual ecological niche of this species (green) ==> single noëtic structure (green).
Collective of individual instances of the qualitative content of the potential ecological niche of this same species (orange) ==> single noëtic structure (orange).
So of every existing (in the Explicate Order) qualitative content there is, as a result of injection (of a copy of it), a noëtic counterpart (immaterially) existing in the Implicate Order.
The single noëtic counterpart of the collective of individual instances of the qualitative content of the species Archirhyphus asiaticus (red) noëtically reacts with the single noëtic counterpart of the collective of individual instances of the qualitative content of the potential ecological niche of this species :
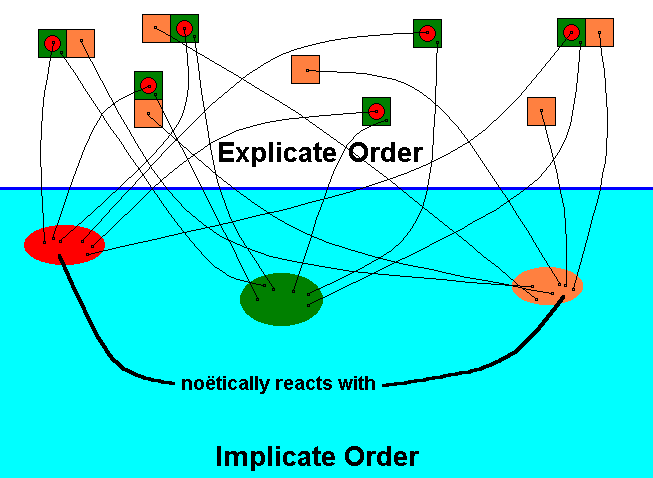
Diagram 4 : The species Archirhyphus asiaticus ROHD. (Family Protorhyphidae [Jurassic] ) (red), its actual ecological niche (green), and its potential ecological niche (orange), as they are present in the Explicate and Implicate Orders of Reality.
Noëtic reaction, taking place in the Implicate Order, between the qualitative content of the species Archirhyphus asiaticus (red) and the qualitative content of its potential ecological niche (orange).
Unlike chemical reactions, in noëtic reactions the reactants are not used up. See next Diagram.
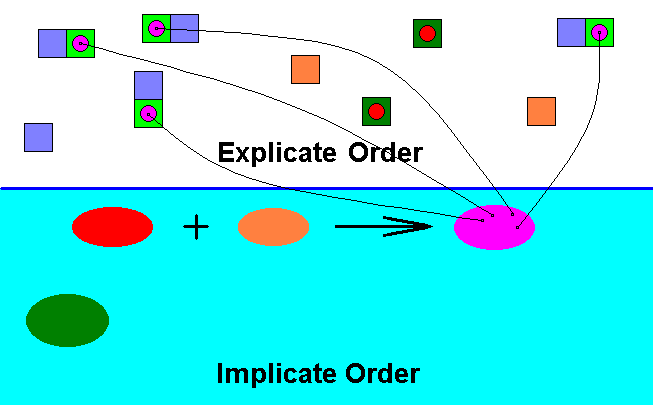
Diagram 5 : The noëtic reaction between the qualitative content of the existing (or having existed) species Archirhyphus asiaticus (red) and the qualitative content of its potential ecological niche (orange) results, as a noëtic reaction product, in the qualitative content of a new species (belonging even to a new family : Rhagionidae) (purple). Where in the Explicate Order individual instances of the species Archirhyphus asiaticus had access to (one or more) individual instances of its potential ecological niche (itself defined -- not caused -- by the not yet [in the Explicate Order] existing qualitative content of the new species) the content of the noëtic reaction product (purple) projects from the Implicate Order into the Explicate Order. What we in fact see there is the successful invasion of members of the species Archirhyphus asiaticus in the potential ecological niche of this species, where "successful" means (1) the evolutionary acquisition of the appropriate adaptations to this ecological niche, and (2) the transformation into a new species.
The former potential ecological niche (orange) of the original species must now be called the actual ecological niche (light green) of the new species. And the qualitative content of this new species defines the next evolutionary step, that is, defines what exactly the states of its features will be after this next evolutionary step, which new (but not yet realized) states in turn define, and delimit, the potential ecological niche (blue) of this new species.
The role played in the evolution of Diptera by their wing-venation, and its description in terms of noëtic dynamics in the Implicate Order.
We have already payed much attention to the question of the role played in the evolution of the order Diptera by the transformation of the wing-venation in its members. To answer this question we had to investigate whether, and if so, in what degree, the wing-venation is a functional character, or just a mere formal character of the insect. When it is functional it embodies a certain flight-regime, and a change of wing-venation means a corresponding change of flight-regime. Assuming that the wing-venation in Diptera is indeed a functional character, functional in the way described, we further had to investigate the relationship of any given flight-regime with the ecology of the species possessing this flight-regime. The most important and determining aspect of the ecology of any given species of Diptera was considered to be its larval development, that is, the habitat, the substrate and feeding habits of the larva. And to "this habitat, etc. of the larva" also belongs (1) the selection -- in the sense of physiological determination -- from the many potential nutritive substances present in its habitat, of precisely those (chemical) substances that are appropriate to, and biochemically compatible with, the overall biochemistry and physiology of the larva of the species, and (2) how precisely the larva makes, biochemically and physiologically, use of these nutritive substances that evolution had chosen to represent its regular food. We preliminarily concluded that the particular flight-regime of a member of any given dipterous species was a natural extension and consequence of the larval ecology of that species. Further we assumed the transformation of larval ecology to be the chief force of evolution. But although there must exist a certain connection between the specifics of larval development and the flight-regime of the adult fly, there does not seem to exist a precise parallel between different ways of larval development and different flight-regimes (and, consequently, different wing-venations) in the corresponding adults. The proper habitat of the larva of course determines the places where the adults can be found (that is, where they will lay their eggs), and the physiognomy (i.e. the micro-landscape) of these places is expected to be more or less reflected in the nature of the flight-regimes, and thus in the wing-venation, of these adults. This connection, however, will be far from specific. Perhaps even it is largely non-existent.
So it is clear that the question of the role played in the evolution of Diptera by the flight-regimes and thus the wing-venation of its members is far from answered, and, perhaps, never will be finally answered. This is even more true with respect to the description of that role in terms of noëtic dynamics in the Implicate Order. We have described the transformation of species of Diptera (leading all the way from Protorhyphidae to Muscidae (s.l.)) in terms of a noëtic reaction of the formal content of the initial species with the formal content of its potential ecological niche. And the sequence of the wing-venations involved in these species transformations showed us that we have to do with a bundle of parallel lines instead of a genealogical tree. This picture of parallel lines is probably a correct description, but a description not specifically accounting for all observed formal relations between wing-venation and (larval) ecology. In addition to the mentioned type of noëtic reaction other types of noëtic events must be assumed to take place in order to account for the full relationship existing between the wing-venation on the one hand, and larval ecology, the nature of the species, and the nature of the higher taxon (especially the family as the most relevant one), etc. on the other.
The nature of these additional noëtic events might be : possible noëtic interactions between the formal contents of two or more different (but closely related) species. We briefly investigated this possibility, but found out that it cannot represent one of the main (noëtic) forces of evolution.
Click H E R E to see the attempt to incorporate the mentioned noëtic interaction into the fabric of evolution. Although that attempt wasn't fruitful, it is nevertheless instructive.
Let us again try to understand the role played by the wing-venation in the evolution of Diptera. For this we must first of all not forget the difference between the Implicate and Explicate Orders of Reality. In the Implicate Order everything is enfolded, that is, folded back along the the space and time dimensions. Entities exist there only in the form of noëtic structures or patterns. These noëtic entities are immaterial and can be compared with mathematical structures. And between these entities a dynamic is going on, that is, a noëtic dynamics comparable with mathematical dynamical systems such as cellular automata or boolean systems (described in First Part of Website [see LINK to Homepage, below] ). Upon projection into the Explicate Order this dynamics is observable as physical, chemical, or biological processes in space and time. And only in the Explicate Order entities of the Implicate Order can be represented by individual instances, and only in this Order we see the essence-accident duality, that is, only here we can encounter the per accidens.
Having said this we can observe the following (four points) about the relationship between the wing-venation and other biological features in Diptera :
To begin with we present the family Blephariceridae (Diptera, Nematocera) as an example :
For the wing-venation in this family click HERE .
Other families turn out to be associated with other wing-venations, that is, other venational groundplans. And again, one particular genus of such a family has the pure groundplan while the other genera have some characters of it in a different state.
Let us give some more examples :
For the wing-venation in the families Tipulidae, Cylindrotomidae, Trichoceridae, Limoniidae and Ptychopteridae ( = Liriopeidae) click HERE ,
For the wing-venation in the family Sciaridae ( = Lycoriidae) click HERE ,
For the wing-venation in the family Scatopsidae, click HERE ,
For the wing-venation in the family Cecidomyiidae ( = Itonididae) click HERE ,
For the wing-venation in the family Heleidae ( = Ceratopogonidae) click HERE ,
For the wing-venation in the family Chironomidae ( = Tendipedidae) click HERE ,
For the wing-venation in the family Simuliidae ( = Melusinidae) click HERE ,
For the wing-venation in the family Psychodidae (s.l.) click HERE ,
For the wing-venation in the family Rhagionidae click HERE ,
For the wing-venation in the family Asilidae click HERE ,
For the wing-venation in the family Nemestrinidae click HERE ,
For the wing-venation in the family Empididae click HERE ,
For the wing-venation in the family Clythiidae ( = Platypezidae) click HERE ,
For the wing-venation in the family Syrphidae click HERE ,
For the wing-venation in the families Muscidae, Phasiidae, Larvaevoridae and Cylindromyiidae click HERE ,
For the wing-venation in the families Drosophilidae click HERE .
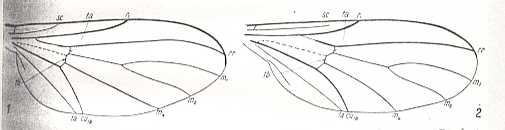
Figure above : Convergence of wing-venation.
1 -- Diadocidia spec. ( Nematocera, Diadocidiidae).
2 -- Empidideicus spec. ( Brachycera-orthorrapha, Bombyliidae).
(After EDWARDS, 1926, indications inserted by HENNIG, 1954)
There can be no doubt that these two species occupy different ecological positions.
On the basis of these and similar observations we can say the following about the biological and evolutionary status of the wing-venation in Diptera :
The wing-venation in Diptera is NOT a mere marking as we see it with respect to colored spots on the wings of many butterflies (excepts patterns connected with mimikry, or with deterring predators [eye-spots] ) and even on the wings of some Diptera (Trypetidae). It has a function. But this function is not confined to merely mechanically supporting the wing-membrane, beause to fulfil this function two, three, or four longitudial veins would be sufficient depending on the absolute size of the wing. But in fact a basic pattern consisting of five vein-systems is persistently present in all (winged) Diptera. And where it is not present it can be easily understood as the result of reductions in this pattern. This persistent basic pattern is :
In the next document we will -- before continuing with the systematic treatment of Diptera -- investigate the non-functional aspects in the family- (or superfamily-) groundplans in Diptera, and their bearing on the theory of evolution as the latter is ultimately in terms of the Implicate Order and noëtic reactions. And then further notes will be added to the biological significance of the wing-venation.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXIV.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII