
When with the construction of a preliminary accommodation for the collections of provisions the wasps began to bring in smaller-sized prey, but in larger numbers, the selection of their prey, as already indicated, strongly broadened. It is wholly natural that the until now developed ability to paralyze one large prey turned out to be insufficient.
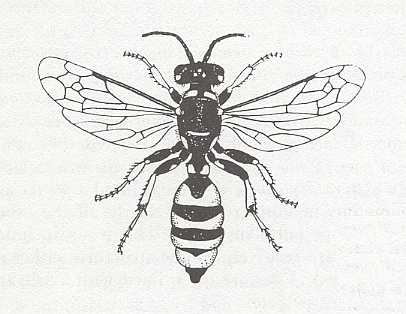
Figure 1 : Cerceris arenaria, female. ca 15 mm. Sphecidae.
(After RICHARDS and DAVIES, Imms' General Textbook of Entomology, 1977)

Figure 2 : A female digger wasp of the genus Cerceris, arriving near her nest with a paralyzed weevil.
(After EVANS and EBERHARD, 1973, Photograph by Edward S. Ross)
However, already FABRE noted that the hunt for small and delicate 'game' sometimes simplifies the complex paralyzing operation to one single sting [act], as is, for example the case in the paralyzation of small caterpillars by the ammophile Ammophila holosericea F.
Ammophila holosericea (" l'Ammophile soyeuse") of Fabre, according to BERLAND, 1925, is in fact A. heideni Dahlb., whereas A. argentata of Fabre is in fact Psammophila tydei le Guillon. Also Sphex flavipennis of Fabre is in fact S. maxillosus F.
He also indicated that when the prey has wings, and could therefore easily escape, the catch of it must proceed with such a speed that does not allow the [separate] stinging acts [at one and the same prey] to be regulated. The wasp stings the prey (horse-flies [Tabanidae] and the like), "but where it was applied, was on that part of the body of the prey which happened to be within reach [of the sting] during the struggle".
So, in the present Phase of the evolutionary development a part of the preys being brought to the cell turned out not to fully correspond to that particular purpose for which they were selected in the first place -- to serve as a living or at least fresh store of provisions for the nursed generation.
Some of these preys are, as a result of the accidental nature of the struggle or of an extreme concentration of the nervous system of the prey, or of its general weakness and delicateness, or by whatever other reasons, in such a strong degree vulnerable to the action of the venom that being stung by a wasp for them became lethal. This is especially true of small and delicate prey such as aphids, midges, small flies, [small] cicadas. In addition, in these new conditions of hunting the stinging acts became more rude and in certain cases even a special application was [evolutionarily] given to them. Thus, Oxybelus uniglumis L. and O. melancholicus Chevr. sting a fly in the region of the throat, but then the sting is not withdrawn, but, after having turned the pinned-up prey in a more convenient position, carry it to the nest. In such a way also certain Crabro's behave. See next Figure.
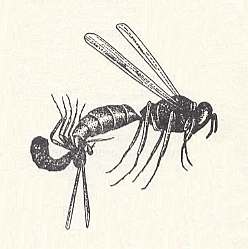
Figure 3 : A Crabro (Crossocerus) elongatus Lind with a prey, pinned on its sting.
(After NIELSEN, 1933, in MALYSHEV, 1966)
Other preys, on the contrary, thanks, perhaps, to the properties of their integument, turned out to be so little sensitive to being stung, that they soon recover and sometimes even again become able to move and fly. Thus ADLERZ observed that the seized, by a bembex-wasp, fly Volucella bombylans L., before it was carried to the nest managed to free itself and flew off and again being chased by the wasp. So, in one and the same cell, in addition to more or less paralyzed preys, easily can be encountered completely dead ones as well as living, wholly motile ones (flies, spiders). All these cases were of great significance to further transformation of the provisioning instinct in wasps.
(constructing a nest, catching the first prey, bringing it into the nest, laying an egg onto it, catching a second prey, bringing it into the nest, closing the nest)] (See for the meaning of the symbols HERE (Part XLIX) )
In such conditions easily might originate a situation in which the provisioning of the given cell was continued when from the egg, laid on the first prey in this cell, the wasp-larva had already emerged and even was ready to begin feeding [That is, provisioning did not stop after the wasp having oviposited onto the one prey brought in. It continues, which marks the beginning of progressive provisioning].
As a fine example of this may serve the comparison of Ammophila urnaria Cress., not thoroughly paralyzing its prey, with its relatives bringing in not more than one prey into a cell. After having prepared a burrow, usually one or two days later the wasps places in it the first caterpillar. In that period the egg, ready for being laid onto the given caterpillar, progresses a little in its development [So in this way the wasp lays an egg from which the larvae will soon emerge. And this larva then finds a still fresh prey]. The wasp brings in the second caterpillar into that same cell again not directly, but sometimes three days later, only after the hatching of the larva from the egg laid onto the first prey. In the last case the second caterpillar is, at the time of it being eaten, still more fresh than it would be when it was brought in directly after the first one, right after oviposition. How much, in this Phase of evolution, the care of the wasp is in keeping fresh the food of the larvae, is shown by the fact that A. campestris Latr. and A. heideni Dahlb. in their visiting their being-fed larva throw out from the nest wrinkled caterpillars and replace them by new ones. This fact is, perhaps, as significant as was the origin of the instinct to close the nest.
According to ROUBAUD, 1910, the inspection by the wasp of the stores of provisions in the cell during the time of growth of the larva protects the latter from parasites (tachinids and other such parasites), accidentally brought into the cell together with provisions. Therefore "progressive" (slowed-down) provisioning is preferred above mass (stepped up) provisioning.
The fact that we here witness the very origin of a new form of provisioning, that is, a provisioning being slowed-down and thus now taking place from day to day, is visible in the instability of the method of preparing provisions in everytime a same species of Ammophila. Thus A. heideni on the isle of Corsica (France) provisions the cell after the larva has emerged, whereas in Algeria it does it before that. In the same way, A. campestris Jur., depending on the place, provisions the cell before the hatching of the larva, or does this after the hatching.
According to the special investigations of BAERENDS, 1941, in Holland, this wasp also there applies slowed-down provisioning of the cell. Having made a short burrow with a widening at its end, the ammophile closes it with a temporary cap (consisting of individual particles, small lumps of soil, etc.). Then it goes out hunting, brings a caterpillar into the cell, lays an egg onto it, and again closes the nest temporarily. Then, during several days the wasp usually does not visit the nest. After this, the wasps enters the cell, but without a caterpillar. At this time the larva has emerged, which usually, in favorable weather conditions, takes place already 48 hours after oviposition. Now the wasp again goes out for hunting, bringing in caterpillars, sometimes alternating this work, taking a number of days, with visiting the nest without a caterpillar, and closing it with a temporary cap. After having delivered the last (usually 7th) caterpillar the nest is closed definitively.
