
Predatory egg-eating (ecto-oophagous) phase
From precisely that moment when the attractive action on the female of the primitive terebrant was coming, not from the gall, but from the egg (in the gall) of the (potential) host, the significance of the gall as a place to lay the egg of the terebrant lost its obligation. As a result of this, the females of the primitive terebrants gained a new possibility : To specialize on finding host-eggs that occur in various ecological conditions.
Such, first of all, can be eggs laid in conditions similar to the original conditions, that is, also under the surface of living plants, but already without obligatory gall-like out-of-control growth of the tissue around them. Such clutches of eggs might be, for example, those of wood crickets, cicadas, some bugs (Hemiptera), etc. Attractive objects were, we must assume, also eggs of ectophytic saw-flies not causing galls. It is very probable that precisely against attacks by primitive terebrants, in certain ectophytic saw-flies special habits have been developed, for instance the habit of the female of the Australian saw-fly Perga lewisi Westw. to guard the double egg row, laid by it into the upper integument of Eucalyptus leaves.
But not only proper clutches of eggs of host-insects, placed under the surface of plants, but also those laid in other conditions -- in special containers, or directly on various parts of plants -- could attract females of primitive terebrants and become typical sites for egg-laying. Such are egg-capsules of cockroaches, the oothecs (egg-pouches) of praying mantises, egg-packs of spiders, piles of eggs of worms.
In this phase of the establishment of terebrants were, in this way, created especially favorable conditions for the transition of terebrants from their original host-sawflies to insects belonging to various Orders, and in certain cases to spiders and even to ticks and centipedes.
In these new conditions, in laying the eggs into clutches of eggs of the host, the larva of the terebrant, devouring one egg after another, obtained the possibility to live exclusively of the egg-stage of the host. In the case where the infected clutch of eggs was large enough the terebrant reached its complete development. In the opposite case -- malnutrition in small clutches of eggs -- the size of the terebrants correspondingly decreased, which could not fail to be expressed essentially in its overall development, resulting for instance in the reduction of (the) wing-veins.
On the other hand, already more or less (qua size) diminished forms of terebrants, infecting larger egg-clutches, obtained a new opportunity : Laying a number of eggs into a single egg-clutch of the host and developing in it together, that is, we might say, to lead a predatory semi-family life. In this case, the larvae of the terebrant, could, after having destroyed [i.e. largely consumed] one egg-clutch of the host, sometimes gnaw channels of a length of some centimeters in the pith of the plant looking for a neighboring clutch in order to complete their feeding in it.
Living in this or that way exclusively of the content of eggs of the host, the terebrant-egg-eating larva does not constipate its intestine with rigid solid plant remains, as it was the case in the original inquilinoid phase. Now the larva does not need anymore to give off excrements during development, which gave it also the possibility to preserve up to a certain degree the embryonic state of its intestine and the absence of a connection between the middle- and hind-gut, as it today is typical for almost all terebrants, except for, as we saw, the typical archaic inquilines.
In this way originated and developed the predatory egg-eating, or ecto-oophagous phase of [the evolution of] the Terebrantia.
It is remarkable that these primitive relationships are again encountered in evanoid terebrants, that is, in the second family of them, the Evaniidae. For a representative of this family see next Figure :
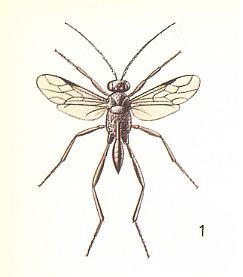
Thus, according to the substantial investigations of Genieyes, 1924, the evaniid Zeuxevania splendidula Costa. develops in the egg-capsules of the southern cockroach Loboptera decipiens Géné. The egg of the terebrant is laid inside one of the eggs of the capsule when the latter is not yet completely hardened. It [the egg] has a cylindrical shape, markedly narrowed at one pole and ends with a tiny stalk. The length of the egg is 1 mm and the width is 0.25 mm. The first-instar larva is almost cylindrical with 13 clear segments, with simple strongly projecting mandibles and with an internal tracheal system without spiracles (= external openings). In this state of feeding the larva of the evaniid limits itself to the content of only one egg of the cockroach, but after shedding its skin [thus being now a second-instar larva] it attacks the other eggs lying nearby. So, in the first stage (instar) it behaves like an internal parasite, but after that becomes a predator. In the following stages it assumes an almost spherical shape and has a large projecting head. The larva has now 9 pairs of spiracles, but its integument lacks spines and other structural features. Only one single evaniid larva can develop in one egg-capsule of the cockroach. Hibernation takes place in the stage of a fully-grown larva inside the capsule. Emergence of the adult and liberation into the open takes place in the spring or in the beginning of the summer.
In the next document we will consider the next evolutionary phase of the Hymenoptera, the Parasitic egg-eating (endo-oophagous) phase.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXXVI.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV