
Before us now stands only one group of Hymenoptera -- the so-called Trigonalidae -- of which the systematic position is disputed, and having its evolution totally unstudied. See next Figures.
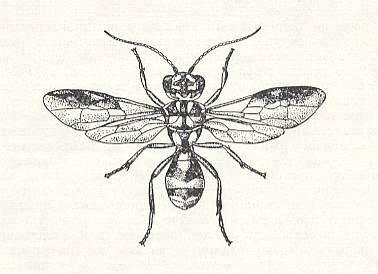
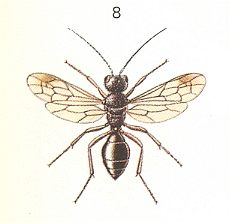
With their about 70 species, making up 17 genera and 5 subfamilies, the Trigonalidae are distributed over all inhabitable parts of the world. In Europe there can be encounterd only one species, Pseudogonalos hahni Spin. (Figure above). All of them are very rare, sporadic, and, as is supposed, are on their way to extinction. According to data collected by Wheeler, 1928, the Trigonalidae are rather small insects, partly colored by light spots or bands, and have a very archaic look, showing a mixture of features of the p h y t o p h a g s [Symphyta (saw-flies and allies)], the t e r e b r a n t s [parasitic wasps and allies], and the a c u l e a t e s [sting-bearing Hymenoptera such as the higher ('true') wasps, the bees and the ants]. Their many-segmented antennae do connect them with terebrants, their head with saw-flies [Symphyta], their overall body shape with bethyloid wasps, and the shape of their abdoment with Mutillidae (ant-wasps). In some species (of the subfamilies Bareogonalinae and Nomadiinae) the anterior part of the head is stretched out and spout-like as in certain scorpionflies (Panorpa, Mecoptera). Cerci [paired posterior appendages of the abdomen] present, but the ovipositor is strongly reduced. BÖRNER, 1919, found out that the lower jaws of them are provided with paraglossic appendages that are so primitive as compared with those of the other terebrants that for the Trigonalidae he created a special division of Hymenoptera, the "Archiglossata". Their wing-venation is very primitive, and usually having three closed cubital cells, it is very similar to that of the aculeata. The feet carry well-developed sole-vesicles, a feature that, according to BÖRNER, is absent in terebrants, but often encountered in the phytophags and aculeates. In addition we can mention that the rudimentary ovipositor of the Trigonalidae is placed apically at the very end of the abdomen, like the sting of the aculeates. Their legs have a twofold trochanter as in terebrants. So, the archaic nature of the Trigonalidae is unquestionable, but as to their morphological features they are certainly terebrants, and not phytophags, neither aculeates.

As CLAUSEN, 1931, informs us, direct observations on a series of trigonalids (of the genera Poecilogonalos, Pseudogonalos, and Orthogonalos), done in India and Japan, where they are more common, showed that the females of the Trigonalidae, in natural conditions, lay their eggs directly onto leaves of different plants (clover, euphorbia, bamboo, and others). Having initially walked on the upper surface of the leaf, the female goes to its edge, and bending the end of its abdomen under the underside of the leaf, lays an egg at a distance of 5 mm from the edge. See next Figure.
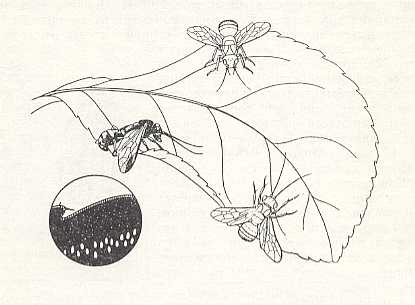
The execution of this operation takes only an instant, and it may be repeated rapidly, although the whole egg-laying period is quite extended : Thus the female of Poecilogonalos thwaitesii Westw. laid during 14 days 10641 eggs, but in another case 4376 eggs in one day.
Clausen, 1931, 1949, tells us about the discovery of one young larva, identical to that of Poecilogonalos maga Ter., inside the pseudo-caterpillar of a saw-fly. There was no mention of any other parasitic larvae inside the latter, as there was also no mention of the fate of the one found. Therefore, his generalization (expressed in a table) that the larvae of Trigonalidae, having ended up in the larva of the saw-fly, also develop at the expense of primary parasites, has no actual basis.
Of the series of typical points discovered in the behavior and development of the Trigonalidae the most significant one is the method of infection of the preys by them, namely their eggs passively ending up in the host through its mouth. Such an exclusive relationship between the terebrant and its preys is, insofar we can judge today, not observed in any other terebrant, and in any of the remaining Hymenoptera for that matter.
Some similar convergent phenomena, known among parasitic worms and flies [tachinids], originated, of course, in their own ways, and therefore, evidently, there are no phylogenetic connections whatsoever to the presently discussed evolutionary process.
There is no doubt that generally archaic terebrants must be archaic not only as to their structure, but also as to their inherited features of behavior or instincts. From this it follows that those clearly archaic terebrants as are the Trigonalidae are expected to express in themselves, in a high degree, fundamental biological features of their original ancestors, closely related to original phytophags. However, about the origin of features of behavior and development, typical only of trigonalids, there are up to now [1966] not expressed any views or explanations, except those proposed by the author (MALYSHEV, 1959, 1960).
In the first [= ancient] conditions of the origin of the terebrants, as we know, each normally laid egg of the archaic inquiline was (if we do not count possible cases of cannibalism) provided with food in the gall. From here it is clear that in the for the trigonalids original conditions their direct ancestors laid a normal number of eggs, whereby the larvae having emerged from those eggs easily fell into appropriate conditions where they developed successfully. (Evolutionarily) subsequently, however, as a result of being too late with laying the eggs into the gall, where as a consequence of this there was an already more or less fully-grown larva of the saw-fly, the developmental conditions for the ancestors of the trigonalids deteriorated [Of course there must have been a cause of the break down of the synchronicity of young galls and egg-laying 'pro-trigonalids', (this break-down being) such that when the eggs are ready to be laid, the relevant galls have become too old.]. Possessing only a short ovipositor, although not so much reduced as it is now, the female of the primitive trigonalid was not able to lay the egg all the way into the cavity of the developing gall [because it had grown already too large] of the saw-fly, but really only into the thickness of its wall or [still] just away from the interior lining of the gall. In this situation some of the few eggs, having ended up, with delay into the already widened cavity of the gall [for these still later laid eggs the wall of the gall had become thinner as a redult of the sawfly's feeding on it, so that the cavity could be reached by the ovipositor], [and having ended up] at the larva of the saw-fly already feeding on the walls of the gall, [these eggs] hardly could have a chance for developing further and giving rise to ectoparasitic larvae. Precisely in these conditions the reverse could happen : The large larva of the saw-fly, gnawing at the wall of the gall, might, together with the green pieces gnawed off by it, also swallow the small eggs of the terebrant.
It is very probable that already in these original conditions a significant number of eggs or young larvae of the original trigonalids perished, compressed by the mouthparts of the host-larva or became lost among the mass of excrements that surround this host in the cavity of the gall. These losses, already in the original conditions, demanded for a corresponding compensation by way of increasing the number of eggs that can be laid by the trigonalid. The young larva, emerging from a survived (i.e. being swallowed without injuries) trigonalid egg, has ended up inside the gut of the sawfly-larva, filled with a green mass gnawed off from the walls of the gall. This essentially was the same nutritive environment at the expense of which also the terebrant-inquiline developed [evolutionarily] earlier. But now the larva of the primitive trigonalid had the possibility to conclude its development not as an inquiline anymore but as a peculiar endoparasite, having ended up inside the prey, not through its integument but through its mouth.
Subseqently, when the chief mass of gall-producing saw-flies became extinct, also the ancestors of the trigonalids disappeared that had laid their eggs into the galls of saw-flies. Of these only those could evolutionarily be preserved that had adapted themselves to parasitism in the new conditions. In this, initially they could infect also larvae of saw-flies, but then those that did not produce galls, but fed out into the open on leaves of trees and shrubs, and accidentally having swallowed together with the green pieces of leaves also eggs of trigonalids laid at the margin of those leaves.