In this way, the order of actions in the present Phase is basically analogous to the order which was worked out in the previous Phase. Here [in the present Phase] only the time of provisioning of the cell has changed. In a general form this might be expressed such (See for the meaning of the symbols HERE (Part XLIX) ) :
(construction of nest, provisioning, oviposition, repeated provisioning over t days, closure of the nest)
where t is the number of days of provisioning of the cell after oviposition, whereas in the previous Phase t = 1 : B + AC + D + (AC)n + E.
Here, apparently, also fit those cases in which one and the same species at one and the same place (locality) and at the same time brings into the cell one to three caterpillars depending on the size of the wasp itself. So behaves, for example, A. procera Dahlb., according to HARTMAN, 1905. In all these phenomena we should see a kind of divergence of characters and the birth of new species.
When a wasp hunts for a prey that lives concealed and is not therefore noticeable, we can easily conclude that the slowing-down in provisioning here results either from the hardship of the hunt for such a prey, or from the difficulty to find it. But if we want to see the real picture, we must focus on those clear cases in which, firstly, the hunt is for preys that live in great abundance in those places where also the wasp lives, and secondly, cases in which the weather conditions are fully favorable for hunting. Then it becomes clear that here the cause of the slow-down is hidden in something else -- in the inherited habit [or structure] of vespine life, established in the course of that life's historical development. This habit is, in addition, intimately coordinated with the maturation of the eggs in the organism of the wasp and with the rates of laying them.
As a good example of all this may serve the hunting instincts of the bembexes Bembex rostrata L. and those similar to it, in which the slowed-down provisioning of the cell is well developed. [The name "Bembex" is probably synonymous with the name "Bembix" : In Imms' General Textbook of Entomology, 1977, we read : "Bembix rostrata" ("Bembex" is not mentioned there at all). In the Elsevier guide of insects we read : "Bembex rostrata", whereas in the Thieme guide of insects we read : "Bembix rostrata"].
See next Figures.



In its burrow, prepared in the sand, the bembex first delivers not more than one small fly of this or that species, onto which it lays an egg. In the course of two or three days while the larva hatches and feeds on this prey, the bembex does not go down to the nest. Then it provides the larva with a second, this time larger, fly, and again waits, until a third one is needed. In the course of almost two weeks, during which the provisioning of the cell continues, the bringing in of prey follows up one after the other, and the more rapidly, the larger the larva becomes. At the end of this period we can see the bembex every now and then return to the nest with a fly, and again flying out for hunting, bringing-in into one cell a total of 60 flies of medium size or up to two dozen large horse-flies (Tabanidae).
In connection with this, the short indication of simultaneously provisioning three nests in Sphex albisectus Lep., based on an accidental and not further clarified finding, is doubtful (FERTON, 1902). [That is, it might have occurred within the species as an individual exception, but cannot form the basis of an assumption that holds this behavior to be the rule in this species.]
There are still features of the habits of Bembex that deserve attention. First of all the non-correspondence between, on the one hand, the complex slowed-down method of provisioning the larvae (and even with prey consisting of diptera!), and the seemingly simple thing of nursing only one single larva in the bembex nest, on the other. In order to understand this non-correspondence we must take into account that some exotic species of Bembex are known to construct branching nests wit a number of cells.
Further, B. rostrata, as well as some other species of Bembex, next to the cell, in the same nest, digs out yet a special "side burrow", completely looking like a side channel. To this also must be reckoned the complex work of B. rostrata inside the nest, connected with the repeated and diverse displacement of small lumps of soil wholly looking like the work of digger bees (such as Colletes or Systropha). All this provides a basis for seeing the nests of B. rostrata as symplified nests with a reduced number of cells, reduced, that is, to one. It is even very probable that the complexification (the slowing-down) in provisioning of the cells with prey in the Bembex's also was the cause of symplifying its nests to unicellular ones [Having now its provisioning become progressive, the certainty of the larva becoming mature is greater, and therefore less cells are needed : Improved care for the brood ==> decreased fertility. In it, the effective fertility remains the same, or even increases].
Not long ago, in southern Texas (USA) and in the south-west of New Mexico (USA) colonies were found of the rare species Bembex u-scripta Parker. In this species, in contrast to the other bembexes, the ocelli [single eyes on top of the head] are well developed corresponding to its twilight way of life. The smoothing-out of the nest heap, the female of B. u-scripta carries out in a peculiar way. As a result, the entrance opening of its burrow, covered up with sand, remains at the side of the nest hill. The burrow is deepened in the soil under an angle of 45-70 degrees. At a depth of 10-22 cm the burrow ends in an almost horizontal cavity -- the cell. The prey of this bembex consists of totally different flies, being very small as compared to the wasp itself. In addition, B. u-scripta does not grab its prey in flight, but takes it in its resting condition -- from leaves and branches. The egg is deposited onto the first brought-in fly. It is sticked with one end at the base of the bent-out wing. The rest of the provisions is brought in progressively, from day to day, proportionally to the growth of the larva. When the provisioning of the cell is finished, the side burrow leading to it from the main channel ("side channel") is firmly closed by dumping sand. In this way, running [going out] from the same end of the main channel, also a second cell is constructed, and after that a third, and sometimes a 4th and 5th, which [all] is not usual in bembexes.
According to the substantial observations of TSUNEKI, 1958, usually one cell is made in the burrows of the Japanese bembex B. niponica Smith. But in some of its colonies usually nests are encountered with a number of cells. The prey of B. niponica consists of various diptera, represented by not less than 55 species. The method of provisioning the investigator categorizes as "progressive-mass provisioning", whereby the number of preys being brought into one cell varies from 10 to 20 (maximally 35). The life of such preys continues for 1-9 days. From the paralyzed, and having visited flowers, flies, the bembex sucks out the sweet content of their croplets. After the bringing-in of the first prey the wasp finishes the digging work on the cell, but sometimes it is observed that she cleans up the cells, removing from them the remains of not consumed prey. The first prey usually is small, dead or almost dead. It is remarkable that it does not serve as food for the larva, but is largely used only as a postament (base) for the egg that is being laid by the wasp.
For a special study of the behavior of the wasp inside the nest, glass cells and tubes were taken, some simple, some branched. It turned out that the wasp, having returned to the cell, touches the larva with its antennae, holding it between them. When the larva lay too close to the entrance to the cell, the wasp took it with its jaws and dragged it deeper toward the middle of the cell. When, on the other hand, the larva lay too far from the prey, the wasp dragged one or two flies to the larva, taking them from the depot at the entrance of the cell. To this we must add that some exotic Bembex-species collect other preys than flies, namely damselflies (Agrionidae), butterflies (Lycaenidae), and orthoptera (Locustidae).
Of much interest is the method of oviposition in bembexes. Today we can say that precisely such habits lay at the base of the development that has led later to a complete bloom of vespine life. Still in the "paniscoid" (orthoparasitic) Phase in the terebrants, we saw how the elongated egg, while it came out of the oviduct with its posterior end in front, was, with precisely this end firmly attached to the skin of the caterpillar, having its anterior end free. This relationship was essentially preserved in scoliid waps with the difference that the egg here is simply sticked with the posterior end to the body of the prey. And in this case the larva, after having emerged from the egg, only needs to bend itself in order to begin feeding on the delicate prey beneath it.
These relationships changed little in those wasps (Psammocharidae, partly Larridae, and others) in which the egg, going out, orientated as before, from the sexual channels of the female -- with the posterior end in front, began to be attached to the body of the prey in a lying position, on all, or almost all, of its length. But later, in order to guarantee, evidently, a better penetration of the newly-born larva through the robust integument of the new preys, the egg began to be attached the other way around, namely with the head-end toward the body of the prey. See next Figure.

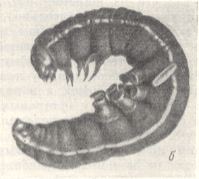
Figure 7 : A caterpillar of a night-owl, paralyzed by an Ammophila lies at the bottom of the cell (top image). Bottom image : The same, taken out of the cell.
(After MALYSHEV, 1966)
This new way of attaching the egg, guaranteeing the immediate penetration of the emerging larva into the proper place of the prey's body, of course demanded special and complex methods of the mother-wasp. With their help the wasp, formerly letting descend the egg with its posterior end in front, first lets the egg be free above the prey's body and then attaches it with the head-end toward the place where the larva will start to feed, as BAERENDS, 1941, showed for Ammophila campestris. In this way behave, evidently, also many other digger wasps (Sphex, Sceliphron, Tachysphex, Notogonia, Oxybelus, and others), and also Scolia hirta Schr., according to FABRE, and S. manilae Ashm., according to WILLIAMS, 1919.

Figure 8 : A paralyzed tachinid fly (Dexiomorpha licta M.) -- one of the 4 preys taken from the cell of Bembex megerlei Dahlb. The egg of the wasp is attached with the posterior end toward the wing-base such that the emerging larva may reach the throat of the fly.
(After MALYSHEV, 1966)
But here this operation is carried out with special precautions. For oviposition, a small fly with a delicate integument is selected. It is placed on its back deeply in the cell. One wing of it is bent out to the side, and the middle leg of that same side is stretched out such that the dislocated wing is held at its place. The egg is sticked with its posterior pole at the base of the dislocated wing, beyond the place of wing attachment, and the egg's head-end projects upwards above the throat of the fly, where indeed the larva will start feeding. One can hardly think that this method [this way of doing things], complexified by supplementary operations, but nevertheless still being essentially original, of oviposition, was derived from those secondary ways of oviposition (with the head-end toward the prey) which were indicated above. For us there is not any basis to assume a reversed process, [namely] to the starting point. The original way of oviposition is also known in Stizidae (for example, Stizus fasciatus F., according to FERTON, 1901). This apparently strange way of oviposition by the bembex-like wasps not only shows their relationship with the higher, pleated-winged, wasps, indicated by FERTON, but, consequently, also with such primitive wasps as are the scoliids forming with the higher wasps one group, the Vespoidea.
Remarkable relationships, [encountered] in the present Phase of evolution, originated between the "bee-wolf" Philanthus apivorus Latr. [and related species] and its prey, [this prey] being a representative of the higher hymenoptera, the honeybee. See next Figures.
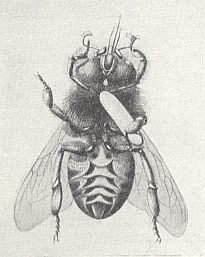
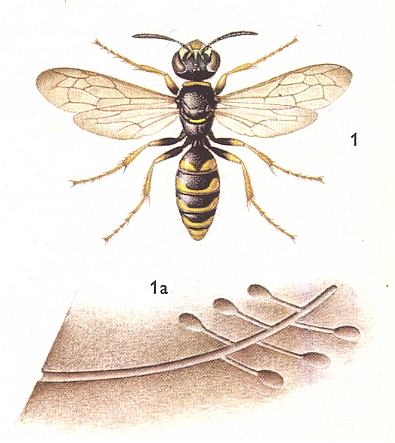
Figure 10 : Philanthus triangulum, 12-18 mm. Sphecidae. 1a - burrow (length 20-100 cm). (After SEVERA, in ZAHRADNIK, Thieme's insektengids voor West- en Midden-Europa, 1977)

In these relationships not any adaptation to the stuggle with its specific enemy has appeared in the bee, but the hunting instincts of the wasp have reached a high level of perfection. The wasp catches its prey on flowers where the bee collects nectar, and with one sting into the throat paralyzes it. Then a peculiar chewing and compressing of the bee takes place, but without injuring its body. As a result, nectar, collected by the bee, oozes out of its mouth, and the wasp licks it off. The prey, now freed from nectar, is offered to the carnivorous larva to which the nectar is said to be lethal.
Here we have to do, apparently, not with [possible] poisening of the larva by nectar through the gut, but with the nectar's contact-action on it (clogging of the stigmata, and similar things).
In artificial conditions a philanth [the bee-wolf] (Philanthus) may kill in this way the prey as if exclusively for the wasp's own feeding, but, we must assume, in nature the wasp does it by the way, when provisioning its cells. Generally the philanth, as also other nectar-consuming wasps, eagerly visits flowers accessible to it.
A substantial study of the behavior of the "bee-wolf" at the moment of paralyzation by it of the honeybee was carried out by RATHMAYER, 1962. For this study a female of the philanth was placed, together with a worker bee, in a wire cage (100x65x40 cm). The philant quickly threw itself onto the prey. After this, one could take with pincers the bee together with the wasp having grabbed it, and easily observe the operation from different angles and at different moments. It turned out that the philant attacks the bee frontally and grabs it at its thorax from above. After having bent its abdomen under the bee, the wasp stings it at the small membraneous region behind the anterior pair of the prey's legs. At the moment of attack the bee attempts to sting the philanth, but in the present position its sting is oriented only at the for it impenetrable chitinous integument of the wasp's abdomen. Usually the sting of the philanth is not applied to the anterior thoracic ganglion itself of the bee, but to its close surroundings. In order to precisely direct the sting to such a small region on the thorax of the prey, the philanth possesses, as does the majority of wasps, special sensorial bristles at the end of the sheath of the sting. After the stinging at the proper place the fore- and middle-legs of the bee are immediately paralyzed.
After [on the other hand] experimentally stinging into the middle thoracic region, the bee is still able to move for 30 seconds. In stinging into the abdomen paralyzation sets in after 220 seconds, but some bees do even recover.
After stinging, the philanth turns the bee on its back [i.e. with its ventral side toward it], and then, having bowed the end of its abdomen to the bee, strongly kicks it forward. As a result of this, the content of the throat of the bee starts to come out of its mouth in droplets which the philanth licks off. Further, we see a third act : massage with the snout and jaws of the wasp of the place of the sting -- the base of the bee's fore-legs, but without inflicting any injury [wounds]. In this way the diffusion of the venom through the haemolymph in the body of the prey is accelerated.
Experiments with artificial paralyzation showed that the venom of the philanth can be effective also independently of the place of stinging. For this it is sufficient when it reaches the haemolymph. The closer it is led to the muscles of the limbs of the prey the faster their paralyzation sets in. The activity of the heart is not directly dependent on the venom. In addition, the venom of the philanth is not rigidly specific to bees : It is effective in insects of all investigated Orders and even also in spiders. But in the haemolymph of the philanth there is a component that neutralizes its own venom, provided the latter is not applied directly into the ganglion. Immune to the venom of the philanth turned out to be only the wasp Palarus variepatus F., hunting for various hymenoptera including philanths.
There is an indication that other philanths, hunting for solitary bees (of the genus Halictus), go their business in a more simple way. Thus, Philanthus punctatus Say., according to PECKHAM, 1905, after having hit the Halictus with its sting into the throat, brings it to its nest without further processing it. This difference of behavior should be expected. After all, the halicti collect to their cells almost exclusively pollen of which they form a fairly firm bun, hardly moistened by hectar (or by saliva). In this way nothing is sqeezed out of the bee's throat, and therefore the [above mentioned] operation is not worked out.
In the moderate zone of North America the philanthous wasps (Philanthinae) are represented by yet another genus, namely Aphilanthops with three subgenera, containing all in all about 15 species. According to existing data, all of them hunt for ants. Such a prey is so exclusive and peculiar that they, as contrasting with the bee-wolf, might be called "ant-jackals". Their hunting behavior, especially with respect to transporting the prey, by some of them is unusual. One species, best known biologically, of the subgenus Aphilanthops (s. str.) -- A. frigidus Sm., carries its prey still in -- we might say -- a rather simple way. Having grabbed the paralyzed ant at its antennae with its jaws, the wasp holds the ant's thorax with its hind-legs, with its middle-legs holding the thorax only from above, and, finally, with its fore-legs lightly holding the ant at its head and prothorax. In this way, in this species, the wasp's abdomen takes no part whatsoever in the transporation of the prey. It is also remarkable that this wasp attacks only female ants of the genus Formica, that still haven't shed their wings. Only when the prey has been brought to the nest the wasp detaches its wings. In one cell it places two, or more often, three ants, onto one of them it oviposits.
The representatives of the other subgenera -- Clypeadon and Listropygia -- hunt for another sort of prey, namely for the wingless worker-ants of the genus Pogonomyrmex. Substantial data on all this are obtained by EVANS, 1962, regarding Aphilanthops (Clypeadon) haigi Bohart. See next Figure.
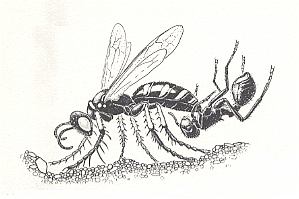
Figure 12 : A female Clypeadon laticinctus digging into her nest entrance while holding a worker harvester ant on the end of her abdomen. (After EVANS and EBERHARD, 1973)
The Clypeadon attacts ants and paralizes them at the entrance of the ant-nest or inside it. In transporting the ant to the wasp's nest, the wasp holds it by means of a special clamp -- an "antclamp" -- at its last abdominal segment. The upper and lower half of this segment (pygidium and hypopygium) carry various projections precisely adapted to clasp the basal segments (coxae) of the middle- and hind-legs of the ant. The effective action of this mechanism is realized, apparently, by blood pressure and the contraction of corresponding muscles. Thanks to such a unique adaptation, the mouthparts of the wasp and all three pairs of its legs are totally exempt from the business of transporting and holding the prey. A similar adaptation of the last segment is observed also in the only known representative of the subgenus Listropygia. And here it is expressed still more perfectly.
With all this we conclude our exposition of the Fourth Vespine (bembecoid) Phase of hymenopterous evolution.
Noëtics
In this section we will partly recapitulate, and develop still further, our noëtic interpretation of organic evolution (It's not an easy text, it depends on theoretical results obtained in earlier documents, and repetitions -- for the sake of clarity -- could not be avoided). And it is in this theoretical -- metaphysical -- context that we should see the evolution of Hymenoptera, and indeed of all organisms.
Let us first determine some important concepts of this interpretation.
First of all we have Reality consisting of two Orders, viz., the Implicate or Noëtic Order and the Explicate or Material Order. Entities in the Implicate Order are immaterial (noëtic) patterns. These patterns can engage in "noëtic reactions and derivations" (where no exhaustion of reactants can take place). Under certain circumstances these originally noëtic patterns, reactions and derivations can be "projected" into the Explicate Order. They then unfold along space and time dimensions an appear as material individuals or material individual processes in different localities in the material order (in the Implicate Order nothing is material, individual, or local). These originally noëtic patterns are, under certain circumstances, not only projected up into the Explicate Order, but are also injected again (without loss of the projected) down into the Implicate Order. In fact there is a constant alternation of projection and injection of such a pattern. When this, for some reason stops, the pattern will disappear from the Explicate Order (but still being present in the Implicate Order).
The central concept in all this is that of Substance in the metaphysical (amended) Aristotelian sense. We here concentrate on organic Substances (inorganic substances are, for instance crystals).
An individual organism is an embodiment of its biological species (and then also of its genus, family, etc.) and exists in the Explicate Order. It, together with all other individuals of that species, is the result of projection, from the Implicate Order into the Explicate Order, of an initially noëtic (= immaterial) pattern representing the (common) Form (in the metaphysical sense) of the individual Substances as they exist in the Explicate Order. Such a Form can thus represent a species, and higher-level Forms can represent genera, families, etc. And on all levels of generality such a Form is itself a Strategy, that is, containing in itself, in fact b e i n g itself, a prescription (or description) of how to exist in the Explicate Order.
In the Implicate as well as in the Explicate Order we have to do with stability of a Form (Strategy). In the Implicate Order it is noëtic stability, in the Explicate Order it is material stability. When we speak of extrinsic stability (noëtic or material) it refers to a Form's compatibility with (noëtically ot materially) external factors. When we speak of intrinsic stability (noëtic or material) we mean the internal coherence of the Form insofar this coherence is not dependent on (noëtically or materially) external factors. Closely connected with, in fact a cause of, stability of a Form (Strategy) is its integration. When we speak of extrinsic integration (noëtic or material) we mean the integration of the Form (Strategy) in its (noëtical or material) surroundings (that is, forming one whole with these surroundings). When, on the other hand, we speak of intrinsic integration (noëtic or material) of a Form (Strategy) we mean the integration of every part of such a Form in that part's surroundings which here constitute the whole Form itself. In the Explicate Order a higher degree of intrinsic integration of a Form (Strategy) is seen as "higher organisms". And indeed evolution leads to higher and higher organisms (anagenesis), that is, organisms that are in a higher degree intrinsically integrated.
Organic species and higher taxonomic units are seen by us as the result of projections, projections, that is, from the Implicate Order into the Explicate Order, of s t r a t e g i e s which are noëtic patterns (Forms) that are, each for themselves prescriptions of how, for such a pattern, to exist and persist in the Explicate Order. And only when such a pattern indeed exists in the Explicate Order, it is ontologically complete : Upon projection into the Explicate Order the (initially) noëtic pattern becomes material (i.e. it, as a Form, is received in Prime Matter), (becomes) individual, temporal, spatial, and local. And indeed, every noëtic pattern, every pure form, residing in the Implicate Order, strives for ontological completion, which is accomplished by being projected into the Explicate Order. The form [pattern] then becomes a true s u b s t a n c e in the metaphysical (and especially Aristotelian) sense [This ontological evaluation of Substances -- being more truly a Substance when materialized -- is an inverse one when compared to the evaluation as it was done in Greek Philosophy, in which, although to a lesser extent in Aristotle, the pure, immaterial Form is the perfect way for a Substance or for any other pattern to exist. For Aristotle the f o r m is inside the material Substance (i.e. it is immanent, not transcendent as in Plato), and only here and there in his works he hints to possible "separate (from matter) entities"]. Anyway, we consider something to be more truly a Substance when it is materialized, i.e. when it is unfolded along space and time dimensions resulting in it becoming material, individual, temporal, spatial, and local.
Another factor enhancing a pattern's degree of being more truly a Substance, or at least the Form of such a Substance, is its i n t r i n s i c stability. If, in the Implicate Order a given noëtic pattern is in a stronger degree intrinsically stable (as a result of it being more intrinsically integrated) than some other pattern, it is in a correspondingly stronger degree a Form of a true Substance, but indeed, still not more than a mere Form of such a Substance : Only upon projection into the Explicate Order it will be a true Substance, being ontologically complete. And if a given Form (Strategy), to be projected, is in a higher degree intrinsically stable than another such Form (as a result of it being in a higher degree intrinsically integrated) it will, upon projection, result in, not only in an ontologically (= metaphysically) complete Substance, but also in a Substance that is in a higher degree materially intrinsically integrated (than the "other such Form", just referred to).
Because we assume two Orders of Reality, we, basically, have to do with two chief sets of existential conditions. And such conditions directly involve, or are in fact equivalent to, the phenomenon of stability. Let's dwell on stability for a while.
Stability, and thus the ability to exist and persist -- despite perturbations -- of a given thing or pattern -- here an organic thing or pattern, i.e. a strategy -- in the Explicate Order, is extrinsic stability, and it is two-fold : First and basic is thermodynamic stability. It depends on the prevailing thermodynamic conditions in which such a thing or pattern exists. Secondly, there is the ecological stability. It depends on the compatibilty of that organic pattern with the prevailing ecological conditions, its ecological environment. That is, whether the strategy, embodied in its morphology, physiology, and behavior, really is, presently, a true e x i s t e n t i a l condition, a condition, that is, to exist and persist in the Explicate Order.
In the Implicate Order we have to do not only with extrinsic stability but also with intrinsic stability. The extrinsic stability of a noëtic pattern, residing in the Implicate Order, depends on its noëtic surroundings. The latter should, of course, not be understood spatially but purely noëtically. As has been explained in earlier documents (where the evolution of dipterous insects was discussed), the organic noëtic patterns -- which we here call "strategies" -- reside, in the Implicate Order, in their proper noëtic stability fields (not spatially understood). Outside these fields they cannot exist in the Implicate Order :
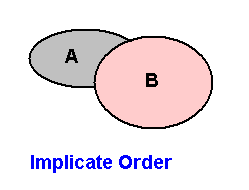
Figure 13 : The noëtic stability fields of two noëtic patterns (strategies) A and B in the Implicate Order. A pattern has only extrinsic noëtic stability when it resides within its proper noëtic stability field.
In addition, we have, in the Implicate Order, to do with intrinsic stability. A given noëtic pattern (strategy, Form) has a greater intrinsic stability than some other pattern if it is in a higher degree intrinsically integrated. For this type of (noëtic) stability we have no proper analogue in the Explicate Order (like we do have it for extrinsic noëtic stability in the form of thermodynamic and ecological stability in the Explicate Order). So we cannot imagine what precisely noëtic intrinsic stability, and therefore "intrinsic integration" is. But its counterpart in the Explicate Order, as a result of projection, is imaginable, at least insofar as it appears in individual organisms : Such an organism is in a higher degree intrinsically integrated than some other organism if its composing parts and functions (including behavior) are less independent, that is, if they depend (as to what they in fact are, and as to their existence) more on the whole organism (the whole strategy) than they do in that other organism. We assume that some kind of intrinsic integration (having its extreme opposite in disintegration) of any given noëtic pattern exists in the Implicate Order, resulting in intrinsic noëtic stability. And it is the one-way transition from lower to higher intrinsic noëtic stability (resulting from higher intrinsic integration), not of the same pattern, but a transition from a less in itself integrated pattern to one that is more so integrated, that is expressed by the noëtic trajectory going from one noëtic pattern (and its corresponding noëtic stability field, representing the pattern's extrinsic noëtic stability) to the other (as explained in the mentioned earlier documents). And it is this noëtic trajectory, irreversibly going -- in the Implicate Order -- from a strategy with a lower degree of intrinsic stability to one with a higher degree of intrinsic stability, that, upon projection of these strategies into the Explicate Order, determines the organic evolution. There, in the Explicate Order, we see the course from lower to higher intrinsic stability (as it takes place in the Implicate Order) as an increase in morphological, physiological, and behavioral intrinsic integration of the parts of the relevant organisms, which increased integration is often called "higher-development" (as we see it, for example, in vertebrate evolution starting from fishes and culminating in mammals).
The main direction in organic evolution in the Explicate Order is driven by the derivation -- in the Implicate Order -- of one noëtic pattern (strategy) from another, expressed by the course of the noëtic trajectory in that Order. This course is a derivation from noëtically simplex to complex, which in turn is from less intrinsically integrated to more intrinsically integrated, which again in turn is a derivation from less an intrinsic noëtic stability to increased intrinsic noëtic stability. This course can be more or less retrieved by Phylogenetic Systematics (as explained in earlier documents on the evolution of Diptera).
Upon projections of these noëtically successive patterns (strategies) into the Explicate Order we see the main course of material and temporal organic evolution expressed as "higher-development" (anagenesis), that is, the evolution leading from lower, primitive organisms -- existential strategies -- to higher, advanced organisms -- also existential strategies.
As has been explained earlier, a "strategy" is, first of all, a noëtic pattern, residing (with many other such patterns) in the Implicate Order. It is a set of coordinated and each other supplementing potential existential conditions -- conditions, that is, making it possible for the pattern to exist in the Explicate Order and so becoming ontologically complete. So a strategy, representing a species or type of organism, if it is to work in the Explicate Order at all, it must be integrated in the existing ecological context. Or, said differently, the strategy must be compatible with the prevailing ecological conditions. We will not dwell on this type of integration any further, because here the matter is clear enough. The strategy, still as it appears in the Explicate Order, is not only -- as described -- extrinsically integrated, but also -- in a certain degree -- intrinsically integrated, as can be visible in the organism's morphology (integration and specialization of body segments in arthropods, for example), physiology (dependency of organs on each other), and behavior (integration of the several instinctive actions connected with provisioning in wasps, for example). In this integration it is the p a r t s of the (given) strategy that have lost their independency. They are now in a certain degree dependent upon each other, and in fact dependent upon the strategy (= the whole organism, as a species or type, morphologically, physiologically, and behaviorally) as a whole. This intrinsic integration can be of different degrees in different strategies, and its noëtic counterpart is the intrinsic noëtic integration of strategies (each for themselves) as they reside in the Implicate Order as noëtic patterns. And here, in the Implicate Order, the greater the intrinsic noëtic integration of a given strategy, the greater its intrinsic noëtic stability. So, as has been said, in the Implicate Order the noëtic trajectory will pass from strategies having lesser intrinsic noëtic stability to those having greater stability.
But a strategy, in a higher degree intrinsically integrated than some other strategy, is, in the Explicate Order not necessarily 'better', in the sense of ecologically and physiologically more stable, meaning that it has not automatically a greater capability to exist and persist in the Explicate Order. When the prevailing ecological conditions change, the species (or even a complete higher taxon) may become extinct : The (particular) strategy will then not be projected (from the Implicate order into the Explicate Order) anymore, because the strategy is now (at the time of the mentioned ecological changes) not a set of true existential conditions anymore. And when, on the other hand, in the Implicate Order, the noëtic trajectory passes over from a pattern (a strategy) having less intrinsic noëtic stability to one with a greater intrinsic stability, this does not necessarily mean that this latter strategy will be projected into te Explicate Order, because it will only be so projected at a given point in time if the strategy, as given in a noëtic form, is fully compatible with the ecological conditions prevailing at that point in time in the Explicate Order.
So the succession, and abundancy (representing ecological success) of species and higher taxa of organisms in the Explicate Order is for an important part determined by the prevailing ecological conditions. So in the Explicate Order -- and that is in evolution -- species and groups can appear and later become extinct.
On the other hand, in the Implicate Order the (noëtic!) succession (consecutiveness) of strategies (noëtically representing species or higher taxa) is purely formal : It is a successive formal noëtic d e r i v a t i o n of each time one strategy from another, based on intrinsic noëtic stability, and expressed by the noëtic trajectory. And because the course of this derivational path is driven by i n c r e a s i n g intrinsic noëtic stability, it is a one-way course (the path - the noëtic trajectory -- will not lead from greater stability to lesser), the explicate counterpart of which we see in the irreversibility of organic evolution : Dollo's Law.
Let us give a more concrete -- but still general -- example of all this (Later, when having dealt with the evolutionary data of all Hymenoptera, we will give truly concrete examples).
Say we have two related recent families of, say, hymenopterous insects, that is, families, of which representatives are still living today. Let us call them A and B. Suppose further that family B, judging from recently living representatives, is definitely more primitive ('lower') than family A, meaning here that the strategy "B" is in a lesser degree intrinsically integrated than is the strategy "A", or, said differently, A represents a 'higher' form than B (but nevertheless still related to it). We will see that, in the E x p l i c a t e Order, being a 'higher form' does not necessarily mean that it will appear later in evolution (although it most often does [i.e. the rule still holds that forms that are 'higher' will appear later in evolution]. In the I m p l i c a t e Order, on the other hand, it indeed is noëtically derived 'later'. But here "later" is, of course, not meant temporally but noëtically : 'later' here in fact means "noëtically derivable" (or, equivalently, "noëtically derived").
Suppose further that indeed there are sufficient indications that (the more primitive) family B appeared a little later in evolution than did (the more advanced) family A (based, for example, on the absence of fossils of representatives of the family B, and the presence of fossils of representatives of family A). So in evolution family B appears after family A. But, family B cannot be derived (morphologically, ethologally, etc.) from family A, because then we would have to assume a reversion of evolution, that is, an evolution from advanced to primitive, and thus violating Dollo's Law which is fairly well established in paleontology. Situations like the one described, often occur in the study of organic evolution, and are not easy to deal with. However, our noëtic theory of evolution is able to throw more light on such questions.
Let us, then, depict the mentioned situation in terms of the Implicate and Explicate Orders, where the noëtic counterpart of a family (of organisms) is (the noëtic description of) a particular s t r a t e g y for it to exist (in the form of individual organisms) in the Explicate Order if allowed by the prevailing ecological conditions. So here a "strategy" means a noëtic pattern, residing in the Implicate Order in its proper field of extrinsic noëtic stability. There are many noëtic patterns in the Implicate Order. Some are very complex and will never come to exist in the Explicate Order, because they are incompatible with whatever condition in that Order, but many of them simply are strategies, and they may at some time appear in the Explicate Order. And they will be there in the form of organisms. Such a strategy may represent a species or higher taxon of organisms existing in the Explicate Order (In the present example it represents a family of organisms). The execution of a given strategy is visible in the Explicate Order in the form of special morphological, physiological, and behavioral adaptations present in the individuals of the taxon. So a species or a higher taxon of organism is in fact at the same time a strategy for it to exist in the Explicate Order :
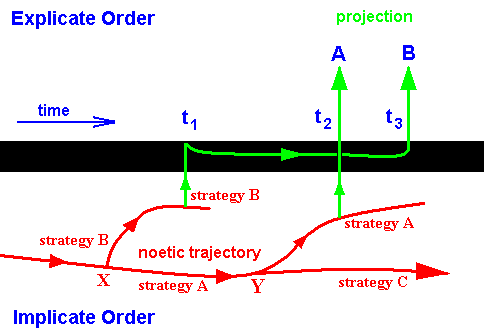
Figure 14 : Implicate and Explicate Order (Symbolically separated from each other by a thick black line). When we speak of p r o j e c t i o n of some pattern (strategy) (from the Implicate to the Explicate Order) "at a certain point, say, t1, in time", this is (merely) seen so from (and happening in) the Explicate Order, because in the Implicate Order there is no time dimension.
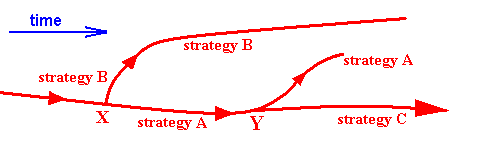
In the Implicate Order we see (part of) the noëtic trajectory (red) visiting extrinsic noëtic stability fields (not drawn) containing strategies (not drawn). The consecutive order of this visiting (derivation) is determined by increasing degree of the intrinsic noëtic stability of the strategies visited. Two strategies are emphasized here, viz. that representing the family A, and that representing the family B. At X the noëtic trajectory splits up into two branches, that, upon p r o j e c t i o n give rise to representatives of the family A and family B in the Explicate Order.
At time t1 the (in itself unchangable) strategy of family B is not yet compatible with the ecological conditions prevailing at that time (for example the strategy demands as prey caterpillars of a certain species which do not yet exist at time t1). So this strategy cannot, and will not, at that time, be projected into the Explicate Order, meaning that at that time this family has not yet appeared in evolution.
At time t2 the strategy A can and will be projected, resulting in the evolutionary appearance of representatives of the family A. As long as this projection continues (in the context of continuous projection and injection of the strategy [which itself is a definite pattern] ) the family will continue to exist in the Explicate Order (and this projection will continue as long as the strategy remains fully compatible with the prevailing ecological conditions).
At time t3 the prevailing ecological conditions are such that now also strategy B can be projected (so the first projection of this strategy came after the first projection of strategy A). Morphological, physiological, and behavioral features of the representatives of family B, although it appeared somewhat later in evolution, showed that family B is more primitive, less intrinsically integrated, and thus 'lower' than family A. So Phylogenetic Systematics in evaluating these features, lets family B branch off from the main stem e a r l i e r than family A. But in our (noëtic) interpretation "earlier" cannot be temporally meant, but only formally so, that is, the strategy of family A is, as a noëtic pattern in the Implicate Order, derived from the strategy of family B, and not vice versa (and only in this sense A is 'later' than B). So the resulting 'genealogical tree' of Phylogenetic Systematics is in fact not a genealogical tree (which implies time) but a noëtic tree 'drawn' by the noëtic trajectory, which indeed ' first' visits strategy B (or some strategy very related to it) and ' then' strategy A (i.e. A is noëtically derived from B, because A has greater intrinsic noëtic stability than B has). So the seeming discrepancy between the time of appearance in evolution of these strategies and the degree of their being primitive or advanced is solved.
The above Figure thus depicts the noëtic interpretation of certain biological, morphological, and paleontological facts. The current phylogenetic interpretation (thus without assuming an implicate order) of these same facts would be :
A more or less similar example might be a case where fossils are found indicating that at time t1 strategy (say) E already existed in the Explicate Order. But this same strategy seems to be still existing at time t3 (which might be signifying present time). The noëtic interpretation of these facts might be as follows :
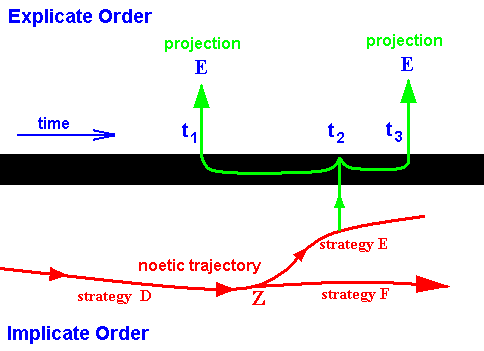
Figure 15 : Implicate and Explicate Order (Symbolically separated from each other by a thick black line). The noëtic trajectory at Z branches off into strategy E (and also into F). Only at (and around) time t1, and only since time t3 (present), the ecological conditions in the Explicate Order are such that projection of strategy E is possible (and will therefore take place). During the long period between t1 and t3 the ecological conditions were incompatible with this strategy.
Current phylogenetic systematics would interpret these facts differently : Strategy E evolved around time t1 and, independently, again around time t3. It would say that we here have a case of convergent evolution :
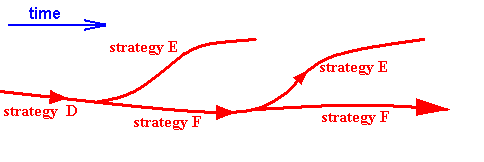
Figure 15a : Current phylogenetic interpretation of the facts connected with the situation as depicted in Figure 15. From strategy D strategy E evolved, while evolution proceeded further through strategy F (also having evolved from strategy D). Then, in addition to continuing with strategy F, evolution (also), from strategy F bends back to strategy E again : convergent evolution.
In the above examples we have indicated the role played by ecology in the appearance of different (or the same) strategies. We should, however, not exaggerate this role. Generally, the order of formal noëtic derivation of one strategy from another, that is the course from strategies being intrinsically less noëtically stable to those that are more stable in this respect, drive evolution throug consecutive projections. But of course, say, the particular strategy that we collectively may call "bees" can only project into the Explicate Order when flower-plants (angiospermae) exist there (i.e. when this particular plant-strategy was already projected earlier, and keeps on being projected).
Let these examples for the moment suffice in making clear our general ideas of organic evolution.
By their, throughout formal, and by intrinsic stability determined, derivability, the noëtic patterns in the Implicate Order are, in a branched way (tree-like) connected with each other, expressed by the noëtic trajectory. This trajectory is, consequently, like everything else in the Implicate Order, in its nature not dynamic but static, or, perhaps better, logico-noëtical, and it places the noëtic patterns in a definite consecutive order, i.e. it determines this order. And, as has been said, this static noëtic order of formal derivability in the Implicate Order drives, through projections, organic evolution in the Explicate Order insofar its 'higher-development' (anagenesis) is concerned, that is, evolution from primitive, still little intrinsically-integrated forms (strategies), to more and more highly intrinsically integrated forms (strategies). The fact that the noëtic trajectory is not simply linear (not just one simple line) is caused by the fact that a given less intrinsically stable pattern might have several, mostly two, different alternatives, both, albeit in a different way, in a higher degree intrinsically stable.
So our theoretical concept of "noëtic trajectory" is certainly not a mere ad hoc construction, but an integral part of our noëtic theory of evolution in terms of the Implicate and Explicate Orders.
We have just said that ecology in the Explicate Order only plays a limited part in the consecutivity of appearance of species and groups of organisms in this Order. The greatest part is played by the noëtic consecutivity of strategies in the Implicate Order. The next Figure shows this :
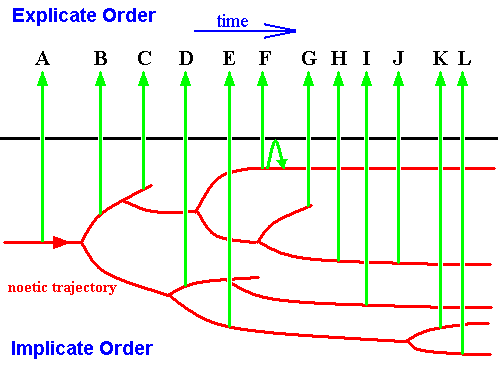
Figure 16 : Implicate and Explicate Orders. In the Implicate Order we see the noëtic trajectory visiting many strategies (as noëtic patterns [not drawn] residing in their proper noëtic stability fields [not drawn] ). Upon the noëtic trajectory visiting them, the strategies are projected (green arrows) into the Explicate Order. Not all projections are shown, only those of strategies A, B, C, D, E, F, G, H, I, J, K, and L.
It might be useful to recapitulate WHAT a Strategy, as noëtic pattern residing in the Implicate Order, actually is.
Each projection is followed by an injection (without loss of projected pattern), then by a projection again, etc., constituting a continuing alternation of projection and injection. Only when the strategy is not compatible anymore with the (changed) ecological conditions this projection-injection will cease. This is the case with strategy F.
As can be seen in the Figure, some branches of the noëtic trajectory terminate, meaning that no further derivation is possible. This is seen in the Explicate Order as an evolutionary dead end.
The Implicate Order is a domain of immaterial entities, patterns. We call them noëtic patterns. As a result of noëtic reactions (comparable to mathematical or logical formal derivations) all possible patterns will eventually exist in the Implicate Order. "Eventually" here is not temporally meant, so all these possible patterns just exist in the Implicate Order, provided they really are patterns, meaning that each one of them is intrinsically non-contradictory and is qualitatively (as to what it is) separated from other such patterns. Because they are only Forms, that is, forms without Matter, they are ontologically incomplete. Therefore they intrinsically strive for ontological completion. They can achieve this when projected from the Implicate Order to the Explicate Order. They then will be unfolded along the space and time dimensions of this Order, or equivalently, they are received by Prime Matter (which is now 'informed' by them), and so become material, individual, local, and temporal. So by having existence in the Explicate Order they obtain ontological completeness. Of course they also still exist (immaterially) in the Implicate Order, because here there does not exist something like exhaustion (of forms).
However, many, if not, most noëtic patterns cannot exist in the Explicate Order, i.e. cannot be materialized, because they are not compatible with its conditions. But many simple ones can be projected into the Explicate Order and are visible there as inorganic patterns (such as crystals). And only a relative small number of complex noëtic patterns are in principle able to obtain existence in the Explicate Order. For this they must carry their own conditions of existence (strategy) in the Explicate Order. And they not only must carry a strategy-to-exist-in-the-Explicate-Order, -- each one of them must BE such a strategy. Only then they are relevant to the Explicate Order. What is the general nature of such a strategy residing as a noëtic pattern in the Implicate Order? The answer is that such a stategy is a description. Not a description done by someone or something, but just a description. And what does it describe? It describes in what (general) way the descripion itself can exist in the Explicate Order. The description, as a noëtic pattern residing in the Implicate Order, is a description of how that very description can exist in the Explicate Order. Such a description is, as description, a true inhabitant of the Implicate Order : It is immaterial, not individual, not temporal, not spatial. And when indeed such a description, that is, such a strategy, is projected into the Explicate Order, we, as observers, see it there in the form of a number of individual living organisms of some particular species. Every individual of such a species embodies the strategy, -- in fact, IS the materialized strategy or description. So when complex noëtic patterns project into the Explicate Order they will appear there in the form of individual organisms.
The whole enormous collection of strategies, descriptions, in the Implicate Order, has an overall structure, meaning that the different strategies are connected to each other in some way. Indeed, they are connected with each other by their noëtic, formal, derivability from each other : in a higher degree intrinsically integrated strategies are derivable from lower ones, because the former are in a higher degree intrinsically stable. So the formal derivation of strategies goes from less intrinsically stable ones to more intrinsically stable ones. And they must also be extrinsically stable : they are only extrinsically stable when they reside in their proper noëtic environment, that is, when they reside in their proper noëtic stability field in the Implicate Order. So the formal noëtic derivation (derivability) goes from one stability field to another and thus from one strategy to another and is expressed by the noëtic trajectory (or several of such trajectories). So this trajectory (which, of course, is not dynamic, but formal), or set of trajectories, expresses the derivational structure of the whole collection of strategies in the Implicate Order. And this noëtic trajectory (or set of trajectories) is, upon projection of the relevant strategies into the Explicate Order, seen there as the evolution from lower to higher organisms.
With all this we conclude this step of further working out our noëtic theory of evolution. In later documents we will proceed still further with it.
In the next document we will expound the Fifth Vespine (moneduloid) Phase of hymenopterous evolution.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LII.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX