
Organic Evolution in terms of the Implicate and Explicate Orders.
Part LII
Hymenoptera (wasps, bees, ants) (Sequel)
The evolutionary diversification in the Order Hymenoptera in terms of Strategies (Sequel).
Fifth vespine (moneduloid) phase
The change of provisioning conditions, in which one cell began to be provided with several preys, served as the beginning of yet another deep, and as it were, unexpected change of maternal instinct of wasps. This is the habit of some of them to oviposit in a still empty cell, and only after that to bring in provisions (into that cell). A number of circumstances led to this change of instincts. An indispensable condition for such a change was, first of all, the above mentioned habit of some wasps to attach the egg with its posterior, aboreal, end toward the prey's body. We cannot assume that this habit was reached by wasps that attach their eggs in another way -- with its entire length, or still more differently, with its head-end. In these conditions [i.e. laying the egg in a still empty cell, and having it attached with its head to the cell-wall] their young, helpless, larvae would [after provisions had been brought in] with their head be turned away from the provisions or having placed their head onto a non-nutritive substrate (cell-wall), and would inevitably perish. The second condition [having led to oviposit in a still empty cell], undoubtedly, was the bringing in of several preys [instead of only one] in one cell. This is, as will now be explained, a consequence of this change of instincts. As yet we note, that we do not know of wasps that in preparing only one prey did not oviposit onto it but onto the cell-wall.
It is possible that with the present change of instincts, in certain cases at least, corresponded also the condition of the preys that are being carried into the cell. When the preys are paralyzed, or, even more so, dead, they form no danger in the sense of inflicting mechanical injury to the egg or young larva of the wasp, while this cannot be said of preys having recovered in the cell, and naturally try to escape. In such conditions it would be important to protect the egg and young larva from this danger. In all this the most appropriate thing to do was laying the egg not onto the prey, which might be insufficiently paralyzed, but away from it, on the wall of the cell.
Apparently, in the beginning of all this the work-order remained what it was : The cell was prepared, the first prey was brought in, the egg was laid, and the provisioning continued. But then a spatial separation of two acts took place, acts that before were closely connected with each other (the bringing in of the first prey and oviposition), and as a result the egg began to be laid isolated onto the cell-wall, close to the prey that was brought in before.
To demonstrate such cases in nature is not an easy matter. When we open up a cell, in which the egg turns out to lie away from the first brought-in prey, doubt will arise as to what the wasp had done before : Did it bring in the prey and after that oviposited next to it, or vice versa? Anyway, in certain, apparently anomalous cases we can find the truth. Thus, NIELSEN, 1945, opening up a cell of Bembex rostrata, once found that the egg in it lay freely on the sand next to one of two brought-in flies. Because it is known that this bembex [normally] lays its egg onto the first brought-in prey, it is evident that in the present case the wasp had laid its egg after the bringing in of the first prey, but already not onto it, but next to it, onto the wall of the cell.
Today we do not need to have recourse to "rational coordination" of the acts of the wasp and supposing a rearrangement of acts in time. Here, as in the case of constructing the accommodation before provisioning [hunting for, and catching the prey], or as in the case of bringing in several preys into the cell, we may hold that a certain act -- the hunt, connected with the bringing-in of the first prey (AC), -- was cancelled, and the wasp continued its activity with the next act, that is, with oviposition (D). The whole series of acts then undergoes little change (See for the meaning of the symbols HERE (Part XLIX) ) :
( B + AC + D + (AC)n + E ) ==> ( B + 0 + D + (AC)n + E ) ==> ( B + D + (AC)n + E )
The result thus is : constructing the nest, oviposition, bringing in the first, second, third, ... n-th prey, closure of the nest.
In this way, we should think, the habit of laying the egg in an empty cell originated. As a result of this change, the first prey, that the wasp now brings in after oviposition, corresponds now not anymore to the first prey (AC) but to the second prey in the series of nests of those wasps with which this habit began.
According to the substantial observations of TSUNEKI, 1958, in some colonies [in the sense of aggregations, not in the sense of societies or states] of the Japanese bembex (B. niponica Sm.), mentioned earlier, usually nests are found with several cells in each one of them, and in the individual nests is observed even a simultaneous nursing by the wasp of two larvae. The first prey, to be brought in, does not serve as food for the larva, but is often used only as a postament (base) for the egg of the wasp. Therefore, in line with what was expounded above, it is easy to think that the instinct of bringing in the first prey, not serving already as food for the larva, may be cancelled, and the egg beginning to be laid immediately onto the bottom of the cell.
Such a peculiar situation, in which the egg began to be laid onto a non-nutritive substrate, beit in a specially built accommodation, onto the wall of the cell, is not observed in any earlier mentioned higher wasp of the previous Phases. Among the digger wasps [Sphecoidea] we encounter it for the first time, but also here it is observed as a mere rare phenomenon, and, as far as is known, only in two closely related families -- in the Stizidae and Bembecidae. Apparently, the danger, not so much for the egg of the wasp, laid before provisioning, but for the hatched larva, later led to the fact that the egg was beginning to be laid in a still empty cell with special precautions : deep into the cell, in a vertical position, with the head-end up, etc. Thus, Stizus tridens F. and Bembex mediterranea Hdl (olivacea auct) do not lay their egg simply onto the bottom of the cell, but place it with its head-end up onto a special postament (base) of sand. See next Figure.

Figure 1 : In the cavity of a cell of Stizus tridens F. the egg is visible, placed onto a postament of sand grains. (After MALYSHEV, 1966)
Having laid the egg, these wasps await the hatching of the larva, and then bring in prey, day after day, proportional to the growth of the larva.
Similar, but evidently having still further evolved instincts are known thanks to the data of HUDSON, 1892, and others, with respect to the, related to the membexes, wasp, from South America, Monedula (Stictia) punctata F., and also M. chilensis Esch. (CLAUDE-JOSEPH, 1928). Usually these wasps gather in the form of colonies on bare and hard soil. The egg is laid deep in the burrow, and when the larva emerges the mother starts to bring in provisions for it, whereby after each visit of the burrow she closes its entrance with loose earth. The prey consists of various insects, which are killed by the wasps.
It is possible, as BOUVIER, 1901, indicates, that not all preys are brough in dead, and on closer inspection may turn out to be just paralyzed.
The end stage of development into this direction represent the instincts of the American wasps Microbembex sulfurea Spin., and Microbembex monodonta Say. See next Figure.
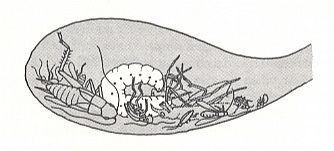
Figure 2 : A larva of Microbembex monodonta in the nest cell, feeding on a variety of dead arthropods (drawn from a photograph). (After EVANS and EBERHARD, 1973)
Also here the egg is firmly placed on the sand deep in the empty cell. In the course of two to three days, provisions are not brought in until the larva emerges. These provisions consist of various insects -- damselflies (Odonata), Hemiptera, Hymenoptera, beetles (Coccinellidae, Chrysomelidae, Carabidae, Scarabaeidae, Gyrinidae), and sometimes also of spiders and even of harvestmen [long-legged spiders]. Such varied prey, for a large part already dead, and even dried-out, the wasp collects while flying low above the soil. Not seldom it also collects parts of insects, especially legs of grasshoppers.
We get the impression that the instinct, having developed into the direction of widening the range of collected preys, has arrived here to a limit [end-stage]. Although a certain ability to paralyze or kill a living prey still is preserved, the necessity for it began to disappear, or has completely vanished. The maternal instinct of the wasp, having it to collect dried-out dead bodies of insects, already not suitable as food for the larva, loses as it were its sense of measure and evokes in life the habit to collect that what is unfit, and which before [this evolutionary phase] was thrown away as being a mere hindrance. From the created dead end a direct way out was not found by the digger wasps, but it was found by, developing in their own way, the pleat-winged wasps [Vespoidea], in which oviposition in an empty cell already began to be a general phenomenon.
On the further evolution of the behavior of digger wasps (sphecoids) the author [MALYSHEV] will dwell more precisely when discussing the question concerning the origin of bees.
The focal moment in oviposition in an empty cell turned out to be the bringing in of the first prey. We saw that the postponement of this moment until the emergence of the larva of the wasp, clearly connected with oviposition in an empty cell, is just a simple adaptation. But placing the first prey before the completely helpless larva, unable to move independently, such that the larva, simply by bending over (perhaps as a result of its own weight) could directly grab the prey, and moreover at the place most suitable for it -- all this demanded on the mother-wasp for very subtle methods, of which it is still difficult to form a picture. See next Figures.

Figure 3 : A young larva of Stizus tridens F., feeding on the first cicadas brought to it. (After MALYSHEV, 1966)

Figure 4 : An almost fully-grown larva of Stizus tridens F. taken out of the cell together with provisions having brought in before : Two jassids, and one fulgorid brought in after the first two preys. (After MALYSHEV, 1966)
A totally other method to evade these same dangers being created when ovipositing in an empty cell, was taken by the solitary pleat-winged wasps, the Eumenidae. For these wasps, see next Figures.

Figure 5 : Female of the solitary pleat-winged wasp Eumenes pedunculatus builds her nest (top image), brings in a caterpillar (center image), puts the prey into the nest (bottom image). (After Königsmann, in Neue Grosse Tier-enzyklopädie, Band 4, Insekten, 1971)
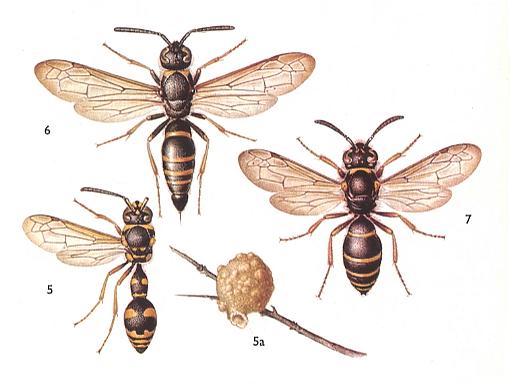
Figure 6 : Eumenidae.
5 : Eumenes pomiformis . 10-16 mm. 5a : nest.
6 : Ancystrocerus antilope . 12-15 mm. [ Ancystrocerus might (also) be a subgenus of the cosmopolitic genus Odynerus including a number of subgenera]
7 : Oplomerus spinipes . 9-12 mm.
(After SEVERA, in ZAHRADNIK, Thieme's insektengids voor West- en Midden-Europa, 1977)
In the Eumenidae the egg is not placed on the bottom of the cell, but suspended from the ceiling, or, more precisely, it is laid at the ceiling of the cell such that the mucus, being secreted at this occasion (secretion from appendant sex glands), is stretched out, and drying up in the air, (this mucus) forms a very thin and elastic thread on which the egg of the wasp is suspended with its head-end down. See next Figure.

Figure 7 : An egg of Odynerus antilope Pz. suspended from the ceiling of an empty cell. (After MALYSHEV, 1966)
Thanks to this, the danger of being pressed down to the bottom of the cell by the weight of one or several preys being piled on top of each other was completely eliminated. On the other hand, the larva, having emerged from the egg suspended from the ceiling, had the full possibility to get into contact with the provisions under it or near to it.
In the present stage of the evolution of the Eumenidae the significance of such a suspension is nevertheless not completely clear. Cases of a successful development of Odynerus callosus Thoms., and O. murarius L. and of others, in [by investigators] deliberately turned-over cells, or from eggs detached [by investigators] from the thread and placed amidst paralyzed prey, do not confirm what was said above. But they [i.e. the mentioned cases of successful development] are also generally little indicative, because the danger of serious squeeze is in all its weight present at the moment in which provisions are piled up in the cell and not after it. In the narrow cells of O. oviventris Wesm. and Eumenes it is indeed observed that suspended eggs turned out to be flattened against the walls of the cell by caterpillars tightly packed into such cells.
The supposition of FERTON, 1901, that the eggs are not suspended because of the danger of being seriously squeezed by the weight of provisions, but because of escaping decay or damage by dampness or parasites, is not convincing because the Odynerus's nest in various conditions while always suspending their eggs, and this does not save them from parasites and lodgers.
On the contrary, Odynerus reniformes Gmel., nesting in the soil, constructs, according to FABRE, peculiar cells, as if divided into two parts : in a rear, more wide one (the "dining room") the egg is suspended from the ceiling, and in the narrow front one (the "storage room") prepared larvae [preys] are piled up. In such conditions the egg is in fact held away from the stock of provisions.
But such a kind of a complicatedly built nest of O. reniformes cannot considered to be primitive among the [species of] Odynerus, usually settling in existing cavities [for example in hollow stalks, and these often offer only little room]. Oviposition in still empty cells is a biological feature of all pleated-winged wasps, Diploptera. The origin of this habit must be sought among the family Eumenidae in the subfamily Eumeninae, especially among the cosmopolitic genus Odynerus Latr., which includes a number of subgenera.
In what way the Odynerus came to oviposit deep in a still empty cell, and what stage of development of their habits went before it? For odynerus wasps, as for Eumenidae, generally, characteristic is preparing in the cells not adult insects but their larvae, chiefly caterpillars of small moths (Tortricidae, Pyralididae, Hesperidae, Psychidae), see next Figure,

Figure 8 : A cell of Odynerus antilope Pz. filled with paralyzed caterpillars of Tortricidae, with the egg of the wasp, suspended from the ceiling. (After MALYSHEV, 1966)
or larvae of small beetles (Curculionidae, Chrysomelidae [See next two Figures], Anobiidae).

Figure 9 : A cell of Odynerus (Symmorphus) murarius L. provisioned with larvae of Chrysomelid beetles. (After MALYSHEV, 1966)

Figure 10 : Part of a cell of Microdynerus biffidus Mor. with an egg, suspended from the ceiling. (After MALYSHEV, 1966)
In rare cases as prey of odynerus are reported the pseudo-caterpillars of saw-flies [Hymenoptera-Symphyta] (in Odynerus capra Sauss., according to ASHMEAD, 1894), and even, according to reports, larvae of caddis-flies (in O. parvulus Lep., according to FERTON. See : BERLAND, 1928, p. 50).
How easy, in similar ecological circumstances, a change of prey or a widening of the range of choice of it could take place in Odynerus is visible in the example of Odynerus regulus Sauss., hunting for prey in the dry flower heads of a composite plant. Depending upon the accidental nature of the hunt, the wasp collects here in some cases small larvae of a beetle (Anobiidae), in other cases larger caterpillars of butterflies (Conchylia), and places them into one and the same cell in a non-regular order.
The Eumenidae not just do not kill their prey, but paralyze them very lightly, such that it often looks quite alive. Such preys may even sometimes molt, changing into pupae or even into adult forms. Here we must reckon also the case of a small percentage of pupae being found among larvae of Curculionidae prepared by Alaster atropos Lep., about which GRANDI, 1935, reported. Also, evidently, we should interpret the finding of pupae of beetles in the cells of Odynerus dubius Sauss.
All these preys are tightly placed in the cells lying in a linear order, and are collected in different numbers depending upon their size : in some cases only from two to six (comparatively large larvae of Chrysomelidae in O. murarius L. (Figure 9)), in other cases up to 30 (small caterpillars of Tortricidae in O. callosus Thoms.). Cases in which only a single prey is found in a cell (in O. murarius L. and in Eumenes) are anomalies, possibly of an atavistic nature.
Only lightly paralyzing the prey in Eumenidae in its densely packed condition in the cells next to the egg suspended from the ceiling, enabled them [the Eumenidae] -- not awaiting the hatching of the larvae --, directly after oviposition to provision the cells one after the other at an increased rate. Therefore, if the upper half of the (reed) stalk, in which the odynerus (f.i. Odynerus callosus Thoms.) earlier built half a dozen cells, is separated, then we may see, below the separated upper part of the stalk, a series of suspended eggs, lightly swinging at the smallest jolt. So fast in these wasps oviposition and provisioning proceed, that when all cells are filled with prey, there is still not a single cell in the nest in which a larva had already emerged.
It is difficult to imagine how, in such narrow accommodations the wasp could preserve the old work-order : oviposition onto the prey after having it brought in an placed inside the narrow nest accomodation [existing cavities in (reed) stalks which are relatively narrow]. Precisely in such spatially narrow conditions, having interfered with the normal depositing of the egg, the act of preparing the first prey could easily be cancelled, and the wasp now beginning with the second act -- deposition of the egg on the wall of the still empty cell [not in the next cell, that is, but simply in the first cell when still empty], attaching it by means of the end of the wasp's abdomen, probably depending simply on the convenience of the pose, on the ceiling of the cell with a thread. The latter could, with respect to the diameter of the nest cavity, in some cases be conspicuously expressed, in other cases hardly be visible -- in the case, for instance, in which the width of the cavity is hardly more than the length of the egg (see Figure 10).
That the events took place precisely in this order, indicate cases described by ROUBAUD, 1916, when the wasp Odynerus bellatulus Sauss. begins provisioning the cell before oviposition, as does sometimes also Rhynchium anceps Grib. [So here still the original work-order]. As a result of difficulties with oviposition onto the prey inside a narrow cell, most probably also takes place the above mentioned unusual method of oviposition by the sphecoid wasp Podium flavipenne Latr. onto the prey brought by it -- a cockroach -- not inside the nest, as do the other wasps, but in front of the entrance to it.
The great majority of Eumenidae proceeds in the described way, that is, after having oviposited in an empty cell, they then, in an accelerated fashion, provision it. Therefore, unexpected is the habit of some of them in, apparently, the same conditions, to await the emergence of the larvae before provisioning them, and moreover [is unexpected] the slowed-down rate of this provisioning from day to day. ROUBAUD, 1916, wanted to see the reason of this slowing-down (at least in certain cases) to be unfavorable conditions of the season. However, not taking into account catastrophic influences, but reckoning with more or less normal conditions, it is not clear why the wasp could not provision before the emergence of the larva, and be in a position to do that [also] after it. Evidently, the reason must be sought for here, as in Bembex, in the inherited slowed-down rate of provisioning. Moreover, the age of the acting individual [wasp] may have a certain significance. Thus, a not yet mature individual, and also an old one, can behave differently than a fully developed, mature, individual. They [i.e. the too young or too old individuals] bring in, for instance, either only a very few, or too much provisions into the cell, but they do not lay eggs in it [judging from their absence], neither before, nor after [provisioning], as was already pointed to by the author (MALYSHEV, 1911, 1936). In other cases (in Odynerus tropicalis Sauss. and Synagris sicheliana Sauss.) slowed-down provisioning is taken as a normal phenomenon in the winter season (central Africa) also by ROUBAUD, 1916, that is, not dependent on drought. And thus we see in the Eumenidae again, as we did in the bembexes and ammophilas, a progressive nursing of the larvae.
One of the most perfect ways of nursing larvae from day to day in all solitary wasps is observed in Odynerus tropicalis Sauss. The prey of this odynerus -- small paralyzed caterpillars (in the beginnining measuring 4-5 mm, and later up to 1 cm) -- is brought in only from the moment of emergence of the larva on, or a little earlier. Usually next to the just emerged larva there is one caterpillar, more rarely two, and never more than three. In line with the growth of the larva provisions are refreshed. It happens that the hungry larva waits until the wasp brings to it a caterpillar. Only upon completion of feeding of the larva the cell is closed. An thus, in closed cells there are no provisions [left].
In connection with all this, the selection of prey in Eumenidae is unexpected -- no flies, as in bembexes, and no adult Holometabola at all, but their larvae. We should not suppose that this selection of prey, typical of early stages of vespine life generally, appeared here for the second time, that is, that the Eumenidae once abandoned hunting for adult insects, and again would have returned all to the original prey of wasps. From here it is clear that the hunt of the eumenids for larvae of beetles, pseudo-caterpillars, and caterpillars shows the roots that go far back into the past, to the lower wasps, and especially to their scolioid ancestors. With the latter [the Scoliidae] they [the Eumenidae] are closely connected by the way of oviposition, and also by the fact that among the scoliids is also observed a hunt for larvae of the curculionid (Rhynchophorus).
With all this, we conclude our exposition of the Fifth Vespine (moneduloid) Phase of hymenopterous evolution.
In the next document we will expound the New Vespine (vespoid) Phase, eventually leading to the social wasps, and concluding the evolution (as it has proceeded up to the Present) of the wasps (We will then continue with the origin of ants, the evolution of bees, and the origin of social bees [which then will conclude our exposition of the evolution of the Order Hymenoptera, as it is conceived by MALYSHEV, 1966. It will be supplemented by addition morphological, palaeontological, and biological data from other sources, and then, at last, our Noëtic Theory of Evolution will be applied to Hymenoptera] ).
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LIII.
Back to Homepage
Back to Contents
Back to Evolutionary Part XIV
Back to Evolutionary Part XV
Back to Evolutionary Part XVI
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XIX
Back to Evolutionary Part XX
Back to Evolutionary Part XXI
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XL
Back to Evolutionary Part XLI
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part L
Back to Evolutionary Part LI