The idea that the evolution of the Trigonalidae went precisely this way is supported by the fact that also in recent conditions trigonalids sometimes develop in ectophytic larvae of saw-flies, and with it in the form of primary endoparasites, as was told above. This is also supported by the very method of infecting the preys, at first shown with respect to a series of trigonalids from India and Japan, but now also shown in the (one) European species. It is very probable that this same passive method of infecting preys is typical also to other trigonalids, having lived up until our days. In the in this way created new conditions the chances for a successful development of trigonalids again significantly decreased. But things became still more complex when instead of by the larva of the saw-fly the [trigonalid] eggs were swallowed by caterpillars [larvae of butterflies] feeding on the same leaves, where the trigonalid eggs were laid. In this case, the trigonalid eggs, having ended up into the caterpillar, did not encounter appropriate biochemical conditions for their development and perished. This, as we now know, did not take place in only two types of cases : (1) when the caterpillar, that had swallowed trigonalid eggs, was infected by an ichneumonid or a tachinid, in which the the development of the trigonalids could successfully continue, or (2) when such caterpillars were apprehended by wasps and being fed by them to their larvae, in which then the development of the trigonalid was completed. So there does not remain any doubt that the precisely very meagre chances for a successful development of the larvae also were the cause that had triggered the unusual fertility of their mother.
So in the evolutionary history of the trigonalids almost from the very origin of them a series of situations followed in which a complete extinction was imminent to them. However, each time when there appeared to be no chances left for their preservation, the Trigonalidae showed a remarkable plasticity and in this way or another escaped from the created precarious situation. As a result they acquired the ability to lay thousands of eggs in a day in conditions in which might develop only a few individuals. And this in a time when such not uncommon relicts, as the evanoid terebrants, the Gasteruptiidae, being inquilines, lay in all their life hardly a dozen eggs. So great the difference turned out to be between these primitive terebrants in their lines of evolution, which went out from a common kernel. Here it is interesting to remember also that remarkable phase of evolution, in which some genera and whole families of terebrants reversed to phytophags, but a phase of an altogether different order, as was told earlier.
Let us, in conclusion, note that the seemingly possible supposition according to which the trigonalids might have been originated directly from ectophytic saw-flies, that is, without going through an inquilinoid phase in galls, meets with a series of unsurmountable difficulties.
First of all, the eggs of saw-flies, in contrast to those of the Trigonalidae, have a simple and delicate, not by anything especially strenghened, shell. Moreover they are relatively very large. See next Figure.
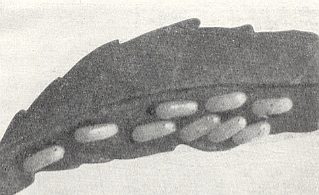
They are laid in relatively small numbers -- not thousands of them as in Trigonalidae, but in fact only tens of them or a little more. Merely an increase of the number of eggs that can be laid, without a provisional adaptation of them with respect to the size and structure of the chorion, could not give a positive result : However many eggs will be laid, no one of them will survive the passage through the chewing mouthparts of the host-larva. Moreover, for such an adaptation to originate, a continuous and regular contact, from generation to generation, is needed, a contact, that is, between more or less fully-grown larvae of one species of saw-fly with the egg-clutches of another [species of saw-fly] which [egg-clutches] these larvae might swallow. In the conditions of life of ectophytic saw-flies such a kind of contact is improbable, which [contact], on the contrary, is completely guaranteed by the conditions of inquilinism. In all this we must see a confirmation of our general thesis according to which all terebrants as a whole originated from gall-producing saw-flies that go through the archaic inquininoid phase. In this the Trigonalidae are no exception.
The Trigonalidae, consequently, just represent a side-branch of terebrants, that directly separated off from the archaic inquilines and developed deviantly, and, as a result of this, their ancient features combined with a series of features of later date. So, with respect to phylogeny there is absolutely no basis in having the trigonalids separated as a special suborder of the Hymenoptera, but also [no basis] in viewing them merely as one of the families of the ichneumonoid terebrants. From the viewpoint of evolution we must see them as an independent superfamily Trigonaloidea, as to its systematic status equivalent to the other superfamilies of the terebrants.
We now see that the basic biological feature of the Trigonalidae is the passive transition of their eggs, a transition connected with a series of fatal contingencies, from the place where they have been laid to the place where they develop. Hence the name of the present stage of evolution -- passively-parasitic, or trigonaloid phase.
With all this we have concluded our exposition of the Passively-parasitic (trigonaloid) Phase of hymenopterous evolution.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XLIV.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII