
Scheme of evolution resulting from the ongoing noëtic transformation of groundplans, and the scheme of evolution as expressed by the genealogical tree obtained from phylogenetic systematics.
Comparing our interpretation of organic evolution as expressed in the Physical-Noëtic Scheme of the origin of, and relations between, organic groups WITH the interpretation of evolution by Phylogenetic Systematics (sensu HENNIG) as expressed by its hierarchic system of monophyletic groups.
Introduction
Already we had seen that according to our interpretation of evolution many organic groups (taxa) can be expected to have evolved polyphyletically, meaning that such a group repeatedly appears from different (but, as we must assume, still more or less related) sources. However, we also found out that this is in fact what we see in the Explicate Order. If we follow things further, that is, follow them all the way down into, and everywhere in, the Implicate Order, then we see the initially (i.e. in the Explicate Order) parallel evolutionary lines often converging onto common lines or points in the Implicate Order. This interesting phenomenon has been touched upon already earlier in the previous Part (previous ducument), namely in the (sub)section Note on the comparison with the hierarchic phylogenetic system of taxa . And the reader might again consult this (sub)section because it can serve as an introduction to the discussion, exposition, and results as they are written down in the present document.
The convergence of evolutionary lines onto common points, and the recognition of monophyletic groups (of organisms) as the only -- within a purely phylogenetic system -- genuine groups, are tenets of Phylogenetic Systematics as it was erected, further developed, and applied, by Willi HENNIG in 1950, 1954, 1966, 1969, and on many other occasions. It is interesting to compare this convergence of evolutionary lines as it is found in phylogenetic systematics (which recognizes only one single Order of Reality, namely the material-physical-biological world of space and time, or in other words, the Explicate Order) with the convergence of evolutionary lines as we see them, not in the Explicate Order, but in the Implicate Order. It is, however, important to realize that Phylogenetic Systematics only recognizes monophyletic groups : If it finds out that some given (and named) group of organisms is in fact not a monophyletic group but merely a paraphyletic or polyphyletic group, then such a group is dropped from the phylogenetic system, whatever its ecological or morphological significance might be -- the phylogenetic system is not a system of "significant" groups but of monophyletic groups. See next Figure.
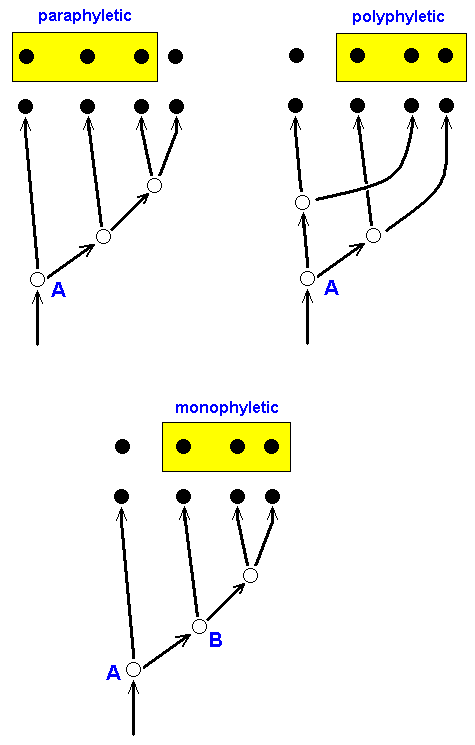
Figure 17a : The phylogenetic status of three groups (indicated by yellow boxes).
The group (marked by yellow box) in the upper left image is paraphyletic : Although all its members descend from a single common ancestor (A), they are not the only ones so descending.
The group (marked by yellow box) in the upper right image is polyphyletic : If we take species A to be the common ancestor (which it is) of the group, the members of this group are not the only descendants of this ancestor. The species A has split up into two species, from one of which a species of our group had descended while from the other all the remaining species of our group had descended. So the direct root of our group is not a single entity : the group has two different direct roots, it is diphyletic (and generally, polyphyletic).
The group (marked by yellow box) in the bottom image is monophyletic : All the members of this group stem from one common ancestor (B) and they are moreover the only descendants of this ancestor. Only such groups are recognized to be genuine groups (taxa) in the phylogenetic system.
Things are different in our interpretation of organic evolution.
Phylogenetic Systematics
The most important tenets of phylogenetic Systematics are already given in Figure 17a, above . It is monophyletic groups that phylogenetic systematics tries to discover among the multitude of organisms. A next step is to find the corresponding sister-groups of these monophyletic groups. Such a sister-group must itself be a monophyletic group, and together they must form a monophyletic group of higher order. Recognition of the fact that a given group is monophyletic is based on so-called synapomorphy of one or more characters (features) present in the members of that group : Every character can, in principle, exist in several states, viz an original or primitive state, one or more intermediate states, and a derived state. Think, for example of the reduction of a particular vein in the wing-venation of a given group of Diptera (let us call it character a ). The process of reduction can show several stages, beginning with the still fully developed vein (a), then a somewhat reduced vein (a'), and, finally, an almost completely (or completely) reduced vein (a''). Such stages, a, a', a'', must themselves of course be homologous features ( In fact, when they really are stages of one transformation process they are then by definition homologous). Together they constitute a transformation series or morphocline. We now call a' apomorphous (= relatively derived) with respect to a, but plesiomorphous (= relatively primitive) with respect to a''. And a'' is apomorphous with respect to a' as well as to a. And, finally, a is plesiomorphous with respect to a' as well as to a''. Now within the confines of a larger group a given subgroup can be expected to be monophyletic if its members share one or more characters that are in an apomorphous state with respect to these characters as they are in other members of the overall group. We then say that the monophyly of the group is probable because of the synapomorphy in this group of the character a :
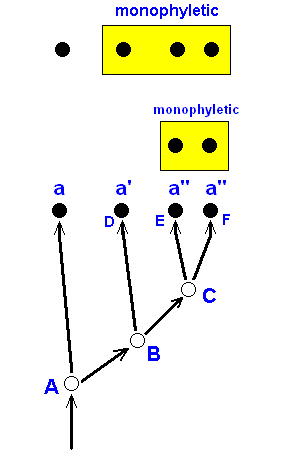
Figure 17b : The phylogenetic status of groups (indicated by yellow boxes), placed within an overall group, is determined (that is, made probable) as being monophyletic based on the common possession (by their members) of relatively apomorphous states (a', a'') of some character a, that is, based on synapomorphy in the group in question. Here it is assumed that state a' originated in species B (from state a) and was then inherited by species D. Then in species C the character became further derived : a''. And this condition was inherited by the species E and F.
So it is assumed that the state a' arose only once (and has been inherited by all the descendants), and that also the state a'' arose only once (and has also been inherited by all the descendants). This is not a general presupposition of the theory of phylogenetic systematics but a particular supposition being made in a particular concrete case of phylogenetic enquiry.
It turns out that synapomorphy when observed in a given series of species does not necessarily imply (morphological) similarity to exist between these species. And because typological systems of organisms, systems that is, that arrange species according to type-categories, are, in fact, entirely based on degrees of similarity, they will often not match with purely phylogenetic systems which are necessarily based on phylogenetic relations between species and monophyletic groups.
Having now given, albeit very succinctly, the essence of phylogenetic systematics, and its difference from typological systematics, especially (the difference from) our version of it, we are now ready to go in some detail in comparing the two. In all this it must be emphasized again that phylogenetic systematics is wholly embedded in only one Order of Reality, the material space-time Order (Explicate Order), and is thus an empirical science, while our version of typological systematics (noëtic theory of evolution) widens (the concept of) Reality such that it considers the existence of yet another Order of Reality, the Implicate or Noëtic Order. This is done in order to account for all the qualitative aspects that are encountered in organic evolution, aspects that cannot be adequately dealt with by empirical science, and thus also not by phylogenetic systematics. But then, of course, such a typological systematics is not a purely empirical science anymore. This fact does not, however, degrade it. Without introducing the Implicate Order, not only phylogenetic systematics, but also typological systematics is for a large part without any firm basis, or at least without any rationale, of a host of observed evolutionary phenomena ('smart' adaptations, the evolution of groundplans, etc.). And phylogenetic systematics, although being fine empirical evolutionary science, is not only very incomplete as a result of not being able to deal with many significant apects of evolution, aspects as significant as, or even more significant than, the mere historical succession of species-cleavages with which it does deal, but is also probably theoretically incorrect because it is based on the assumption that organic evolution is driven entirely by random genetic mutation and natural selection, which is in fact the very reason that it cannot deal with the mentioned phenomena. And as we now know, especially from the work of BEHE ( The Edge of Evolution, The Search for the Limits of Darwinism, 2007 ), but also from that of other biologists, random mutation (combined with natural selection) cannot be the chief factor of organic evolution. The next consideration will show the close connection that exists between (1) the view of phylogenetic systematics that evolution is a temporal succession of species-cleavages, and (2) phylogenetic systematic's dependency on the neo-darwinistic thesis of random genetic mutation in combination with natural selection being the chief driving force of evolution, for explaining the qualitative transformations that have taken place so often in evolution. Because phylogenetic systematics moves only in the plane of the space/time Order of Reality, and because in phylogenetic systematics the begin- and end-phases of at least any major (macroscopical) morphological (and other sorts of) transformation are connected by a series of species-cleavages, the transformation must be rooted in those species-cleavages. So every cleavage must result in the qualitative change of at least one of the daughter species. Species cleavage, however, is as such not something morphological (to emphasize just one of the most important aspects of evolutionary change) or even physiological (it does not all by itself, that is just as species-cleavage produce two different physiologies). So the transformation of the organisms that are involved in such a species-cleavage, or in a series of these, must first of all be genetic. So it must be genetic change that is responsible for the (major, macroscopic, morphological) transformations having taken place during the succession of species-cleavages, especially because incipient species-cleavage is connected with geographical or ecological isolation which finally result in reproductive isolation, the two sister-species going genetically their own way. But apart from the minor genetic changes that each sister-species will undoubtedly undergo during its existence, the only more or less major genetic changes must originate each from a series of consecutive small genetic changes (that together add up to a major change). And because these incremental changes are supposed in the end to create genuine novelty they must be random genetic changes. And so an adaptation to a new environment or the transformation of a body-plan into another are supposed to be the result of longer or shorter series of individual random genetic mutations.
But such a thesis implies great difficulties, because the individual random genetic mutations must appear in the right order, and be such that they or their effects do not in a detrimental way interfere with other genetic processes and their effects (otherwise they will be 'selected away' by natural selection). The chance that such a directed series of random genetic mutations will actually take place, at the right time and in the right place, is enormously small, that is, it is almost zero. So phylogenetic systematics, but at the same time also all sorts of typological systems that move, like phylogenetic systematics, exclusively in the plane of material-biological reality (the Explicate Order), cannot explain, or give at least some rationale for, the mentioned larger evolutionary transformations in organisms. So with respect to phylogenetic systematics there must be something apart from mere species-cleavages that is responsible for these transformations. And our theory of the Explicate / Implicate Orders and the noëtic derivation and generation of organic groundplans in the Implicate Order followed by projection into the Explicate Order is a result of such considerations.
In phylogenetic systematics it is about monophyletic groups, and these are considered to be real entities being each for themselves so and so, that is, entities that are definite with respect to intrinsic content, that is, to what each of them is wholly in virtue of itself. And this 'intrinsic whatness' of a monophyletic group is based on the fact that all its members, and only its members, have descended from a single species, the stem-species of that group. It is thereby assumed that all the members of such a group have inherited important parts of the genome of the stem-species (= the complete set of this species' genes). And apart from recombination of genes (in sexual reproduction) and random mutations there is no further influx of new genes. So the intrinsic whatness of the monophyletic group must, for a considerable part at least, be rooted in the genome of the stem-species. All this according to, or implied by, neo-darwinism. But things are not so simple : According to phylogenetic systematics the Birds and the Crocodiles are sister-groups, which means that they have descended from a single stem-species. So this one stem-species can, by splitting into two, and by subsequent strong development starting from one of the daughter-species (the evolutionary line starting from the other remaining relatively unchanged), give rise to two totally different body-groundplans, the crocodiles and the birds (of which the former represent the more conservative branch). Many other examples of such a phenomenon can be presented. So it seems that the intrinsic whatness of a monophyletic group (such as the Birds) is not completely rooted in the stem-species of the group Crocodiles + Birds. And that it is not completely so rooted is not solely the result of subsequent transformation of the genome of the stem-species as this species is being split up again and again (because that repeatedly splitting up has happened in both lineages (birds and crocodiles)), but surely is also the result of the fact that not every feature of an organism or of a group of organisms is determined by genes (as is demonstrated by Bonellia and by the angler fish). Nevertheless, a lot of them are apparently so determined. Therefore a monophyletic group can -- within the theory of phylogenetic systematics -- be considered to be an entity that is intrinsically determined to be what it is. But we should not absolutize this. We'd better say that such a group is partly intrinsically so determined, meaning that a given monophyletic group, such as the recent Birds, might owe its whatness largly to the genome of its latest (i.e. most recent) stem-species (already far away from the earliest stem-species of the Birds), while the whatness of the monophyletic group consisting of the crocodiles PLUS birds is not so qualitatively uniform that we can consider it to be largely rooted in the genome of the stem-species of this group. We should realize that the only dimension of the phylogenetic system is time : it gives the temporal order of species-cleavages, and nothing more. And this temporal order exclusively belongs to the Explicate Order, while the qualitative dimension (which in fact comprises many dimensions) belongs to the Implicate Order. And time is not explicitly present in that Order.
(1) Evolution as a result of the continuous noëtic generation, transformation, and derivation of organic groundplans, and (2) evolution based on random genetic mutations and natural selection of mutants, resulting in an ongoing process of simultaneous and successive species cleavages, in turn resulting in monophyletic groups, compared.
Before we finally will proceed with this comparison, forming the intended content of this document, we must still consider a few more things first :
In Figure 17 in the previous document we have shown that the stability field of an order-groundplan is partitioned into a number of stability sub-fields, namely those of family-groundplans and those of elementary noëtic reactants.
It is perhaps useful (meaning theoretically plausible) to assume the same relationship of subordination (fields-subfields) with respect to genus-groundplans : the stability field of a family-groundplan of a given family is partitioned into a the stability fields of the groundplans of the genera belonging to that family and those of elementary reactants, that can, upon noëtic reaction, generate the (noëtic content of the) groundplans of these genera. And in the same way the stability fields of two or more genus-groundplans can either be immediately adjacent to each other, or only mediately so, in which (latter) case they are separated by two zones of elementary noëtic reactants. In the first case the groundplans are very much alike each other because the one can immediately be derived from the other. In the second case the groundplans differ substantially, they cannot simply be derived from each other. If, in this case, the one groundplan is formed from the other, there are always (qualitatively) new elementary reactants involved.
Let us compare the two cases :
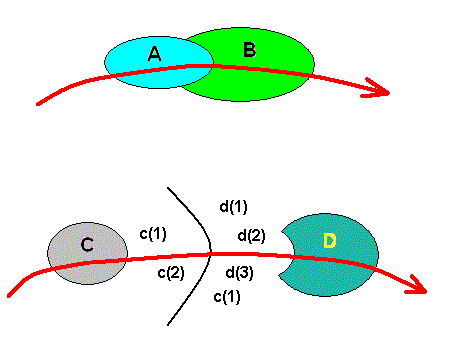
Figure 18 : Two different cases of the successive noëtic formation of two genus-groundplans : ==> A ==> B, and ==> C ==> D.
The genus stability fields (blue, green, brown, dark green) reside within the overall stability field of the groundplan of the family to which these genera belong.
Top image : As the noëtic trajectory (red curve) enters the stability field of the genus-groundplan A, the noëtic content of this groundplan is noëtically stable. So the noëtic content of this groundplan exists there, while that of groundplan B does. Upon leaving this stability field the noëtic trajectory directly enters that of genus-groundplan B, meaning that the noëtic content of genus-groundplan B is stable and thus noëtically exists there, while that of groundplan A does not. This genus-groundplan B is, accordingly, directly noëtically derived from the genus-groundplan A. They are therefore closely noëtically related, that is, they are structurally similar.
After the noëtic trajectory having left the stability field of genus-groundplan B the noëtic content of this groundplan is unstable and does not noëtically exist there.
Bottom image : As the noëtic trajectory (red curve) enters the stability field of the genus-groundplan C, the noëtic content of this groundplan is noëtically stable. Upon leaving this stability field it enters that of the elementary reactants c(1) and c(2). Here these reactants are stable and thus exist. The genus-groundplan C cannot exist here. It has been disintegrated according to the noëtic reaction C ==> c(1) + c(2). When the noëtic trajectory leaves this reactant stability field it enters the stability field of the reactants c(1), d(1), d(2), and d(3). They are stable here and are the result of the noëtic reaction D ==> c(1) + d(1) + d(2) + d(3), that is, the disintegration of genus-groundplan D. When the noëtic trajectory enters the stability field of the just mentioned groundplan, the reactants are unstable and they generate genus-groundplan D according to te noëtic reaction c(1) + d(1) + d(2) + d(3) ==> D. Finally, the noëtic trajectory having left the stability field of genus-groundplan D, this groundplan is no longer stable and cannot exist there. And indeed, because the stability fields of C and D are not adjacent to each other but are separated by the respective stability fields of the mentioned elementary reactants, the genera, respectively corresponding to these genus-groundplans are not closely noëtically related to each other, that is, they structurally stand farther apart from each other than do the genera A and B of the top image.
If we say that the noëtic content of the family-groundplan reacts with the noëtic content of some potential ecological niche, it is in fact, as we now know, a noëtic super-content of the family-groundplan, that is, the noëtic content of the groundplan of one of the family's genera that so reacts. This genus-groundplan contains the family-groundplan, as do the groundplans of all the family's genera. And, as we know, when the noëtic content of the groundplan of this genus reacts with the noëtic content of some potential ecological niche, and (when) the reaction-product has been subsequently projected into the Explicate Order, we there see, now extended along the time dimension, a species adapting itself to some ecological niche (that is, individual instances of this species adapting themselves to this niche).
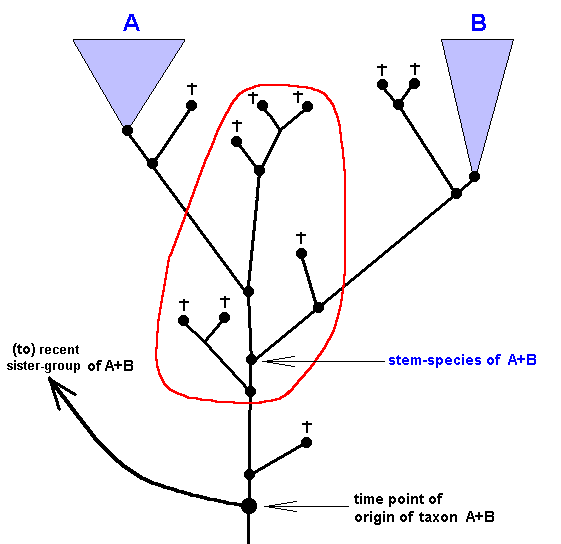
We are now ready to compare the general form of the biological system (the 'genealogic tree') as it results from our ontological conception of evolution (two different ranges of Reality) with that of the hierarchical biological system (genealogic tree) as it results from the presuppositions and methods of (as now standard) phylogenetic systematics (monophyletic theory) as developed by HENNIG (1950, 1966).
To begin with the most familiar of the two, the next Figure presents the typical form of the genealogical tree of organisms resulting from the presuppositions and methods of standard phylogenetic systematics.
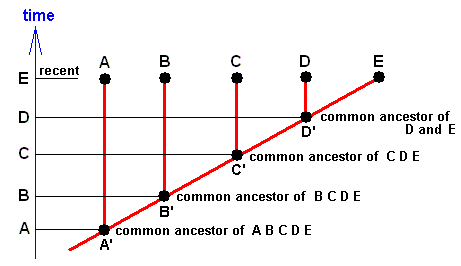
Figure 19 : Example of the typical form of a genealogical tree of a group of organisms in standard phylogenetic systematics.
In the present case we here think of a group of five families, A, B, C, D, and E, together constituting a monophyletic group. The ancestral family of this whole group is the family A' which is supposed to be still -- only having undergone minor changes -- present. That is to say, the same family-groundplan still exists. A' means merely the fossil representative of A. The common ancestral family of the family group DE is the family D' (= fossil representative of D) also still existing in the present. The same applies to the family groups CDE and BCDE.
Along the time dimension (vertical in the diagram) we see the families A, B, C, D, E (and thus their groundplans) successively appear.
Instead of families, we can as well let the symbols A (=A'), B (=B'), C (=C'), D (=D'), and E stand for genera or other taxonomic categories.
A', B', C', and D' can also be interpreted as representing stem species of the respective families. The forks in this diagram (as traced out by the red line) then indicate species cleavages. And after each having undergone many subsequent cleavages (not shown) the families in their recent form appear. Indeed, phylogenetic systematics holds that every family (and every taxon whatsoever) originated from a single ancestral species.
Now, in accordance with phylogenetic systematics the family D is more closely related to the family E than it is to the family C because the common ancestor of D and E, which is D', is more recent than the common ancestor of D and C, which is C'. In the same way C is more closely related to D than to B because the common ancestor of C and D is more recent than that of C and B, which is B'. And the same applies to BC and AB. In short : In standard phylogenetic systematics "phylogenetic relationship" is determined by "recency of common ancestry".
The next Figure tries to 'translate' exactly the same event as was meant to express phylogenetically by the genealogic tree in the previous Figure into the language of our own developed view (involving the Implicate and Explicate Orders) of how families (and other taxa) originate. In this view they are not supposed to have evolved from each other IN THE EXPLICATE ORDER, but did so in the Implicate Order. As has been said, the Implicate Order is an immaterial ('noëtic') dynamical system or multitude of such systems, having trajectories (consisting of, or connecting, dynamic states or stages) that run through the various noëtic stability fields and in this way activate organic groundplans or other noëtic patterns to project themselves from the Implicate to the Explicate Order where they unfold along space and time dimensions resulting in material things or processes ( When we speak here of "dynamical systems" and "dynamical states" as constituting the Implicate Order, these dynamical systems and states are no more than noëtic analogues of real or even purely mathematical systems and states).
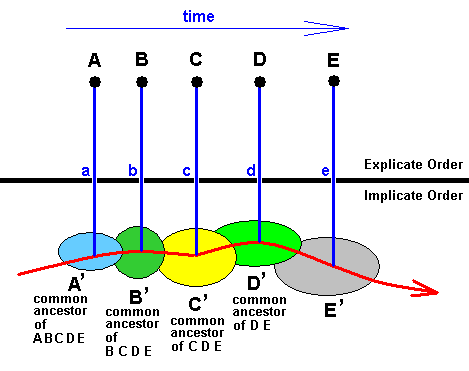
Figure 20 : Translation of the genealogic tree of phylogenetic systematics as depicted in Figure 19 resulting in : an example of the typical form of a 'genealogical tree' of a group of organisms as obtained from considering the interactions between the supposed Implicate and Explicate Orders. This tree typically consists of a noëtic trajectory (red line) in the Implicate Order and successive projections (vertical blue lines) of noëtic groundplan patterns (colored) into the Explicate Order (= unfoldment of these patterns along time and space dimensions), resulting in the successive appearance of groups of concrete organisms.
The colored areas stand for the noëtic stability fields of five organic groundplans (here we take them as family-groundplans). In the blue area (left) the noëtic content of family-groundplan A is stable and consequently exists there. It is indicated as A'. This content A' noetically reacts with the content of some potential ecological niche and the reaction-product then is projected into the Explicate Order appearing there as a family of concrete organisms indicated by A. The same applies to the families B, C, D, and E. The time direction in this diagram is horizontally to the right.
The points a, b, c, d, and e indicate the successive appearance in time of the families A, B, C, D, and E. Also in Figure 19 these families appear in this temporal order. But in contrast to the five vertical lines in the tree of Figure 19 which run parallel to the time direction and are lines of genetic descent, the five vertical lines in the present Figure (Figure 20 ) are lines of projection. They do not run along the time dimension. Only the sequence of points (a, b, c, d, e) at which the noëtic contents were first projected ("first" as seen only in the Explicate Order) lies along the time dimension (which here is horizontally to the right).
In the present Figure we can consider A' to be the analogue of the ancestral form A' in Figure 19 , and in the same way B' in the present Figure to be the analogue of the ancestral form B' in Figure 19. The same applies to C' and D'. But it is clear that the phylogenetic relations as expressed by the cladogram of Figure 19 , cannot as such be found in the present Figure (Figure 20 ). Only the noëtic trajectory in that Figure (red line) determines some kind of kinship relations between the five families.
Having established this we can say the following, having before us Figure 20 :
Family D is more closely related to family E than it is to family C, because the common ancestor of D and E, which is D', is more recent than the common ancestor of D and C, which is C' (C' lies further back in time as the latter's direction is set here). And this corresponds exactly to what had been established in Figure 19 with respect to the families D and E.
Further, C is more closely related to D than it is to B, because the common ancestor of C and D, which is C', is more recent than the common ancestor of C and B, which is B'. Also this corresponds exactly to what had been established in Figure 19 with respect to the families C and D. And the same applies to the grouping BC.
So now we have established the conceptual translation of the events as expressed in Figure 19 , that is, expressed according to phylogenetic systematics, into our own terms, that is, into the terms of the Implicate and Explicate Orders, the noëtic trajectory 'running' through groundplan stability fields, noëtic derivations (here one-reactant noëtic reactions), and projections.
In addition to the type of phylogenetic cladogram as was depicted in Figure 19 there can be others, that is, (there can be) phylogenetic cladograms, although still hierarchic, nevertheless having a different structure :
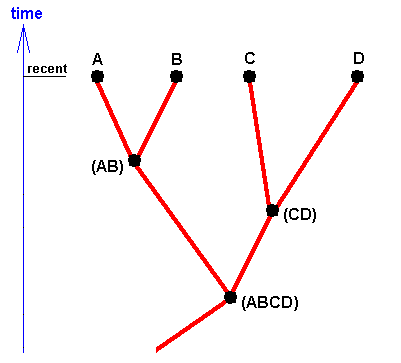
Figure 21 : Phylogenetic cladogram expressing the fact that the recent taxa A, B, C, and D have phylogenetically evolved as a result of repeated evolutionary splitting up of earlier groups into separate taxa.
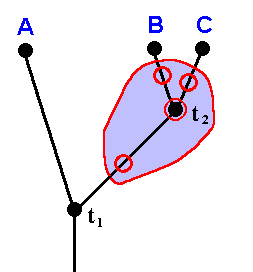
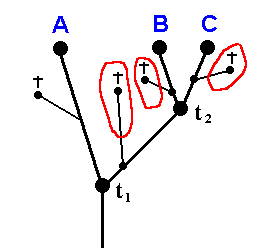
According to phylogenetic systematics, in the geological past there existed a species (ABCD) (that is, a single reproductive community of individuals, or a set of such communities in which the individuals of each community are supposed physiologically and genetically to be able to interbreed with those of all the other communities of that set) that came to be split up in two species (AB) and (CD) (that is, in two reproductive communities, or two sets of such communities, such that the individuals of one such community or of one set of them are physiologically and genetically unable to interbreed with those of the other community or set of communities).
Then, the species (AB) in turn splits up, resulting in the two species A and B. And also the species (CD) splits up, resulting in the two species C and D.
The recent taxa (A + B) and (C + D) are called sister-groups ( We can also call (AB) (A+B) and (CD)(C+D) sister-groups). The species (ABCD) is the stem-species of the two sister-groups taken together.
The groups (ABCD), (AB), (BC), A, B, C, and D were here considered to be species and thus consist of individuals (only). But we can also interpret these groups as being higher taxa, that is, their being such that they consist of species rather than of mere individuals, or of genera. In the latter case all the genera of such a group together form a family, and then we would have the families (ABCD), (AB), (BC), A, B, C, and D. The cladogram then does not depict the details, that is to say, it does not then depict explicitly all the species cleavages involved. However, within pure phylogenetic systematics such an interpretation is not possible. That is to say, in phylogenetic systematics we must interpret all the junctions and all the end-points of the genealogic tree (cladogram) as species. The structure of the tree is then founded on species cleavages. Every two species resulting from such a cleavage are by definition reproductively isolated, and this not only because their respective individuals are spatially far away from each other, but because they are incompatible physiologically and genetically. So the tree is historically constructed by simultaneous and successive cleavages of species. And if we, in this genealogical tree, want to replace the species at the junctions by higher taxa, say by genera or by families, then there is no way to have these taxa split up as we go up in the tree from bottom to top. This is so because we cannot genetically split up a family or genus into two reproductively isolated (sub)families or (sub)genera because any family or genus (and whatever higher taxon) itself already consists of different species which are by definition reproductively isolated from each other. So the only way to let families or genera split up at the junctions of the tree is to let them grow apart, not reproductively (which they and their different species already are), but morphologically (or holomorphologically which includes physiological and behavioral properties). But if we do that, then our system, so obtained, has become typological. It is not genuinely phylogenetic anymore. So if we want to stay within genuinely phylogenetic systematics (sensu HENNIG), then we must hold that all the junctions of the genealogic tree, and all its true end-points, represent species, and only species. And this also means that as long we go with phylogenetic systematics we adhere to its definition of monophyly, and admit that there indeed are large animal groups, such as Diptera, Coleoptera, and bone-fish, that have evolved from a single stem-species. And it is only when we translate the findings of phylogenetic systematics -- its genealogic tree (expressing the phylogenetic relationships of one group or another) -- into the shape and structure of the noëtic trajectory (in the Implicate Order) together with its associated lines of projection,- that we discover the true meaning of the concept of "monophyly".
But, returning to our original point, however we interpret the groups at the junctions of the genealogic tree (as species [groups of individuals] or as higher taxa [groups of species] ), the structure of the present cladogram (Figure 21) is clear, and it is evident that it structurally differs from that of Figure 19 .
If we, for the moment, assume that phylogenies such as depicted in the above Figure (Figure 21) are really possible (which is a sound assumption), then how can we translate the phylogenetic tree, expressing this type of phylogeny, into the language of our theory? The next Figure shows this.
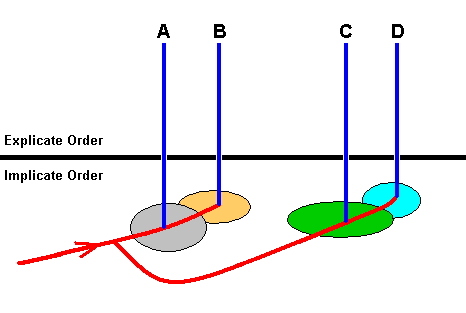
Figure 22 : Translation of the phylogenetic tree of Figure 21 into that ('tree') resulting from our theory of the Implicate and Explicate Orders.
Let us interpret the groups A, B, C, and D as families.
When the noëtic trajectory has entered the noëtic stability field of the groundplan of family A, it is stable and will, after having noëtically reacted with the noëtic content of one of the potential ecological niches of this groundplan, be projected into the Explicate Order (in fact there is a bundle of such projections each involving another potential ecological niche). So in the Explicate Order we see the family A appear, the species of which occupying several ecological niches. And when, after this, the trajectory has entered the stability field of the groundplan of the family B, this groundplan will be projected also when it has reacted with the noëtic contents of the potential ecological niches of this groundplan. And so the family B appears in the Explicate Order having its species dispersed over some ecological range.
But for the noëtic trajectory there is still an additional course to follow that leads, via some intermediate noëtic space, to the stability field of the groundplan of family C. Having entered this field, groundplan C will be stable and will be projected into the Explicate Order in the same way as described for the families A and B. And while progressing still further the noëtic trajectory enters the stability field of the groundplan of family D upon which (entering) the content of that groundplan will be projected, giving rise, in the Explicate Order, of family D.
In terms of dynamical systems there is something odd in all this, consisting of the apparent fact that our noëtic tractory reaches a point, or an area, representing a system-state we might say, where there are two possible next states, namely A and C. This means that the dynamical system (a trajectory of which we are now considering) is -- at least locally -- non-deterministic. Standard physical or mathematical dynamical systems are not like that. And this fact, consequently, demonstrates that the dynamics in the Implicate Order can only be seen as an analogue of 'real' physical or mathematical dynamical systems.
Another possibility is, of course, that a phylogeny as expressed in Figure 21 is actually not possible, which seems to me not very plausible.
Further we can say that -- while still discussing Figure 22 -- family C is more closely related to family D than it is to family B, because there is no connection (through the noëtic trajectory) from B to C.
At the same time we can see that the genealogic tree of Figure 21 (and with it, any genealogic tree of phylogenetic systematics), provided it be interpreted in the way as is done in Figure 22 , can have its junctions representing higher taxa (such as genera or families), or, in any case, not species. Maybe we can better express this as follows : If we interpret (reformulate) a genealogic tree, as obtained by phylogenetic systematics, in terms of Implicate/Explicate Orders, noëtic trajectory, and of (generic-, family-, or order-) groundplans, then we can let higher taxa play the chief role in evolution rather than species ( The latter are only differentiated when at last the noëtic content of a [genus-] groundplan has reacted with that of one of the groundplan's potential ecological niches and when the reaction-product has finally been projected into the Explicate Order, where we then see different species of the family adapting to different ecological niches).
Problems in phylogenetic systematics as compared with that based on the Implicate / Explicate Orders and noëtic reactions.
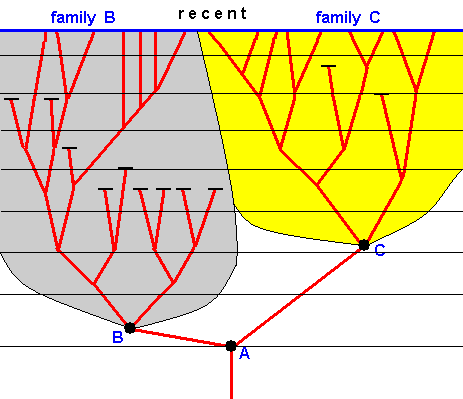
Figure 23 : Genealogic tree according to the principles of phylogenetic systematics.
If we ascend from bottom to top we ascent from the geological past to the present. Each horizontal section through the tree represents some geological period. All the junctions and all the end-points of the tree represent species, and all the junctions at the same time represent species cleavages. Species A, B, and C, highlighted. The (red) line segment connecting two immediately successive junctions represents the existence of a species during which it doesn't change. Also the terminal (red) line segments represent such an existence.
Some species have come to be extinct as is indicated in the Figure.
Species A splits into species B and C. All the descendants of species B together form the family B, and all descendants of species C together form the family C.
In all 'horizontal' sections through the tree lying above the first appearance of species B, the family B is represented by more than one species. That is, in all geological periods since the origin of species B the family B is present, not as a single species but as a complex of several species.
In all 'horizontal' sections through the tree lying above the first appearance of species C, the family C is represented by more than one species. That is, in all geological periods since the origin of species C the family C is present, not as a single species but as a complex of several species.
This Figure will now clarify the following considerations :
In phylogenetic systematics (sensu HENNIG) the genealogic tree of organisms is constituted of a pattern of simultaneous or successive species cleavages. Moreover, we can discern 'horizontal' layers or sections in such a tree each representing a certain period in geological history. Lower layers represent older periods, higher layers younger. Every junction in the tree represents a species, and also the tree's end-points represent species (provided the tree is drawn completely, that is, if the upper layer of the drawn tree represents recent times). These latter species, that is, the top layer of species, are distributed over several groups, the higher taxa, as dictated by the structure of the whole tree lying below this top layer. Said differently, the structure of the whole tree lying under its end-points dictates that the species at the end-points form, among themselves, distinct larger or smaller groups, which are the higher taxa. But also in every layer below the top-layer the species form groups following from the structure of the whole tree below such a layer. And every such group is monophyletic, that is, all the species of this group, and only these species, have historically descended from a single stem-species, the ancestral species of the group, (the stem-species) itself also belonging to this group.
In order to dig deeper into our interpretation of evolution and comparing this interpretation with that of pure phylogenetic systematics, let us add more details into our established Figure 22, above which presented our interpretation of the phylogenetic tree of Figure 21 . In Figure 22 the four depicted noëtic stability fields were considered to be those of the groundplans of the families A, B, C, and D.
But each family-groundplan stability field is partitioned into subordinated stability fields of the groundplans of the family's genera and the stability fields of the elementary noëtic reactants that after having reacted with each other generate the generic groundplans (genus-groundplans) :
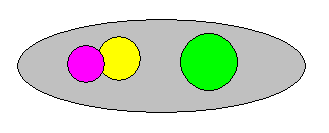
Figure 24 : Stability field of a family-groundplan containing the stability fields of the groundplans of the family's three genera.
Depending on the course of the noëtic trajectory, one genus-groundplan can be directly derived from another (one-reactant noëtic reaction) and another can become the product of a noëtic reaction between elementary reactants :
Figure 25 : Stability field of a family groundplan containing the stability fields of the groundplans of the family's three genera.
When the noëtic trajectory (red curve) enters the stability field of the family-groundplan, the latter will be stable. And also the elementary noëtic reactants that can produce the genus-groundplan, that is, either the groundplan of the one genus or that of another of the family, are stable. When, upon proceeding, the trajectory enters the stability field (purple) of a genus, those elementary reactants that can produce this particular groundplan become unstable and noëtically react with each other creating the groundplan of that genus. When, upon proceeding, the noëtic trajectory directly enters the stability field of the groundplan of a second genus (yellow), that groundplan will be noëtically derived from the previous groundplan. And, when, the noëtic trajectory leaves the stability field of this genus-groundplan, this groundplan will disintegrate into individual reactants. And when the trajectory comes near the stability field of the groundplan of the third genus of the family, this genus' elementary reactants become stable. And when, finally, the trajectory actually enters the stability field of the groundplan of this genus (green), this groundplan will be stable, and comes to exist as a result of the reaction between the mentioned noëtic reactants.
The (noëtic content of the) groundplan of each genus, will, when it is stable, noëtically react with several of the family's groundplan potential ecological niches (that is, react with their noëtic content), more precisely, (react) with several of the potential ecological niches of the genus-groundplan, before this genus-groundplan (including the family-groundplan) will be projected -- in the form of a bundle of species -- into the Explicate Order :
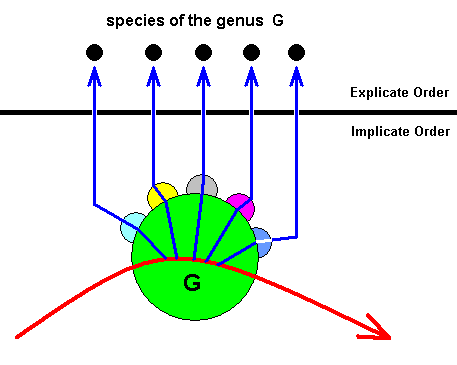
Figure 26 : After the noëtic trajectory (red curve) having entered the stability field of the groundplan of, say, genus G :
The noëtic content of the groundplan of genus G has (noëtically) reacted (sucessively) with the noëtic content of five of its potential ecological niches. The five reaction-products are then projected into the Explicate Order where they appear as individual instantiations of five species of the genus G (living) in their proper ecological niche, where each species has become specifically adapted to its niche. Here we see the monophyly of a genus as this is expressed in the Implicate Order. It is not expressed in the Explicate Order.
So, to the type of situation as depicted in Figure 22 we have added details, namely the partition of the family-groundplan stability field into subordinated genus-groundplan stability fields and the projection of these groundplans into the Explicate Order in the form of a bundle of species. All this is meant to have general significance, that is, it not only is relevant to our example, but also definitely points beyond it.
The relative and absolute (geological) age of phylogenetic taxa (monophyletic groups) and the absolute order of rank of these taxa.
We are now well under way in expounding in what way monophyletic groups and phylogenetic relations in phylogenetic systematics are to be understood and recognized, and in what way they should be translated into the events as they take place in the Implicate Order. It is important to realize that this 'translation' does not mean that the noëtic events in the Implicate Order, insofar as they represent the noëtic derivation of one organic groundplan from another or the generation of new groundplans from noëtic reactants, are seen in the Explicate Order in the form of a sequence of species-cleavages. The splitting up of a species into (two) daughter species is never actually observed in the Explicate Order. It is an assumption, and especially an assumption of phylogenetic systematics, and it results in the genealogic tree of phylogenetic systematics. And as has been said repeatedly, we, here on this website, cannot agree with these assumptions and what results from them. We can only agree with them after having them re-interpreted in terms of our theory of the Implicate and Explicate Orders and noëtic reactions. And this means that according to us all phylogenetic events, as described by phylogenetic systematics, take place, not in the Explicate but in the Implicate Order. In the Explicate Order all we have (and all there is) are parallel lines of temporally successive or simultaneous appearances of orders, families, genera, and species, not, however, as such, but in the form of individual instantiations, that is, in the form of concrete material individual organisms.
Having understood this, we can now proceed to discuss the historical development of taxa as it takes place according to phylogenetic systematics and as it will lead us to an understanding and an appreciation of the concept of stem-group.
Because phylogenetic kinship is equivalent to recency of common ancestry, it is naturally connected with the (geological) age of taxa, and this age determines the absolute rank of them in the hierarchic system of phylogenetic systematics : genus, family, order, class, etc. If groups of organisms have, on the basis of unequivocal and objective considerations, the same taxonomic order of rank in a system of classification, then this expresses a certain degree of comparability of these groups. And that is important because in paleo-geographical and paleo-ecological reconstructions we must have at our disposal systematic units that are historically comparable. Consequently, these units must themselves be historical independent entities, historically 'operational' units, that is, monophyletic groups. And in such groups we can, as to their geological age, discern certain things. Let us do this with the help of a simple example :
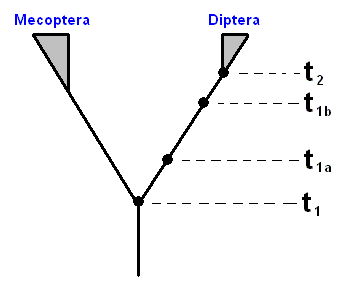
Figure 27 : Simplified phylogenetic scheme of the supposed monophyletic group (Mecoptera + Diptera), itself consisting of two monophyletic sister-groups Mecoptera and Diptera.
Historically this diagram expresses the origin of the (supposed) sister-group relationship between the insect Order Mecoptera (Scorpion-flies) and the insect Order Diptera (Two-winged insects).
There is the period (or moment in time) in which a monophyletic group, such as the Diptera, has originated, that is, having branched off, by species-cleavage, from its sister-group (the Mecoptera). This period we call the period of origin t1 of the group (and also that of its sister-group), while we call the period in which the youngest common ancestor of the r e c e n t species of the group began to be engaged in repeated division the period of differentiation t2 of that group (see Figure above). Of course this is not meant to mean that in the preceding period no differentiation occurred, it just means that from this period onward the morpho-functional whole, as we at present know it with respect to the taxon concerned, was completed, while, of course additional development continues. Here we spoke about the time in which certain events (origin, differentiation) took place. If, on the other hand, we wish to emphasize how long ago it was that these events took place, then we call -- with HENNING, 1966 -- t1 the age of origin, and t2 the age of differentiation.
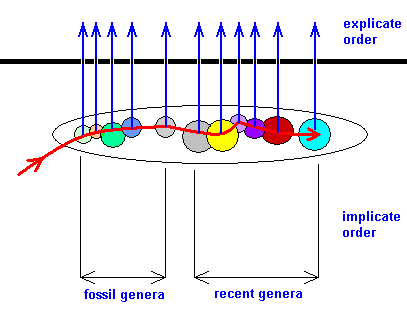
Figure 28 : Development of the morpho-functional groundplan of the recent (part of the) Order Diptera. The noëtic trajectory (red curve) enters the stability field (large ellips) of the order-groundplan of Diptera. In it, the trajectory successively enters the stability fields of the groundplans of dipterous genera (after having entered those of the corresponding families) which are only known as fossils. During this course the morpho-functional groundplan of recent Diptera is stepwise noëtically developed and completed, and the results are successively projected into the Explicate Order. And upon the noëtic trajectory entering the stability fields of the recent genera the corresponding noëtic results are successively projected and then seen in the Explicate Order as further differentiations within the morpho-functional groundplan of recent Diptera.
If we suppose that the Two-winged insects (Diptera) and the Scorpion-flies (Mecoptera) phylogenetically relate to each other as sister-groups, then it is very well conceivable that, say, at the time t1a (See Figure 27, above ) the representatives of the taxon 'Two-winged insects' still possessed four wings, and that two-wingedness and the transformation of the hindwings into halteres was accomplished only later on in the history of the taxon, for example not until time t1b, whereas another groundplan character was already accomplished much earlier. As a result of these findings HENNIG, in Die Stammesgeschichte der Insekten, 1969, has introduced the concept of stem-group, with which we will deal shortly.
It is theoretically fully legitimate to have the absolute age of a monophyletic group, that is, the age of origin (the time point of branching off from the sister-group, or, equivalently, the origin of the sister-group relation) be a measure of the absolute order of rank of that group ( the older it is, the 'higher up in the hierarchy' [= the lower in the phylogenetic system] it is, and thus the higher its rank is) and so directly showing with which other group it is comparable : First of all with its sister-group, which has originated not only simultaneously but also under the same conditions. Secondly, with other groups that have originated approximately at the same time and have developed under the same more global conditions. The significance of these considerations for paleo-biogeography is evident. Of course the time point of branching off from the sister-group is not the only important event for paleo-biogeography but does belong to its foundations.
Let us now see in what way the absolute age of a given monophyletic (not typological) group can be determined. Several methods are available for this of which by far the most important is the paleontological method.
In this method two things are presupposed :
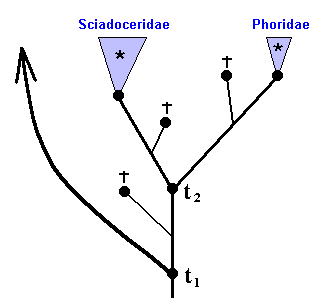
Three families from the insect Order Diptera form a monophyletic group ( This is supposed to be already determined) as follows :
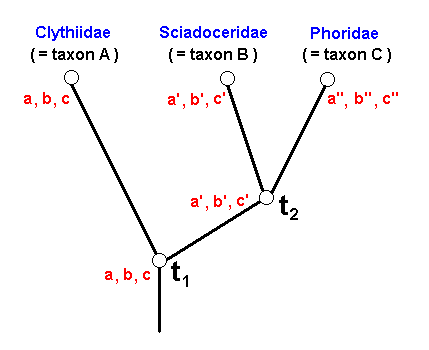
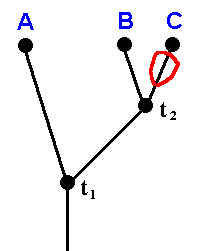
Figure 29 : Phylogenetic relationships between Clythiidae, Sciadoceridae, and Phoridae.
t1 is the time point of origin of the sister-group relationship between A and (B+C).
t2 is the time point of origin of the sister-group relationship between B and C.
The red letters indicate the stages of three transformation series :
a ==> a' ==> a''
b ==> b' ==> b''
c ==> c' ==> c''
The representatives of taxon B+C (Sciadoceridae + Phoridae) all possess character a in a derived state (namely a' and a'') with respect to the state of it possessed by all representatives of taxon A (Clythiidae) (Where it is already given that A+B+C is a monophyletic group, and that A is a monophyletic group). This makes it probable that also B+C is a monophyletic group. Moreover, all representatives of taxon C (Phoridae) possess character a in a derived state (a'' ) with respect to the state of it (a') possessed by the representatives of taxon B (Sciadoceridae). Therefore it is probable that taxon C is a monophyletic group. And because B and C are sister-groups, together forming the monophyletic group B+C, taxon B is also a monophyletic group.
These conclusions are strenghtened by the characters b and c, which are possessed in precisely the same way as character a is possessed by the members of the group A+B+C.
(See also next Figure, where it is given what the characters a, b, c and their derived states are.)
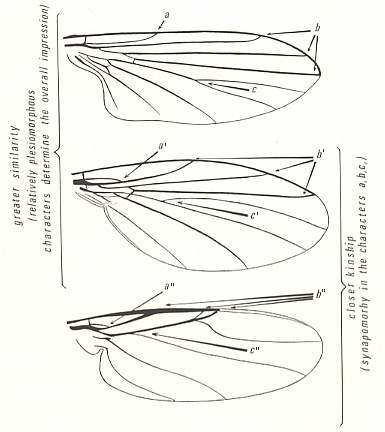
Figure 30 : Great difference between morphological similarity and phylogenetic relationship. Wings of the dipteran families Clythiidae (above), Sciadoceridae (middle), and Phoridae (below). sc = Subcosta, r1 = Radius, r2 = first branch of Radial Sector, r3 = second branch of Radial Sector, r4 = third branch of Radial Sector. Characters a, b, c and their derived states, as what these characters and states are, indicated by arrows.
Synapomorphy of the Sciadoceridae and Phoridae :
From the (recent) Clythiidae, through the (recent) Sciadoceridae, to the (recent) Phoridae, we can discern three transformation series
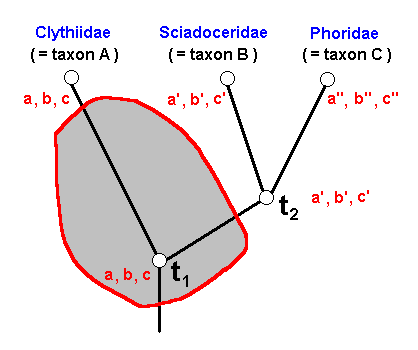
Figure 31 : Phylogenetic relationships between Clythiidae (A), Sciadoceridae (B), and Phoridae (C), as in Figure 29.
Area of possible taxonomic allocation within the group A+B+C of a fossil wing with 'clythiid venation'.
a, b, c, a', b', c', a'', b'', c'' states of characters a, b, and c (see Figure 30 ).
This uncertainty in determining the fossil results from the fact that we -- basing ourselves on the wing-venation of recent forms, including those of the sister-group of A+B+C -- must assume a venation like we see it in recent Clythiidae to be present already in more remote ancestors : The 'clythiid venation' is without change inherited from these ancestors. The allocation of the fossil to precisely the stem-species of the whole group corresponding to time point t1 is very improbable, because of the small chance of having precisely this fossil. The allocation of the fossil thus could be :
W're now going to find out how much bearing all this has to the determination of the minimum age of the taxa (See next Figure). We will consider the three just enumerated cases :
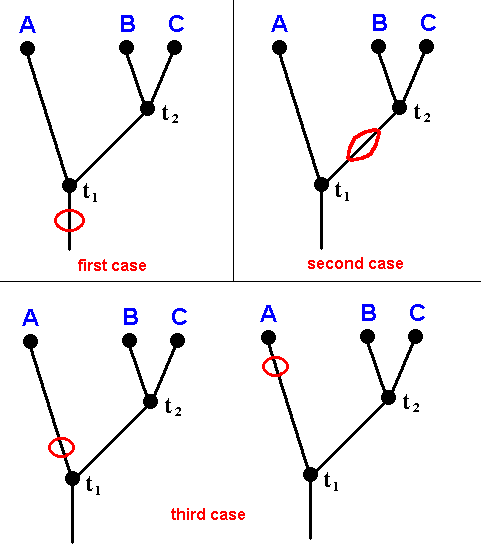
Figure 32 : Possible positions of the fossil dipterous wing with 'clythiid venation' with respect to the time points of species-cleavages t1 and t2 in the ancestral tree of the taxon Clythiidae (A) + Sciadoceridae (B) + Phoridae (C).
In the first case the time point t1 is later than the age of the fossil (= period of deposition of the layer carrying the fossil).
In the second case the time point t1 is before that which marks the age of the fossil.
In the third case the time point t1 also is before that which marks the age of the fossil, and perhaps also t2 is before it.
Accordingly, if we know the age of the fossil it does not give us anything with respect to the time points of (1) the Clythiidae, (2) the Sciadoceridae, and (3) the Phoridae. These cleavages could have taken place after the sediment layer containing the fossil was deposited (first case, and, with respect to B and C, second case), or before it was deposited (third case, either with respect to A and B+C, or with respect to A and B+C, and B and C ).
A further possibility is that the fossil (still our fossil wing with 'clythiid' venation) does not at all lie in the direct genealogic lineages of the taxa A, B, C, B+C, and A+B+C. One might think of the following possibilities, compiled in the next Figure :
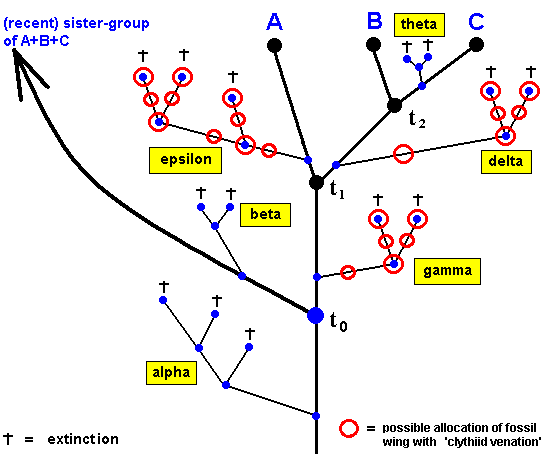
Figure 33 : Possible positions of the fossil dipterous wing with 'clythiid venation' with respect to the time points of species-cleavages t0 , t1 and t2 in the ancestral tree of the taxon Clythiidae (A) + Sciadoceridae (B) + Phoridae (C) with fictitious extinct side-branches ALPHA, BETA, GAMMA, DELTA, EPSILON, and THETA.
As has been said, the fossil wing with 'clythiid venation' might lie, not in the direct ancestral lineages of the group A+B+C, but in one or another (extinct) side-branch : a number of such possible positions of the fossil wing is indicated by small red circles.
Time point t1 marks the beginning of the period of differentiation of the taxon A+B+C, and at the same time the point in time of the origin of taxon B+C and taxon A.
Time point t0 marks the origin of taxon A+B+C and of its sister-group.
As indicated, we have here the (fictitious) side-branches ALPHA, BETA, GAMMA, DELTA, EPSILON, and THETA. To one or another of these the fossil might belong. We here have assumed (just as an example) that it either belongs to GAMMA, EPSILON, or DELTA.
The above cladogram (Figure 33) suggests that there was some sort of 'main stem' leading to the recent group A+B+C (and its sister-group), and that in addition to it this stem had a number of 'side-branches' that finally became extinct. But this is apparent only because we reason from the recent species. The group A+B+C might just as well be considered a mere side-branch of some main stem, which main stem, however, became extinct. Indeed, the cladogram might just as well look like this :
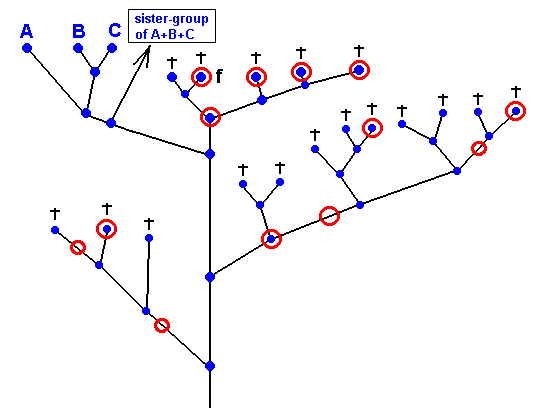
Figure 34 : Possible positions (red circles) of the fossil dipterous wing with 'clythiid venation' within the (alternative) cladographic pattern containing the group A+B+C (= Clythiidae + Sciadoceridae + Phoridae).
If the real position of the fossil wing (with 'clythiidae venation') turns out to be that of the species f in the above Figure, then we can assume (as just an example) that all the species marked with a purple circle -- see next Figure -- also possessed a 'clythiidae venation', while all the other species possessed one or another venation deviating from that of the Clythiidae :
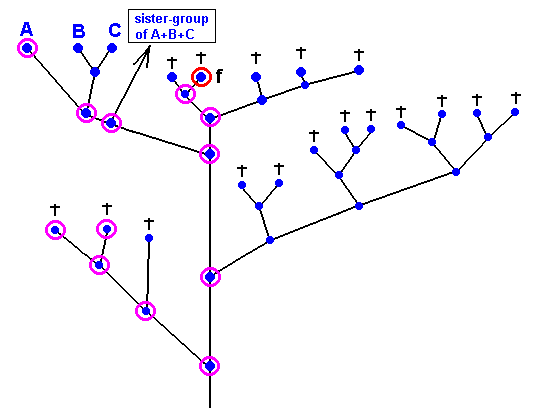
Figure 35 : The fossil dipterous wing with 'clythiid venation' is here assumed to belong to the (extinct) species marked with f. All the species -- within the (alternative) cladographic pattern (of Figure 34) containing the group A+B+C (= Clythiidae + Sciadoceridae + Phoridae) -- marked with a small purple circle are here supposed to possess clythiid wing-venation. All the remaining species are supposed to possess a wing-venation that deviates in one way or another from that of the Clythiidae.
Paleontological experience teaches us that almost all fossils that are actually discovered do not lie in one or another direct genealogical lineage of, and leading to, any recent species. These so-called 'direct genealogical lineages' do not have essential significance (that is, are not fundamental). They are only as such recognized after the fact with respect to the recent species. Many species (and 'lineages') have become extinct in the course of biological history, and only relatively few have continued one or another lineage. The conditions prevailing during the periods of the Earth' history have acted as a kind of filter through which only (relatively) few species could pass and fan out into the ecological landscape to occupy the many niches that had become free as a result of extinction.
Stem-group
The possibility that a given fossil can belong to some extinct side-branch brings us to the important concept of s t e m - g r o u p introduced by HENNIG in his magnificient book on fossil insects Die Stammesgeschichte der Insekten, 1969. We will now expound this concept.
For this, let us -- just as an instructive example -- consider the following phylogenesis (expressed as a cladogram) :
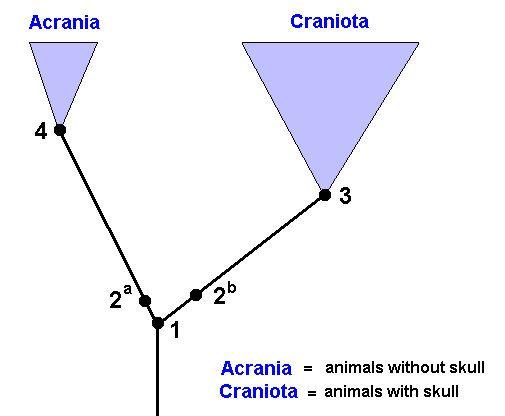
Figure 36 : Genealogic tree of the Vertebrates (Acrania + Craniota).
Species 1 is the youngest common stemspecies of the group Acrania + Craniota.
Species 4 is the youngest common stem-species of all recent Acrania. It possessed precisely that set of characters which we must presuppose as composing the groundplan of the (recent) taxon Acrania.
Species 3 is the youngest common stem-species of all recent Craniota. These recent Craniota possess the character set of species 3 (= the groundplan of the [recent] Craniota), or they show deviations from this character set ( All this can be found by studying recent representatives).
Species 1 has bifurcated into the daughter-species 2a and 2b that each for themselves, through new divisions, give rise to respectively the Acrania and the Craniota.
The groundplan of the Acrania is completed in the line between species 2a and species 4, while that of the Craniota is completed between species 2b and species 3.
Fossils, that, for example, show the complete groundplan as we have to suppose it to be in the Craniota, of course must be considered to belong to the Craniota if this groundplan contains apomorphous (= derived) characters (because these derived characters demonstrate that). And if these fossils already possess derived characters belonging to the groundplan of one of the subgroups of the Craniota, then they already belong to such a subgroup.
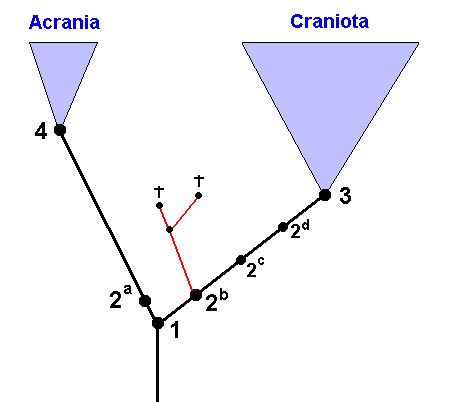
For the most recent common ancestor of that side-branch and the recent Craniota is still more recent than the most recent common ancestor of this side-branch and the recent Acrania. This can be easily verified in the above Figure.
Stem-group and *group
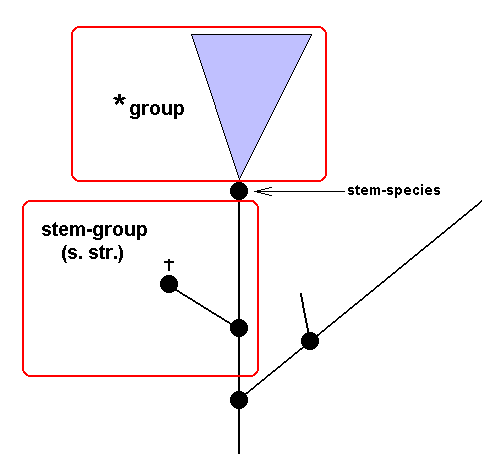
These distinctions are in fact of a typological nature, and they accordingly have no theoretical significance within phylogenetic systematics : The stem-group is a purely methodological concept, introduced to deal with the intrinsic incompleteness of the paleontological record : The stem-group is not a phylogenetic concept, it is not a taxon. On the other hand, the *group PLUS stem-species is a taxon, and also the stem-group PLUS stem-species PLUS *group.
If, in the identification of a fossil, it can positively be determined that it does not belong to the *group (but nevertheless belonging to the taxon to which the *group belongs, so, for example, not belonging to the *Craniota, but still to the Craniota), because, for instance, it is positively visible that one or more characters of the groundplan of the *group have not yet been developed (that is, that with respect to these [in contrast to other] characters the apomorphous [= derived] condition is still not present, whereas the plesiomorphous [= primitive] condition is ), then the conclusion "belongs to the stem-group (of the *group)" means that the stem-group is delineated (as in Figure 38 ).
Spurious stem-groups
When in paleontology one proceeds typologically (in contrast to phylogenetically), one will group together a number of fossils based on existing characters (In the context of Typology these are the characters that are supposed to show the intrinsic whatness of species and higher taxa), and one often will end up with a group as outlined in the next Figure.
On the absolute age of taxa (continued)
Having now expounded the concept "stem-group" (and also the concept "spurious stem-group") -- so important in Paleontology -- we return to the determination of absolute age of phylogenetic taxa with the help of fossils, using HENNIG's example of the fly families Clythiidae (taxon A), Sciadoceridae (taxon B), and the Phoridae (taxon C). We started to deal with this already above, see, for example, Figure 29 .
We had discussed a fossil wing having a 'clythiidae venation', and had found out that we could do only little with such a plesiomorphous fossil. Recall that the wing-venation of the representatives of the group A+B+C showed three transformation series, each transforming the venation as we go from Clythiidae, through Sciadoceridae, to Phoridae. Here the wing-venation of the Clythiidae is the most plesiomorph (primitive, original). If we had a fossil wing with 'clythiid venation', then there is only an area, not a point, in the phylogenetic system of A+B+C whereto we can allocate such a fossil. Recall Figure 31 .
Things become more promising when we have found fossils which have relevant apomorphous characters (instead of just plesiomorphous ones), that is, characters that are deviations from the groundplan of the whole group (A+B+C), or said more exactly, derived conditions of characters that are, in underived conditions, present in more primitive forms. For on the basis of such character states we directly can see whether one or more subgroups were already present at the time corresponding with the age of the fossil. When, for example, a wing impression (i.e. a fossil wing) has been found possessing the characters of the wings of the family Sciadoceridae, that is, a', b' and c' (See Figure 30 ), then we know that the time point t1 must lie temporally before the age of the fossil. The point in time that marks the age of the fossil is here, with respect to the origin of B+C a terminus post quem non (= a point after which, the origin did not take place) : The bifurcation into A and B+C has then already taken place :
It can still not be proved that this fossil wing with 'sciadoceridae venation' already belongs to the family Sciadoceridae, because it could still belong to precisely those species that are common to B and C, or it could belong already to the Phoridae. It is then -- when belonging to the Phoridae -- situated after the bifurcation into Sciadoceridae and Phoridae, and on the lineage leading to the stem-species of the recent Phoridae, where (on that lineage), however, the wing-venation had not yet reached that of the groundplan of the *Phoridae (that is, this transformation of the venation has yet to get started).
In summarizing, such a fossil can have belonged to one of the following parts of the genealogic tree (Figures 41, 42 and 43) :
It (that is, our fossil wing with 'sciadocerid venation') cannot belong to the *Phoridae, nor to the *Clythiidae, and also not to the stem-group of the Clythiidae, and also not to the stem-group (s.str.) of the Clythiidae + Sciadoceridae + Phoridae (= A+B+C), and, finally, also not to the stem-species of A+B+C.
Both time points t1 and t2 must lie further back in time than the period of deposition of the sediment layer with the fossil.
According to the nature of the second fossil here considered, that is, the wing impression having a 'sciadocerid venation' and thus possessing the character states a', b', and c', we have the age of the fossil (assumed to be known independently of the fossil) as a terminus post quem non for the origin, and thus the age, of the taxon B+C.
According to the nature of the last fossil here considered, that is, the wing impression having a 'phorid venation' and thus possessing the character states a'', b'', and c'', we have the age of the fossil (assumed to be known independently of the fossil) as a terminus post quem non for the origin, and thus the age, of the taxon C.
It is clear that if only we find a sufficient number of relevant fossils (of known age), we can at last also determine lowest time-boundaries (termini ante quem non), for example when we would find a fossil that with certainty belongs in the direct ascendency to the stem-species of a given taxon, that is, a fossil definitely belonging to the stem-group s.str., but not in a side-branch : Then we know that in the period in which the sediment layer with the wing to be fossilized was deposited the *group was not yet formed, or, which is the same thing, the subtaxa of the taxon were not yet formed.
As we said earlier, we can imagine that the better and the more completely we know the genealogic trees (cladograms) of organisms (established on a phylogenetic basis), included the ages of the taxa, the better it will be possible to feed back to precisely the age-determination of rock layers, now in turn based on these genealogic trees.
One of the important conclusions of our discussion is also the fact that only relatively apomorphous fossils count : A formal similarity of a fossil with (to stick with our example) the Clythiidae still does -- within the group A+B+C -- not say anything (more definite), it says at most something in a typological sense, not, however, in a phylogenetic sense.
The allocation of such a fossil wing to some type is then dependent on a mere contingent fragment of the character-set that has been preserved in the fossil, which in our case is the wing-venation, which for recent species has perhaps diagnostic value, but is perhaps not 'typical'.
The fact that in the venation of such a fossil 'clythiidae wing' do exist one or more apomorphies relative to insect wings outside the taxon A+B+C, is, in our considerations, presupposed, and with it, consequently, the fact that the fossil belongs to that taxon.
The case of the Clythiidae-, Sciadoceridae-, and Phoridae-wings also clearly demonstrates that 'overall similarity' does not a priori mean closer kinship in the phylogenetic sense (see again Figure 30 ) : The sciadocerid wing globally is more like the clythiid wing than like the phorid wing (comparing wings of recent representatives) (because the 'morphological distance' between at least b, c and b', c' is much smaller than that between b', c' and b'', c''), while the sciadocerid wing phylogenetically is closer to the phorid wing, than to the clythiid wing, on the basis of the synapomorphies a', a'', b', b'', and c', c'' (with respect to, respectively, the primitive conditions a, b, and c).
Interpretation of the genealogic tree of Clythiidae (A) + Sciadoceridae (B) + Phoridae (C) in terms of the Implicate Order, noëtic reactions, and projections.
The genealogic relationships between the Clythiidae, Sciadoceridae, and Phoridae, and their respective stem-groups, as discussed above, represent an instructive example to show in what way these findings can be translated in terms of the Implicate Order. According to our general theory the findings of phylogenetic systematics, as they are laid down in the form of a cladogram, do not express events in the Explicate Order but 'events' in the Implicate Order. In the Explicate Order we have the contingency of taxa (species and groups of species) getting either sooner or later extinct or persisting for some longer period of time. Therefore, as has been said, the answer to the question which part of the genealogic tree of a given taxon is its stem-group and stem-species, and which part is the *group, depends on which part of that taxon is actually present in the recent fauna (which presence is a contingent fact). The significance of the concepts of stem-group and *group, lies first of all in the practice of paleontology (as explained above) and, secondly, in recognizing the fact that a groundplan, as we encounter it in the recent representatives of the taxon, is, generally, developed step by step during the course of evolution. In all this we should not see the groundplan of the recent representatives as some sort of evolutionary goal toward which evolution was directed, because the definition such a groundplan, that is, its actual qualitative content that it has at the time, i.e. in recent times, is contingent. Let us show this in terms of the Implicate Order (in which nothing is contingent) :
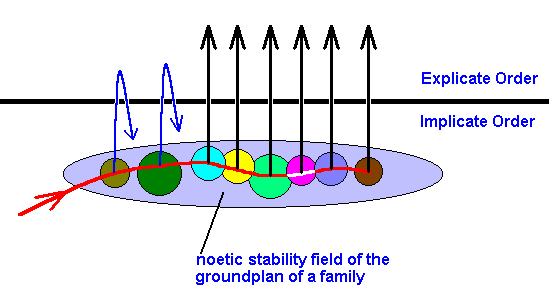
Figure 45 : Noëtic stability field of the groundplan of a given family. This field contains stability sub-fields : the stability fields of the groundplans of the genera of the family.
The noëtic trajectory (red curve) enters the stability field of the family-groundplan, and then takes its course successively through the stability fields of the genus-groundplans. The noëtic content of each of the latter reacts successively with the noëtic contents of potential ecological niches, followed by projection (a bundle of these from each genus, here drawn as a single line) of the reaction-products, resulting in the Explicate Order as the appearance of (individual instances of) several species, each one adapted to a particular ecological niche.
The persistence of these species in the Explicate Order is equivalent to the repeated projection and injection, that is, as long as projection-injection-projection-injection-projection- . . . of the mentioned reaction-product continues, the species continues existing in the Explicate Order.
In the present Diagram it is supposed that the alternation of projection-injection- comes to an end in the case of the species of the first two genera. This is symbolized by the 'returning' arrows. In the Explicate Order this means that the two genera become extinct, while the remaining genera persist (for the time being). The continuance of the alternation of projection-injection- depends on the relevant ecological conditions in the Explicate Order. A noëtic content (such as the product of some noëtic reaction) can only continue to be projected into the Explicate Order, if it carries with it the noëtic content of the proper existential conditions, that is, conditions for existing in the Explicate Order. In this way it is determined which part of the genealogic tree (which tree exists in the Implicate Order) 'now' actually exists in the Explicate Order (and which part has become extinct).
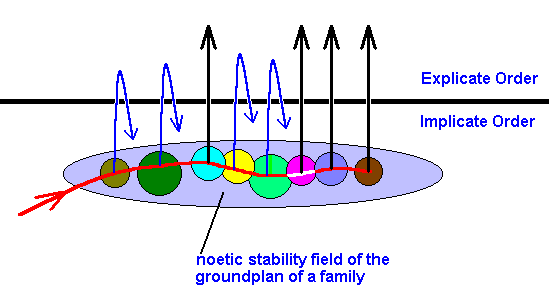
In the next Figure we see a different part of the genealogic tree 'now' existing in the Explicate Order :
Having said all this, we now will expound the 'translation' of the genealogic tree of the group Clythiidae + Sciadoceridae + Phoridae into the 'language' of the Implicate Order. But before we do so, we first indicate the lineages in the cladogram of this group, and then delineate the respective *groups, stem-groups and stem-species (Figures 47, 48, and 49).
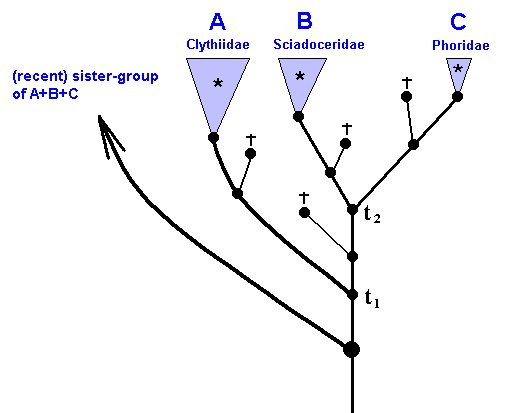
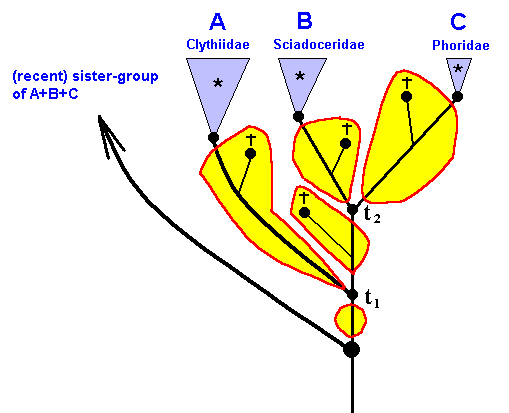
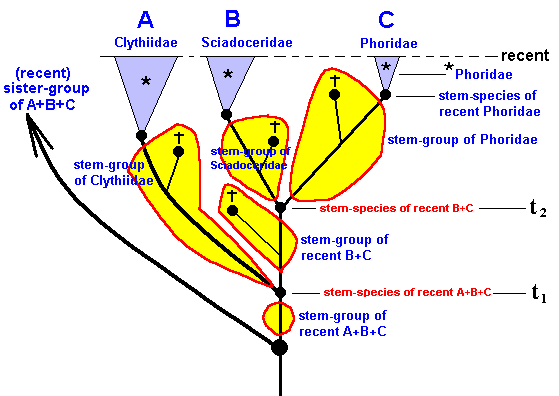
Figure 49 : Same as previous Figure. *groups, stem-species, and stem-groups.
There are three *groups : *Clythiidae, *Sciadoceridae, *Phoridae.
Further, there are three stem-species and three stem-groups : Those of recent Clythiidae, of recent Sciadoceridae, and of recent Phoridae.
There is the stem-species of B+C and of A+B+C. And, finally, there is the stem-group of B+C and of A+B+C.
We will now translate this into the terms of our theory :
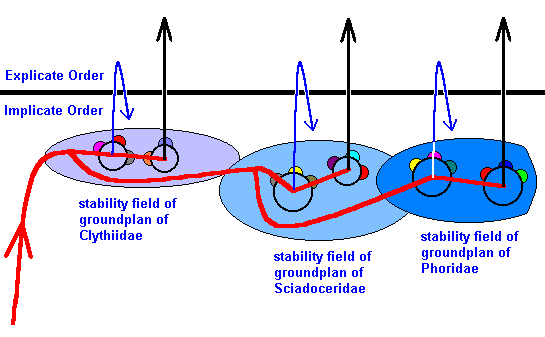
Figure 50 : Cladogram of previous Figures interpreted in terms of the Implicate Order, noëtic stability fields, noëtic reactions, and projections.
Indicated are the noëtic stability fields of the groundplans of the families Clythiidae, Sciadoceridae, and Phoridae. Further there are the stability sub-fields representing the stability fields of the genus-groundplans.
The number of genera in each family not necessarily corresponds to the true number of them (for our purposes this true number is immaterial). Also the number of (noëtic contents of the) potential ecological niches with which each groundplan noëtically reacts, and consequently the number of species of any one genus, does not necessarily correspond to the actual number of species (also this actual number is immaterial in the present context). Further the bundle of projections emanating from each genus (corresponding with the fictitious number of species) is drawn as a single straight or returning arrow.
The stability field of each family-groundplan contains two subfields of generic groundplans. As has been said, the number "two" is just symbolically meant, that is, it does not point to anything special.
In each family-groundplan stability field the first subfield (together with the not-generically differentiated part of the former) is the noëtic counterpart of the stem-group of the family. The second subfield is the noëtic counterpart of the *family.
By referring to our present Figure we can trace the course of events :
The noëtic trajectory (red line) enters the stability field of the groundplan of the family Clythiidae. Then it bifurcates. One branch of it traverses the stability subfield of the groundplan of the first genus. As drawn, this one genus and the projections emanating from it symbolize the noëtic counterpart of the stem-group of the family Clythiidae. After leaving this stability subfield, the noëtic trajectory then visits the stability subfield of the groundplan of the second genus. As drawn, this second genus and the projections emanating from it symbolize the noëtic counterpart of the *Clythiidae. We have now described the course of the first branch of the noëtic trajectory.
The second branch runs through the 'general' Clythiidae family-groundplan stability field and then enters the stability field of the groundplan of the family Sciadoceridae and again bifurcates. One branch of this bifurcation then successively visits the two stability subfields of the groundplan of the family's genera (also, for clarity, assumed to be two), where the first of them and the projection emanating from it symbolize the stem-group of the family Sciadoceridae, whereas the second one symbolizes the *Sciadoceridae.
The second branch of this bifurcation of the noëtic trajectory runs through the 'general' Sciadoceridae family-groundplan stability field, and, after having entered the stability field of the groundplan of the family Phoridae it successively runs through the noëtic counterpart of the stem-group of the family Phoridae and the noëtic counterpart of the *Phoridae.
In our noëtic interpretation of the evolution within the group Clythiidae (A) + Sciadoceridae (B) + Phoridae (C) we see that the 'cladogenesis' as established by phylogenetic systematics takes place, not in the Explicate Order but in the Implicate Order. Indeed, we recognise the genealogic tree (cladogram) of phylogenetic systematics in the course of the noëtic trajectory in the Implicate Order. The results of phylogenetic systematics as to the evolution of A+B+C is here supposed to be based on the three morphological transformation series a ==> a' ==> a'', b ==> b' ==> b'', and c ==> c' ==> c'', in the wing-venation. Of course the conclusion will be strengthened when other such transformation series are found, especially in other parts of the body of these flies or in that of the larvae. For the time being, however, we concentrate on the findings in the wing-venation and assume these to be sufficient to warrant our conclusion about the phylogenetic relationships between these three families. Let us put the elements of the mentioned venational transformation series in their proper places in the above Figure :
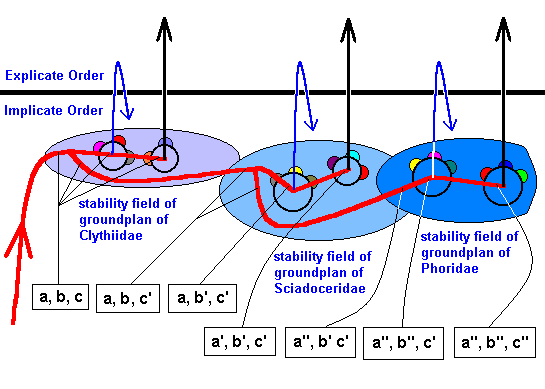
Figure 51 : Same as previous Figure.
The transformation a, b, c ==> a', b', c' ==> a'', b'', c'' is indicated ( The details of the course of this transformation, as given here, are fictitious, they just serve as an example).
Absolute order of rank of phylogenetic taxa.
Possible objections to Phylogenetic Systematics
The most important objection to Phylogenetic Systematics is the fact that it depicts evolution as a temporal succession of species-cleavages. We have already discussed this objection : If evolution consisted entirely of species-cleavages, then the transformation of organisms must largely be a cumulative result of a succession of such cleavages. This is because more or less permanent changes are supposed to be genetic changes. Such genetic changes are supposed to take place as a result of isolation and separation of parts of a given population. The selection pressures then will be different for such parts, that is, in different partial populations different random genetic mutations will have survival value, and so these parts will develop differently until there is complete reproductive isolation, that is, the formation of different species from the original species, or said differently, the individuals of the set of separated populations do not, and cannot, interbreed anymore. And because we can legitimately assume that the precise moments at which we will have the several reproductive isolations are not all simultaneous, we can hold that species-cleavage is always dichotomous, albeit that there will be many cases of a rapid following-up of dichotomous cleavages, not as such discernible by the methods of science. Thus because phylogenetic systematics sees evolution exclusively as a temporal succession of species-cleavages, the main factor for it that determines evolutionary change is random genetic mutation combined with natural selection.
However, we have seen that random genetic mutation and natural selection of advantageous mutants cannot be the main driving force in evolution, because every morphological change generally must involve, not a single mutation but a whole series of such mutations taking place in the right order at the right time without being detrimental to other processes in the organism (because that would not encourage natural selection of the mutants). Further there are important limitations of random genetic mutations to drive evolution coming from the microbiological and genetical point of view as is convincingly expounded by BEHE, 2007. And, as has been said, this was the main reason for us to transpose the evolutionary process within an altogether different ontological domain, the Implicate Order. And transposed in this way it has turned out that phylogenetic systematics can still have significance in our theory of evolution, at least insofar as at least many of its cladograms are recovered in the Implicate Order. And thus, with a re-interpretation of monophyly, we can still do phylogenetic systematics.
By involving the Implicate Order, another problem is at least partially and preliminarily solved : If evolution is nothing more than a series of species-cleavages, then there cannot be the evolution and succession of organic groundplans. As we have seen, the organic groundplans generally have little or no functional or ecological significance. Nevertheless they have appeared on the evolutionary scene. In conventional theory, and also in phylogenetic systematics, the origin of new characters or of new character states is closely connected with genetic mutants that are favored by natural selection. They are favored, because the changes of the mutants, expressed in the phenotype, endow the individuals with increased chances of survival (and this is their being selected). This means that the mentioned new characters or character states must be highly funcional in some relevant ecological context. So it is clear that an evolution that is exclusively driven by random genetic mutation and natural selection (as presupposed by contemporary biology including phylogenetic systematics) cannot produce the series of different organic groundplans as we know them to exist. At least a 'rationale' for such a production is provided by our noëtic theory, by presupposing that such groundplans noëtically already exist alongside all other noëtically possible patterns or structures in the Implicate Order, and that some analogue of dynamics (in our theory called the noëtic trajectory ) in that Order successively lets these groundplans being projected into the Explicate Order, during which projections they are unfolded along the Explicate Order's space and time dimensions.
There are other objections raised against phylogenetic systematics, but these all boil down to a defence of typological systematics in favor of the former (without explicitly assuming a second Order of Reality as we have done). These objections are important for getting a correct distinction between phylogenetics and typology, and are especially important to us because in our noëtic interpretation of evolution typology plays a role too. So let us deal with these objections. They group around the following items :
1. The persisting stem-species.
A complaint often levelled against phylogenetic systematics is that, with respect to species-cleavage, it proceeds unrealistic and artificially. One then has in mind the following case : A given organismic species persists during long periods in which it, from time to time, produces an offshoot without itself undergoing any morphological change or at most only a minor one. Diagrammatically expressed :
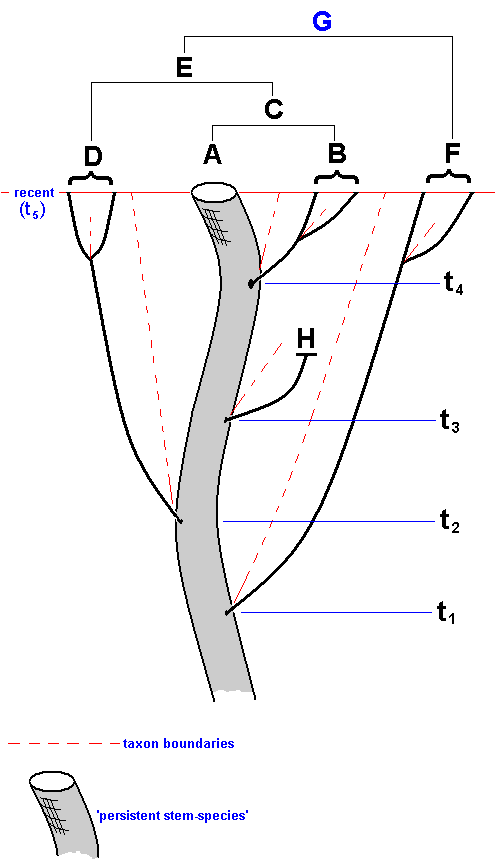
Figure 52 : Fictitious example of a splitting-off sequence along a persisting stem-species. t 1 -- 5 : time points. A, B, C, D, E, F, G, H -- taxa. See also next Figure, which is a phylogenetic rendition of the same events, for comparison.
This is the well-known case of a 'living fossil', a relict form, or the 'persistence of a stem-species'. Phylogenetic systematics, however, interprets this phenomenon quite differently, namely such that it interprets this phenomenon 'away' : Phylogenetic systematics, namely, exclusively investigates the order of bifurcation (without, of course, denying other evolutionary events !). It partitions the 'persistent stem-species' into taxonomically non-equivalent elements, resulting in the continuity of the stem-species to be completely lost. See next Figure.
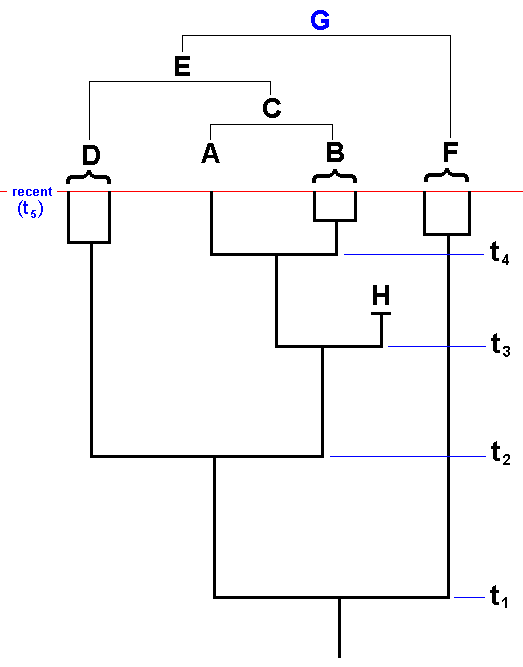
Figure 53 : Phylogenetic interpretation of the genealogic tree of the previous Figure ( Figure 52 ). What counts are the sister-group relations.
And this is held to be unrealistic, as a result of which phylogenetic systematics would have an incorrect point of departure.
This genetic interpretation of the species and speciation in which bifurcation as a result of isolation is central, also implies that we should not consider the population that remains, after a part of the original population has split off, to be conspecific with the original population (i.e. to remain the same species), because to distinguish between (different) species must take place in the horizontal dimension (ecological and geographic isolation resulting in intrinsic genetic divergence) rather than in the vertical dimension (time dimension) (the taxon- and species boundaries run vertically). In accordance with all this the initial species disappears and is replaced by two new daughter species. This is consistent with the fact that the remaining population must, after bifurcation, have become genetically different, because that what has been split off is always a non-random part of the gene pool.
In the typological view of the persistent identity of the 'stem-species that lives on' moreover appears a strongly contingent element in the reasoning, because, namely, the alleged 'persistent species' can be followed into the past only by means of fossils. The type (= here with respect to the morphologically 'persistent stem-species') is indeed conceived with respect to precisely that fortuitous section out of the (holo)morphe of the organisms that happens to be fossilized. For even if we know recent representatives, then still the assertion that the so-called 'persistent species' extends far into the past, and that this is then a stem-species that lives on, despite the many off-shoots, rests on precisely those characters that can be observed, that is, that have been preserved, in that remote past (In Molluscs, for example, the characters of the shell). The type allocation thus depends on taphonomic conditions [conditions of burial] which only very conditionally relate to the fundamental organization of the living organism : The type allocation brings only one, or just a few, not a priori fundamental aspects of this organization into the foreground, under exclusion, out of sheer necessity, of all remaining aspects of it, resulting in our obtaining a distorted picture, that is, a 'distorted type'. Moreover, we have the fact that it is more or less subjectively determined that some characters from the available set are considered to be more significant than others. But this is for the time being unavoidable. If such a subjective status of evaluating characters is recognized it will stimulate efforts to render it more objective. Indeed, the observed morphological continuity in the 'persistent stem-species' should not go unnoticed or ignored. It, however, should be considered outside phylogenetic systematics, for example in a natural philosphy of type development. It is in fact what we do with our noëtic theory of groundplans.
We must always realize that phylogenetic systematics designs a system only made up of phyletic bifurcation lines, clades. Morphological data are here first of all only a means. They are not the direct objects of such a system. In typological systems, on the other hand, it is not the clades but the grades that are central. And the clades and the grades can, it is true, be brought together from a methodological viewpoint, they should, however, not be confused qualitatively.
In the discussion of the typological and phylogenetic interpretation of a so-called 'persistent stem-species' I had, in Figure 52 and Figure 53 , added a fictitious extinct branch : I considered this extinct branch H as belonging to the stem-group s.str. of C. Here the practical usefulness of the concept 'stem-group' is again evident :
Before the (fictitious) discovery of the fossil representing H, the taxa C and D were ' thought of ' as sister-groups (that is the phylogenetic consideration), that is, the taxon that is most closely related to C is D, which, in the classification, is a good reason to unite these two taxa to represent one super-ordinate taxon C+D. After discovery of the fossil representing H, and after the establisment of the genealogical relation of H with other forms (Figure 52 and Figure 53 ), we are now forced to hold that the closest relative of C is not D but H. And now of taxon C+H (instead of C) taxon D is the closest relative. So the sister-group relation now becomes : C+H -- D (instead of C -- D). And this correction must be inserted into the classification. But because the interpretation of a fossil often needs to be amended as a result of further enquiry, this whole course of things is very impractical and, moreover confusing (but nevertheless necessary), in inclusive classification, that is, in setting up a classification that includes all forms, recent and fossil. The concept 'stem-group' provides a way out of this : In such a case we still maintain that the taxon that is most closely related to C is the taxon D, and that the fossil taxon H belongs in the stem-group (s.str.) of taxon C. Theoretically and phylogenetically this is completely correct and unequivocal and thus theoretically flawless, while at the same time having created a much less laborious procedure : We always place fossils into the stem-group of the relevant taxon which itself consists of recent representatives. If a fossil belongs to the *group (that is, not to the stem-group s.str.), then it also belongs to the stem-group of some subordinate (lower) taxon within that *group.
Again I would like to emphasize that the concept of 'stem-group' has no theoretic (and also no phylogenetic) significance. The practical significance lies in the fact that for fossils we do not have to create new taxa : A fossil (which thus is lying back in time) always belongs to a stem-group of one or another recent taxon. And this is the taxon to which the fossil is most closely (phylogenetically) related, established with the help of synapomorphies.
Crocodile tears
That, according to phylogenetic systematics, the Crocodiles are more closely related to the Birds than to the remaining reptiles for many went down the wrong way : This results, so they say, in a line of division straight across the (alleged taxon) Reptilia -- namely between the Crocodylia on the one hand, and the Rhynchocephalia + Squamata on the other (here I waive from considering the uncertain status of the Chelonia) -- merely in virtue of the fact that from the Reptilia the Birds have evolved. If the latter had never evolved one never would have drawn such a line, that is to say, one would interpret these same Reptilia differently.
To this objection I answer by saying that if Birds never had evolved (for methodological reasons I pass over the origin of Mammals), then there indeed would not exist a group (taxon) outside the reptiles to which the crocodiles would be more closely related than to the remaining reptiles. But precisely because from a subgroup of the Reptilia the Birds have evolved, the Reptilia now form an artificial group when we exclude the Birds from it while we know them to exist. Precisely because birds have actually evolved from poikilothermic scale-bearing animals that lay their eggs on land, these 'Reptilia' are phylogenetically not the same anymore as they were before the branching off of the Birds. But that they nevertheless (also phylogenetically) still figure within a unity is also expressed by their phylogenetic systematics : Together with the Birds they constitute the taxon Sauropsida : The Crocodylia (as being a subtaxon of the Sauropsida) -- connected, it is true, with the Birds, and so forming the taxon Archosauromorpha -- still are (also) connected with the remaining reptiles (in which the status of the Chelonia (Testudines) remains uncertain) :
Figure 54 : Phylogeny of the Sauropsida (Reptiles and Birds [Aves] ) according to some recent (with respect to 1975) authors.
(After HENNIG, 1975)
Figure 55 : Same as previous Figure : Phylogeny of the Sauropsida (Reptiles and Birds). If we leave out the Birds (Aves), then we have the Reptilia (yellow). But these 'Reptilia' do not form a monophyletic group anymore because they are not the only descendants of their common ancestor. They are thus a mere paraphyletic group, despite the fact that they represent a definite holomorphological type or groundplan different from that of the Birds.
The fact that the birds have developed divergently with respect to the rest of the Sauropsida (feathers, homoiothermic physiology) as such has nothing to do with phylogeny (= sequence of species-cleavages, cladogenesis) but with (holo)morphology : how differently the birds will ever develop, it does not influence in anyway their phylogenetic kinship with the crocodiles! Genealogic kinship always has meant : recency of common ancestry. The divergent morphology is only methodologically relevant to phylogenetic systematics : By way of the transformation series scales ==> feathers (= derived scales) and also those transformation series more directly connected with homoiothermic physiology (that is, how it has been evolutionarily developed in especially birds) the birds show to possess synapomorphies (with respect to the rest of the Sauropsida) that form a strong indication that the birds are more closely phylogenetically related to each other than to other animals, while other synapomorphies indicate a closer phylogenetic kinship of this monophyletic group Aves with the monophyletic group Crocodylia than with any other monophyletic group.
Contradicting phylogenetic designs (proposals) of genealogic trees (cladograms).
The fact that phylogenetic trees of a given animal group worked out by different authors can turn out to be different appears to strengthen the general objection against phylogenetic systematics. When two systems, both based on the principles of phylogenetic systematics contradict each other (which is, for example, the case in the system of moths and butterflies), one might see a proof that also phylogenetic systematics does not lead to definitive solutions and authors still disagree, resulting in the fact that phylogenetic systematics does, in this respect, not represent an improvement against typological systematics.
However, this critique levelled against the (priority claim of) phylogenetic systematics, forgets that the differences of opinions, as expressed in the proposed phylogenetic systems, appear as a result of contradicting empirical findings (in contrast to presuppositions) concerning, for example, the direction of certain character transformations (is a' more primitive, that is, less derived, than a, or is a' derived with respect to a?), and these problems can be objectively discussed and investigated, while in typological systematics some investigators hold character X to be 'determining', while other investigators hold character Y, and not X, to be 'determining', 'characteristic', 'typical', or 'essential'. And as long as these terms maintain their vagueness such differences of opinion cannot be settled by discussion or even observation.
Polyphyly
In fact, only one form of critique against phylogenetic systematics would be possible. It should be based on the assertion that the whole of the recent world of organisms is polyphyletic. Not only higher but also lower taxa have polyphyletically developed. In that case there indeed would be no phylogenesis in the sense of the formation of sister-groups and monophyletic groups. Such a polyphyly has been extensively and impressively defended by Ignaz LICHTIG in 1938 (Die Entstehung des Lebens durch stetige Schöpfung). However, subsequent developments, especially in biochemistry, genetics, and molecular biology, could not confirm such a polyphyly.
Of course there do exist groups (of organisms) that are polyphyletic, that is, their members having not descended from a single common ancestral species but from a number of them. But in phylogenetic systematics such a group will not be acknowledged as a true phylogenetic taxon. We must admit, however, that there could very well exist much more of such groups than is generally assumed by phylogenetic systematics (Indeed, we ourselves hold that monophyly is only to be found within the confines of the Implicate Order, and that it is parallel instead of converging lines, that we find in the Explicate Order).
With this we conclude our exposition of possible critique that could be levelled against phylogenetic systematics.
In the next document we continue, and conclude, our discussion of the generalities of the evolution of groundplans involving the theory of phylogenetic systematics.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXV-C.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXIV-A