
Further REMARK on noëtics
"although the formally derived patterns in branch 'A' of the noëtic trajectory represent indeed true existential strategies, they cannot yet be projected into the Explicate Order because the ecological situation there is still not yet so structured that it can already receive the projected strategy-pattern. Only if it will finally have become so ecologically structured it can receive that pattern".
The course and the branching of the noëtic trajectory (in the Implicate Order), while it visits one noëtic-stability-field-of-some-noëtic-pattern (in the Implicate Order) after another, that is, when successively one noëtic pattern is formally derived from another, resulting in a branched sequence of derived patterns, is ABSOLUTELY TIMELESS. But upon projection of precisely that c o n t i n u o u s series of derived e x i s t e n t i a l s t r a t e g i e s, not interrupted by derived patterns that do not represent existential strategies, this series will be seen in the Explicate Order as an evolution of strategies -- and therefore of organisms -- IN SPACE AND TIME. That is, the unfolding of the sequence of derived immaterial strategy patterns along the space and time dimensions of the Explicate Order will result in a series of events that we call evolution, that is, concrete evolution, in which ecological situations and changes will play a role. And every strategy involved here is represented by an organic species (or higher-order taxon), itself represented by individual locally existing living organisms. And it is only the Explicate Order in which every thing is spatial, temporal, individual, and local. And indeed the Explicate Order is pure in this respect : There do not exist immaterial, or, as one may say, noëtic, entities anywhere in the Explicate Order. And this means that also 'minds' do not exist in the Explicate Order.
So concrete organic evolution, as we see it in the Explicate Order, is the physical reflection of the corresponding (branched) sequence of formal derivations in the Implicate Order.
( End of remark on noëtics)
In Part XXXI, where we began the exposition of the evolution of the Hymenoptera, we had given an overview of this large insect Order, by presenting a diagram outlining -- as it is proposed by its authors -- the chief evolutionary courses in it, resulting in the evolutionary establishment of the larger hymenopterous groups. To see the place of the 'true wasps' (which are about to be expounded) within the Order (these true wasps being represented by all the aculeates except the bees and the ants), it is perhaps instructive to repeat the mentioned overview :
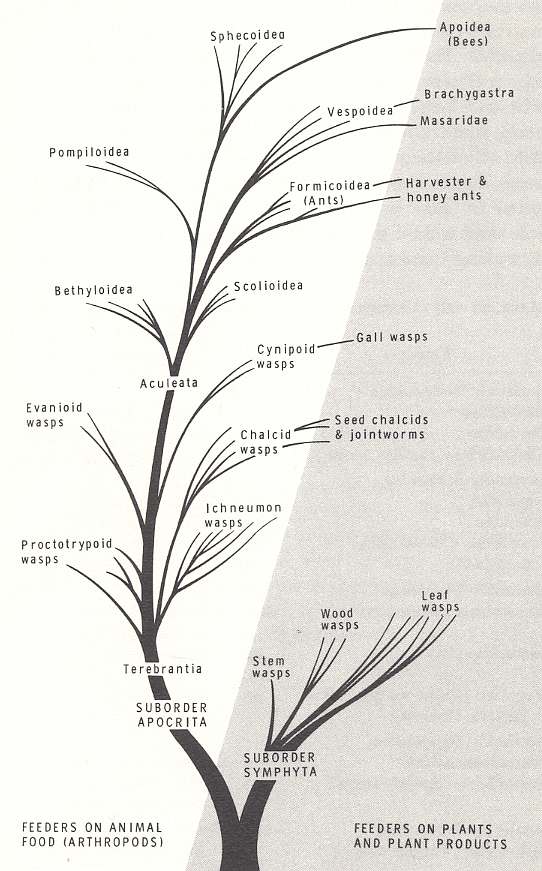
Figure above : Dendrogram showing the probable evolution of the major groups of Hymenoptera. For a magnification click (twice) on the image (and get back by the return button of the browser). (After EVANS and EBERHARD, 1973, The Wasps).
In this dendrogram the Hymenoptera are presented as consisting of two Suborders, the Symphyta and Apocrita. The latter suborder then consists of the Terebrantia and the Aculeata (In our own exposition we consider the Hymenoptera as consisting of three Suborders, the Symphyta, the Terebrantia, and the Aculeata). With respect to the feeding habit of the larvae, the Order originally consisted of phytophags (suborder Symphyta), but soon gave rise to feeders on animal food (arthropods), the terebrants and aculeates. But some representatives of the latter two groups returned to feeding on plants and their products, as is expressed in the diagram.
The question of the origin of the true wasps.
The third suborder of the Hymenoptera -- sting-bearers (Aculeata) -- is a well-defined group. However, when it becomes familiar to us in a more detailed way the distinguishing features between the Terebrantia and the Aculeata almost disappear.
If one takes into account the fact that three superfamilies are contained in the suborder Aculeata, namely the wasps, bees, and ants, among which the wasps undoubtedly represent the basic group, then the question of the origin of the Aculeata in fact boils down to the question of the origin of the wasps, or, more completely expressed, [the question] of which ways and under which circumstances these or those archaic forms -- ancestors of the Hymenoptera with a petiolate abdomen (Apocrita, to which also belong the terebrants) -- could have obtained vespine (= wasp-like) features and evolved in the direction of the Aculeata.
According to HANDLIRSCH, 1908, the groups, usually and rightly so unified as the Sting-bearing Hymenoptera (Aculeata), and which we may, on the basis of the structure of the thorax, divide into Vespiformia and Sphegiformia, undoubtedly separated off from the Terebrantia. Here, the starting point of the Aculeata, according to him, was a form still winged in both sexes, not yet constructing any independent accommodation [for the larva], and, consequently, having been in its larval stage an ectoparasite almost similar to what is still the case today in many Scoliidae and Pompilidae [both true wasps]. Later we will see, that although the vespiforms s.l. (= true wasps) originated together with the terebrants, their lines of evolution [of vespiforms and terbrants] branched off from a wider base [that is, they did not arise from a single point].
As chief distinguishing characters, separating the terebrants from the true wasps, and through them also from the other aculeates, systematists come up with in fact two of them.
The first is : The presence of a two-fold trochanter (at least on the hind legs) in the Terebrantia, and a simple, one-segmented, one in the true wasps.
When at the base of the femur, adjoining the trochanter, a constriction is visible, the trochanter is called a two-fold trochanter. Without this constriction it is called a simple trochanter. [The trochanter is a small segment connecting the femur of the leg with the 'hip' (coxa)]
As to the second distinguishing character : In the Terebrantia the to-be-laid egg is led along the ovipositor all the way down to the precize place of oviposition, whereas in the Aculeata it exits at the very base of the sting, which latter organ is homologous with the ovipositor.
Apart from being only little indicative and often badly distinguishable, the first of these two features is also not always true. Thus, for instance, there are forms (Pelecinidae) which in virtue of a series of characters undoubtedly belong to the Terebrantia, but which nevertheless possess a one-segmented trochanter. Or, the other way around, in the Rhopalosomatidae, belonging to the true wasps, the trochanter is two-segmented. So the first [alleged distinguishing] character cannot, at least in the doubtful cases, held to be decisive.
As regards the second character, connected with egg-laying, in practise it often turnes out to be impossible to determine its presence, especially when one has to do with dead and dry specimens.
According to BOUVIER, 1919, the sting of the wasps does not function in leading the eggs anymore, [this] as a result of displacement of the genital opening toward the outside, and [thus] not located at the base of a tubular organ [anymore] [meant is evidently : not located at the end of the tube, i.e. not at the apical 'beginning' ('base') of it, but at the proximal beginning of it] as in the Terebrantia. This displacement also had to evoke deep-seated changes of the behavior of the insects having undergone this. However, the question, when and under what circumstances, such a displacement took place, is not asked.
The insufficient definiteness of the mentioned distinguishing characters urged MUESEBECK, 1951, and other writers to abandon the division of the Hymenoptera under discussion into Terebrantia and Aculeata, and to confine themselves only to establish one single series of superfamilies, a series that is common to both. However, such an establishment does not contribute to a clarification of the chief evolutionary directions in the Hymenoptera. When working from the evolutionary point of view, it is very important first of all to focus on the vespine features -- i.e. features typical of true wasps -- [already] present in the behavior of these or those terebrants, and with it also to focus on the conditions under which these [vespine] features here [among certain terebrants] appear.
Elements of vespine life in terebrants
Most significant to the transformation of maternal instincts of terebrants into vespine instincts was one particular character, having appeared in [certain] terebrants as it were unexpectedly, and subsequently having been applied most widely. This is the ability to paralyze the prey. The origin of this ability must be located at least at that particular moment when the terebrants began to place their eggs at a prey that lives in the open [that is, unconcealed], although the proper elements to this were possesed by them also already in earlier times : secretion from the appendant sex glands and the stinging device itself, the ovipositor. While the incarcerated concealed prey, almost lacking the ability to move, is itself powerless to struggle successfully with terebrants, this cannot at all be said of free-living preys. It is known, for instance, how a caterpillar usually defends itself against the attack by a terebrant on it : It tries to ward it off by a violent movement of its body, to clasp it by its jaws, to sprinkle it with fluid coming from its mouth, to run away, or even to fall off from the plant. And this activity of the prey is especially dangerous to the terebrant when the egg is not simply inserted under the prey's skin, as in the case of internal parasitism, but must be placed on it such that the prey cannot reach it with its jaws, nor the ectoparasitic larva emerging from the egg. To such terebrants the ability to paralyze the prey, if only partially and transient, apparently turned out to be a necessity from the very beginning. And it is indeed observed in similar circumstances.
As a clear example of this we can give the description of I. SHEVIREV of the hunt of the terebrant Paniscus ocellaris Thoms. (an ichneumonoid) after the caterpillar of the winter night owl Agrotis segetum Schiff. (see first next Figure) :
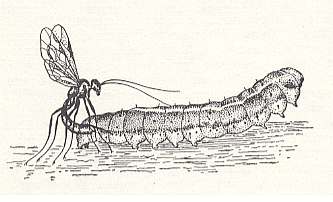
Figure 1 : Female of the terebrant Paniscus ocellaris paralyzes a caterpillar of a night owl. (After SHEVIREV, 1912, in MALYSHEV, 1966)
" Upon detecting the female terebrant, the caterpillar makes some violent movements and speedily runs away, but the female [terebrant] hunts it down, and rising on its feet such that its thorax is directed to the prey, bends toward it the tip of its abdomen with the stretched-out borer and stings it at one of the last segments of the body. Promptly the caterpillar's movements slow down, it stops running and more and more quietly swings its lightly elevated trunk. The female mounts on it and stands on all feet holding its head toward the tail end of the caterpillar, and then attaches the egg behind its second thoracic segment. Now the female withdraws the ovipositor and walks away from the caterpillar, while the small egg, tightly attached by a stalk inserted under the skin, remains on the prey, of which the paralyzed state has already subsided, and which rolls and bends on the ground as if it were attempting to free itself from, or crush in virtue of its weight, the nasty larva" (SHEVIREV, 1912, p. 81).
If the female applies sperm from the seed purse to the egg to be laid, then a female individual will develop from it. In the other case (when sperm is not applied) a male will develop.
This truly remarkable phenomenon of the instinct in terebrants became widely distributed in the higher forms, where the gender of the to-be-born individual is determined, as was wonderfully shown by FABRE, by the size of the chamber assigned to this individual, and by the amount of provisions beforehand prepared for it, and by other circumstances.
Having now seen the appearance of vespine features already in some terebrants, we will now (next document) turn to the evolutionary development (diversification) of the True Wasps, starting with the Semi-vespine (bethyloid) Phase.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XLVII.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV