After having, in the preceding two documents, started and continued with the thematic treatment of the functional wing-types and subtypes of the Suborder Nematocera of the Order Diptera, and their derivation from the order-prototype, - in the present document we continue with those of the rest of the Order (Brachycera-orthorrapha and Brachycera_cyclorrapha), not already treated in earlier documents.
Traction-Lifting many-veined (tabanoid) type
Representatives of the type
Wings, built according to the traction-lifting many-veined (tabanoid) type, are possessed by representatives of various dipterous families, mainly belonging to the suborder Brachycera-Orthorrapha : Rhagionidae, Tabanidae (horse flies), Erinnidae, Coenomyiidae, Solvidae, Therevidae, Apioceratidae, Acanthomeridae, Omphralidae, Nemestrinidae, Mydaidae, Asilidae (robber flies), Bombyliidae (bee flies), and some Cyrtidae. In addition, tabanoid wings are also possessed by certain recent-but-relict and fossil families of Nematocera, viz., Phryneidae (= Rhyphidae), Olbiogastridae, and Protorhyphidae.
Because we haven't systematically and biologically treated all these flies in earlier documents (except for the Rhyphidae, that were so treated together with the rest of the diptera-Nematocera), we will give here Figures of some of the representatives of the tabanoid functional wing-type (All Figures, After CHINERY, in Thieme, Nieuwe Insekten Gids, 1988) :
 Rhyphus [= Phryne = Sylvicola] fenestralis Scop. Rhyphidae [= Phryneidae]. |
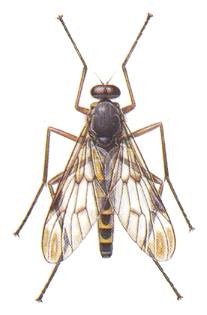 Rhagio scolopacea. Rhagionidae. |
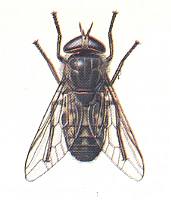 Tabanus bromius. Tabanidae (Horse-flies). |
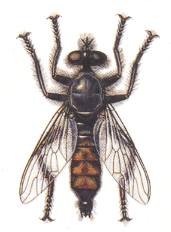 Laphria gilva. Asilidae (Robber-flies). |
For the study of the venation of the wings to come reference should be made to the dipterous venational order-prototype :
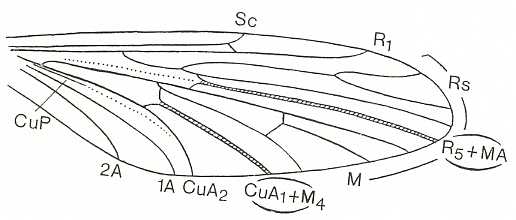
Let us now, then, depict wings belonging to the present functional wing-type, i.e. the tabanoid type.
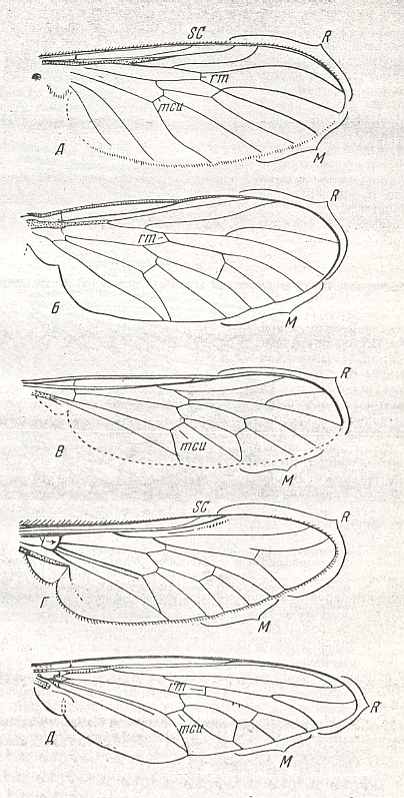
| Figure 1 : Recent traction-lifting many-veined wings of representatives of the phryneoid, ragionoid, erinnoid, tabanoid-proper, and asiloid subtypes. |
Size of the wings
Shape of the wings
Skeleton of the wing
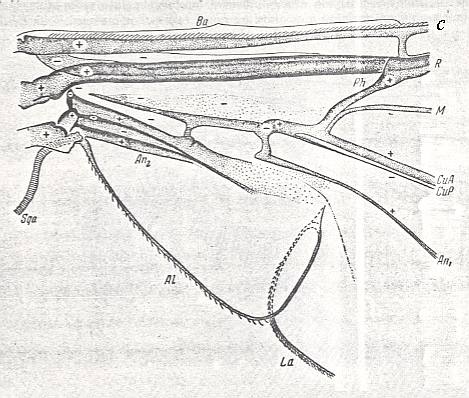
Figure 2 : Nemestrinus capito Loew (Nemestrinidae). Basiala of right wing of a male, from above.
Length : The whole wing 17.0 mm. The basiala 4.5 mm. (For the abbreviations see the aforementioned Figure).
(After ROHDENDORF, 1951)
The phragm is, as a rule, well-expressed as a strong and firm outgrowth, connecting the base of the Radial vein with the lower element of the basiala (the base of Cubital and First Anal vein). Seldom is the phragm in the form of a sclerotized and darkened fold (phryneoid, rhagionoid, and cyrtocioid subtypes). Chaetaria [characteristic patches of bristles on the basiala, see Figure 7 ] present on the bases of the Radial and the Second Anal vein, but small and weakly expressed.
Coverings of the wing
Functional features
In evaluating the features of the tabanoid type, one should first of all direct one's attention to the very high indexes (numerical values) of the load (weight of the insect per surface-unit of wing). In fact to this type do belong forms displaying the greatest load among all Diptera (such are species of robber flies of the genus Laphria, of which the load is up to 0.3 gr/cm2 ). The high load is partly the result of great absolute size (and, consequently, weight) of the body, which size is very large in these insects. As already told, wings of this type are possessed by the largest gigantic forms of recent Diptera (Acanthomeridae, Mydaidae, Asilidae). Therefore the analysis and evaluation of features of the traction-lifting many-veined (tabanoid) type should be carried out while the size and weight of the body of the insect are contantly taken into account : only in complying with this condition it is possible to uncover the functional significance of the features of the wings and the reasons of preservation or change of this or that structure of their skeleton (venation). As was stated already above, of this type a rich, little reduced venation is characteristic. Earlier I [Rohdendorf] even had called it "the most archaic", " having been preserved almost with certainty since jurassic times" (Rohdendorf, 1949, p.99-100). At that time the structure of the basiala of these flies was preliminary described by me, while I remarked that these diptera with their "weakly costalized wings b y c o n t r a s t were provided with a complex basiala" (ibidem, p.103), and that these insects possess "the most primitive forms" of flight apparatus (p.150). It must now be said that, evidently, this analysis was insufficient, which analysis, as to the mechanics of the wing, only concerned the skeleton of the wing without taking into account the size of the body of these insects. As a result only the peculiarity of the structure of the wings was recorded, viz., the "ancientry" of the venation of the wing-blade, together with the high specialization of the basiala, while a clarification of this process was not attempted. The real reason-to-be of the features of tabanoid wings consisted in the perfection of the mechanical properties of the wings in diptera of large size. Increase of size of the insect inevitably brings with it an increase of load per unit surface of the wings. Therefore the processes of costalization, the strengthening of the anterior margin of the wing as an adaptation to an increase of wing-beat frequency (with a growth of the forces of traction and lifting), follows a different path, different from that of small (or in the process of becoming small) insects. Large insects cannot realize costalization of the wing by displacements and reduction of veins, because when this happens the strength of the blade of the wing as a whole will decrease, which is not allowed by the increase of the load. The strengthening of the anterior margin is realized by the thickening of the anterior veins (costal, radial, and subcostal) and [the mechanical features of the wing are further worked out] by the perfection of the mechanism of transition [of muscle force] of the wing, its basiala (formation of a phragm, strengthening of the cells of the basiala). The wing-venation [that is the venation of the wing-proper, the wing-blade] remaining rich, essentially not being reduced, turns out to be a very useful supporting skeletal element of a large wing. The veins of the wing only insignificantly changed their mutual position, guaranteeing a firmness of certain parts of the wing-blade and, probably, playing an important role in the creation of air-whirls during wing-beat. Consequently, the assertion of "primitiveness" of the tabanoid type of wings is not correct : Only the plan of venation of the wing-blade is primitive, whereas the whole wing [and thus including its basiala] of the tabanoid type is not at all primitive, but represents a characteristic form of perfected flight-organs of large diptera. The rich venation on the wing-blade of these flies, although being a relict feature [i.e. a feature that has been preserved], nevertheless plays an important functional role.
History and transformations of the type
|
Venational Prototype of the Order Diptera |
Figure 3 : Traction-lifting many-veined (tabanoid) wings of fossil representatives of the rhagionoid and eostratiomyioid subtypes.
Upper image : Rhagiophryne bianalis Rohd. ( Rhagionidae ), Upper Jurassic of Karatau, southern Kazachstan. Length 4.25 mm.
Lower image : Eostratiomyia avia Rohd. ( Eostratiomyiidae ), Upper Jurassic of Karatau, southern Kazachstan. Length 7 mm, width 2.5 mm.
(After ROHDENDORF, 1951)
Figure 4 : Traction-lifting many-veined (tabanoid) wings of fossil representatives of the palaeostratiomyioid and rhagionoid subtypes.
Upper image : Palaeostratiomyia pygmaea Rohd. ( Palaeostratiomyiidae ), Upper Jurassic of Karatau, southern Kazachstan. Length 2.5 mm.
Middle image : Rhagionempis tabanicornis Rohd. ( Rhagionempididae ), Upper Jurassic of Karatau, southern Kazachstan. Length 3.5 mm.
Bottom image : Protorhagio capitatus Rohd. ( Rhagionidae ), Upper Jurassic of Karatau, southern Kazachstan. Length 5 mm.
(After ROHDENDORF, 1938, in ROHDENDORF,1951)
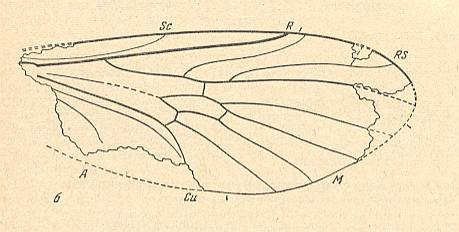
Figure 5 : Traction-lifting many-veined (tabanoid) wing of a fossil representative of the rhagionoid subtype.
Wing of Protorhyphus turanicus Rohd. ( Protorhyphidae ). Liassic (lower Jurassic) of Issic-Kul. Length of remains 2.7 mm. Length of the whole wing about 3.0 mm.
(After ROHDENDORF, 1962, in ROHDENDORF, 1964)

Figure 6 : Traction-lifting many-veined (tabanoid) wing of a fossil representative of the rhagionoid subtype.
Wing of Protorhyphus stigmaticus Handl. ( Protorhyphidae ). Upper Liassic (lower Jurassic) of Mecklenburg (northern Poland [formerly Germany] ).
(After HANDLIRSCH, 1938, in HENNIG, 1954)
These families were : Protorhyphidae (suborder Nematocera) - ancestors of the recent Phryneidae (= Rhyphidae), and of the suborder Brachycera-orthorrapha : Rhagionidae (preserved also in the recent fauna), Archisargidae, Palaeostratiomyiidae, Rhagionempididae and Eostratiomyiidae (the last four families are only known from the Jurassic). All these ancient mesozoic diptera were at the time already fairly diverse : The Protorhyphidae (Nematocera) differed from the complex of Brachycera only by the fact that in them the anal and cubital veins were relatively far apart from each other. And all five brachycerous families also were not uniform as to the structure of the wings, the difference of which forces us to divide them into ten more subtypes (in addition to the phryneoid subtype) of the tabanoid type.
1. The wings of the Phryneidae [= Rhyphidae] (Nematocera) -- see Figure 1, top image -- strongly differ from the other representatives of the tabanoid type by their small size and a smaller number of Radial branches of wich there are only three (including the Radius-proper). The Medial veins form a well-expressed intermedial cell (discoidal cell) and count four branches whereby the posterior branch [M4] is not the continuation of the cross-vein mcu [better written as m-cu], but being close to the common Medial trunk. The anal vein runs far away from the Cubitus. It does not coalesce with it nor coming near to it.
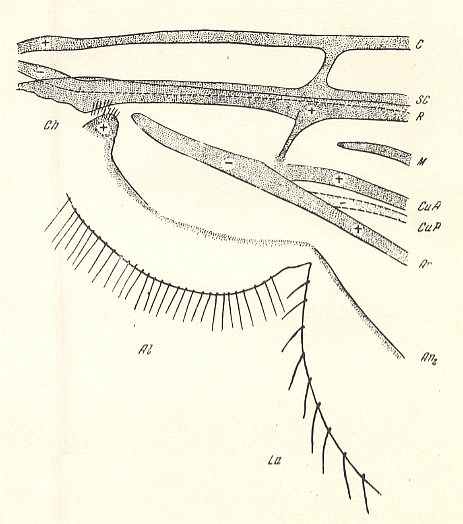
Figure 7 :
Basiala (schematically) of the wing of Phryne sp. ( Phryneidae ).
At the base of the Radius (R) and of the second anal vein (An2) we see the chaetaria, and from the Radius projecting downwards, we see the phragm (or that by which it is replaced).
(After ROHDENDORF, 1946)
In the basiala (generally) of this subtype, instead of a phragm there is only a well-visible fold.
2. The jurassic Protorhagionidae (the genera Protorhagio, Figure 4, bottom image , and Rhagiophryne, Figure 3, top image ) and their descendants, the caenozoic Rhagionidae, Figure 1, second image , also living today, form the special rhagionoid subtype, characterized by the moderate size of the body, the weak development of the phragm (in the basiala), instead of which there is only a more or less expressed fold. However, in contrast to the previous subtype, the number of Radial branches is not reduced, having remained four [including the Radius-proper]. Moreover, an approach of the first anal vein and the Cubitus is evident, converging to the margin of the wing. The fork of the two last Radial branches is long and narrow. The posterior Medial branch is differentiated from the remaining system. Its base changes direction and becomes transverse. The hind-margin of the wing, as a rule, with the Costal vein [i.e. it is not reduced there]. In repose the wings are directed obliquely backwards. This subtyp is a derivative of not precisely known forms of the Tipuloid type, forms that have given rise to the Bibionoid and Fungivoroid types. In its turn, the rhagionoid subtype was the point of departure of a series of other forms of tabanoid wings, namely of the already above described phryneoid subtype, further of the tabanoid-proper, the erinnoid, and mydaoid subtypes (see below).
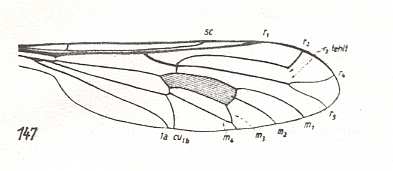
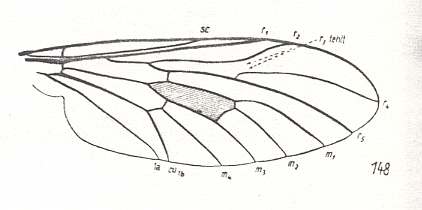
Figure 8 :
Left image (147) - Wing of Lampromyia pallida Meig. Rhagionidae. In all Rhagionidae the vein R3 [also written as r 3] is missing. In this wing we can see where it originally was. Because of its slender form this wing might belong, not to the rhagionoid, but to the erinnoid subtype (see below).
Right image (148) - Wing of Rhagio scolopaceus Lw. Rhagionidae.
(After HENNIG, 1954)
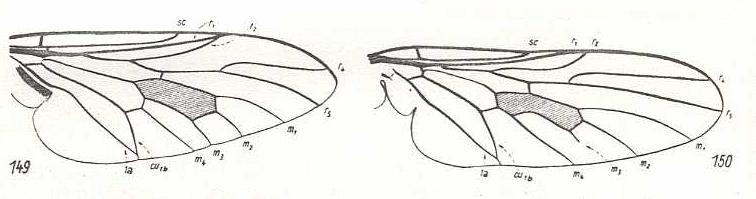
Figure 9 :
Left image (149) - Wing of Atherix ibis Fabr. Rhagionidae. The vein R2 [= r 2 = anterior branch of Radial Sector] ends up in R [= r 1].
Right image (150) - Wing of Spaniopsis clelandi Ferg. Rhagionidae.
(After HENNIG, 1954)
3. Special forms of tabanoid wings were shown in the specialized jurassic diptera, the Eostratiomyiidae ( Figure 3, lower image ), which form the peculiar eostratiomyioid subtype. This family distinguishes itself by a relatively large wing-size, by the presence of four Radial branches, several interradial and radio-medial cross-veins, the absence of the Costal vein at the wing's hind-margin, and by a broad wing with a pointed and well-differentiated apex [differentiated, that is, from the rest of the wing]. This subtype is all-out idiosyncratic, being characterized by very ancient primitive features (cross-veins) together with a great mechanical perfection of the shape of the wing. Unfortunately the structure of the basiala could not be investigated in this fossil. Apparently, the phragm was not developed -- this assumption is based upon the nature of the venation of the wing-blade, which (venation) is relatively weak. The origin of the eostratiomyioid subtype is not clear, although, apparently, one may be convinced that there does not exist a direct affinity of this subtype with rhagionoid wings. The eostratiomyioid subtype is, probably, a derivative of certain not precisely known forms of that same original tipuloid type, namely of those forms in which there was still an abundance of cross-veins and an overall rich wing-venation in the absence of mechanical specializations in the structure of the basiala. This remarkable subtype probably was the point of departure of the development of two others : the extinct palaeostratiomyioid and nemestrinoid subtypes, represented also in the recent fauna.
4. Special jurassic diptera, namely two representatives of the families Palaeostratiomyiidae ( Figure 4, upper image ) and Rhagionempididae ( Figure 4, middle image ) possess peculiar wings the structure of which may be derived from the previous subtype, and which form the special palaeostratiomyioid subtype. Of this subtype small sizes are characteristic, further (are characteristic) a marked development of costalization, appearing in the form of the thickening of the Costal and Radial (its two anterior branches, that is, the Radius-proper and a branch of the Radial Sector) veins, a broad costal field, the peculiar shape of the broad wings truncated at the apex, and the reduction of the Costa at the posterior wing-margin. Characteristic of one form, (Rhagionempis, Figure 4, middle image ), is a perfection of mechanical qualities, expressed by the strong divergence of the two posterior Radial branches, reminding us of the analogous phenomenon in proper-tabanoid wings. The phragm (in the basiala) was absent and in its place there was merely a fold. The posterior Medial branch is situated in the original fashion, branching off from the main trunk of the Media. Cross-veins present only in the form of the basic r-m and m-cu [the presence of the latter seems to be assumed by Rohdendorf]. This subtype, only known as two fossils, undoubtedly shows a high degree of specialization, while the possibility is not excluded that it is further divided into two independent groups. The latter assumption is evoked by the marked differences in the structure of the wings of those two species. Being a derivative of the eostratiomyioid subtype, the present subtype, probably, is one of the sources of the stratiomidoid type [the next main type, being discussed later].
5. Several jurassic, tertiary, and recent diptera, belonging to the relict groups Erinnidae (Figure 10, left image), Solvidae ( Figure 1, third image and Figure 10, right image ), Vermileoninae, and certain Cyrtidae (the genera Cyrtus, Opsebius) possess peculiar wings of the tabanoid type, and as such form the special erinnoid subtype. The shape of the wings of this subtype is elongated. The phragm (in the basiala) is in the form of a fold, more rarely well developed (Cyrtidae). Especially characteristic is the form of the fork formed by the two posterior Radial branches, which (fork) is short and fairly wide. The hind-margin of the wing is often without Costal vein. The posterior medial branch [M4] is individualized with respect to the rest of the Medial system of veins [its basal section looks like a cross-vein]. How the wings are held in repose is not very well known to me [Rohdendorf]. This subtype is a derivative of rhagionoid wings, and in turn was the point of departure of the development of the cyrtocioid subtype and the stratiomidoid type. To the present subtype belong little studied insects, in their majority rare forms.
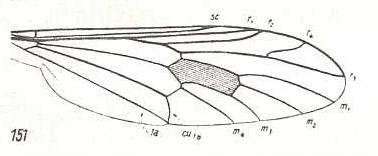
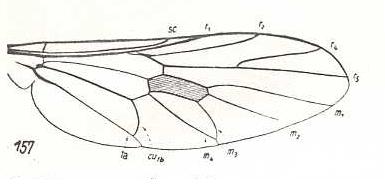
Figure 10 :
Left image (151) - Wing of Erinna atra Meig. Erinnidae.
Right image (157) - Wing of Solva marginata Meig. Solvidae. The veins M3 and M4 [= m 3 and m 4] have a common ending at the posterior wing-margin, so also the veins CuA and A1 [= Cu1b and 1a]. The vein M2 [= m 2] has lost its end-part (i.e. it is retreating from the wing-margin).
(After HENNIG, 1954)
It might be that also the above mentioned Lampromyia (Rhagionidae) belongs to this subtype.
6. Another derivative of the rhagionoid subtype form the wings of horse flies (Tabanidae), pseudo-robberflies (Therevidae), Coenomyiidae, and, probably, the tropical Acanthomeridae not known to me [Rohdendorf] in nature. For this truly extensive and rich collection it is natural to let it represent the tabanoid-proper subtype. Examples : Figure 1, fourth image , and next Figures.
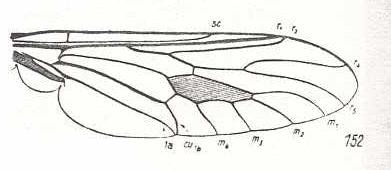
Figure 11 :
Wing of Coenomyia ferruginea Scop. Erinnidae. Here, the basal section of M4 is suppressed by CuA [= Cu1b].
(After HENNIG, 1954)
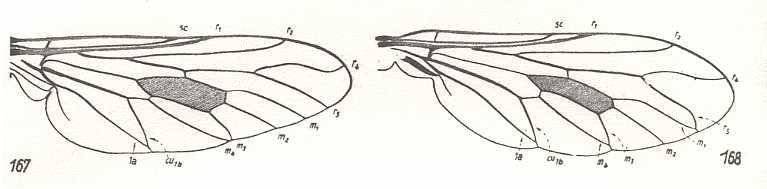
Figure 12 :
Left image (167) - Wing of Thereva nobilitata Fabr. Therevidae.
Right image (168) - Wing of Chrysanthemia chrysanthemi Fabr. Therevidae.
(After HENNIG, 1954)
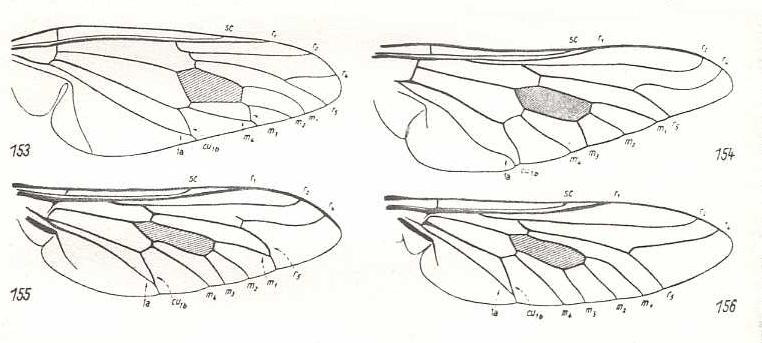
Figure 13 :
Upper-Left image (153) - Wing of Lycops zoos End. Pantophthalmidae.
Upper-Right image (154) - Wing of Pelecorrhynchus personatus Walk. Tabanidae
Bottom-Left image (155) - Wing of Pangonia maculata Rossi. Tabanidae.
Bottom-Right image (156) - Wing of Tabanus bromius L. Tabanidae.
(153 after ENDERLEIN, 1931, in HENNIG, 1954. 154-156 After HENNIG, 1954)
This subtype is characterized by a number of features : The phragm (in the basiala) well expressed in the form of a firm oblique outgrowth connecting the basal part of the Radial vein with the Cubital and the first Anal. In addition, the basiala in the representatives of this subtype is characterized by the development of a broadening of the base of the Costal vein, by the presence of well-developed wing and thoracic scales [i.e. a wing scale and a thoracic scale]. The two posterior Radial branches are strong, and sharply diverging, where the anterior one ends up at the anterior margin, while the posterior one ends up at the margin of the wing only after the latter's tip. There are four Medial branches, where the posterior branch usually is individualized, more rarely tigthtly unified into one common complex with the three other anterior branches (Coenomyiidae, see Figure 11 [ There, the genus Coenomyiia is assigned to the famliy Erinnidae (by Hennig) ] ). The veins at the posterior margin of the wing often form closed cells, or cells that are narrowed toward their end (for example the cubital cell, as a result of the coalescence of the terminal portions of Cu and An1 [= CuA and 1A, or CuA and 1a] ( Figure 13, 153-155-156 ), the fifth Radial cell as a result of such a coalescence of M1 and R5 [= m1 and r5] ( Figure 13, 155 ), the third Medial cell as a result of such a coalescence of M3 and M4 [= m3 and m4] ( Figure 13, 153 )). The posterior margin of the wing is provided with a costal vein (C, Costa). The outline of the wing is fairly broad, with a well-differentiated apical part often with a pointed apex, a convex anal lobe, a large alula (winglet) and scales [the wing-scale and the thoracic scale]. This subtype is known only to have existed since the beginning of the Cenozoic (Teriary and Quarternary). In the Mezozoic fauna there is, until now, not found any form carrying tabanoid wings. The connections of this tabanoid-proper subtype are fairly clear : Undoubtedly, from the base of tabanoid wings the asiloid and anthracoid wings have evolved.
7. One of the derivatives of the tabanoid-proper subtype of wings is expressed by representatives of the large family of robber flies (Asilidae). See Figure 1, fifth image . This asiloid subtype is characterized by the positioning of the wings in repose : they are held folded over the abdomen, and by the development of an all-out firm phragm, looking like a strong outgrowth, unifying by means of a special kind of hinges the basal sections of the Radial and Cubital veins. Another characteristic feature consists of the sclerotization, thereby transformed into a firm surface, of the whole central cell of the basiala : Essentially this cell is, qua firmness, the direct continuation of the phragm. The venation of the elongated wing-blade is characterized by the very long first Radial vein (R), which often unifies its end, not with the Costal vein, but with the next, second, Radial vein. The posterior branches of the Radial veins are variable. Usually they strongly diverge, more rarely they run parallel, and proceed in certain cases to the wing's hind-margin (Leptogastrinae), in other cases to the front-margin (several Asilinae, for example the genus Satanas). The Medial cells, as also in the previous subtype, often closed. The hind-margin[al area] of the wing is often attenuated. The Costal vein runs all around the whole wing, more rarely only reaches the Cubital vein. Sometimes the posterior margin is devoid of the Costal vein, whereby the venation of the posterior margin is also weakened. The in this way costalized wings are found in some Asilinae and Dasypogoninae. The anal lobe and the alula are well developed. Only in the Leptogastrinae the wings are narrow and almost without such structures. The thoracic scale is rudimentary. Of this subtype is, in addition, very characteristic the small size of the wings, which are shorter than the body. This subtype is known to exist only since Tertiary times, apparently already represented in the faunae by recent subfamilies and by almost exclusively recent genera. From Mesozoic faunae the asiloid subtype is not known. Transformation in this subtype consists of the development of a high degree of costalization, on the one hand, and of the formation of the narrow specialized wings of the Leptogastrinae, on the other. It is possible that these forms allow them to be individualized as separate subtypes. The next Figure provides some more examples of this subtype of wings.
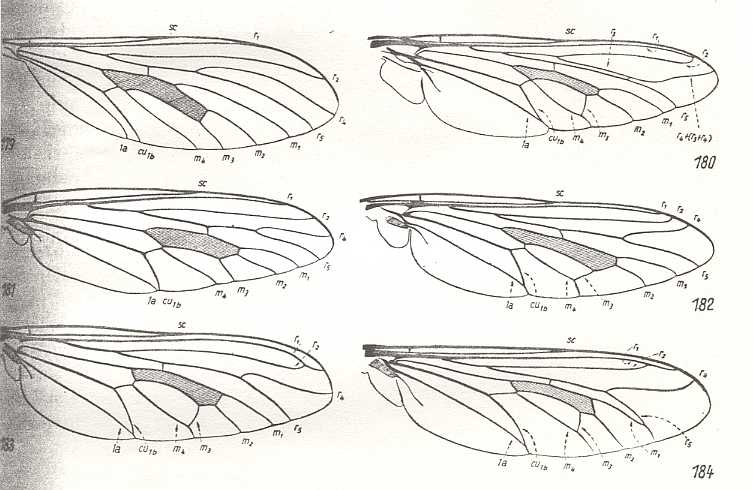
Figure 14 :
Upper-Left image (179) - Wing of Leptogaster cylindrica Deg. Asilidae.
Upper-Right image (180) - Wing of Promachus leoninus Lw. Asilidae
Middle-Left image (181) - Wing of Heteropogon scoparius Lw. Asilidae.
Middle-Right image (182) - Wing of Bathypogon brachypterus Macq. Asilidae.
Bottom-Left image (183) - Wing of Laphria marginata Lw. Asilidae.
Bottom-Right image (184) - Wing of Dasythrix grisea Herm. Asilidae.
(After HENNIG, 1954)
In the next sections we will have to do with families such as Bombyliidae (bee-flies), Mydaidae, and Nemestrinidae. For good orientation we here give some Figures :

Bombilius major. 8-12 mm. Bombyliidae. |

Leptomydas corsicanus. Mydaidae. |

Fallenia fasciata. Nemestrinidae. |
8. Another sort of derivative of the tabanoid-proper wings we can see in many representatives of the extensive and diverse family of bee flies, Bombyliidae (especially of the group Anthracinae and some other groups). These characteristic forms of tabanoid wings constituting the special anthracoid subtype, distinguish themselves by many features shared with the previous, asiloid, subtype. Similar is the development of a firm basiala, having a thickened Costal vein, a sclerotized central cell and a strong phragm. Also the anterior Radial branch (R) is significantly lengthened, and the wing-size has increased markedly. See next Figure (upper image).
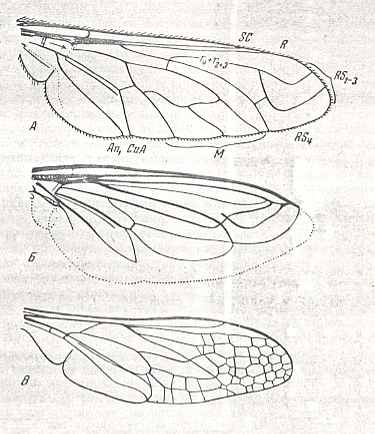
Figure 15 : Recent traction-lifting many-veined wings of the anthracoid, mydaoid, and nemestrinoid subtypes.
Upper image - Wing of Exoprosopa sp. Bombyliidae. Length about 20 mm. (After HENDEL). y = phragm (see the "y" sign and the arrow added [by Rohdendorf] in the drawing of the wing).
Middle image - Wing of Eremomydas bek Sem. Mydaidae. Length 14.5 mm. (After ROHDENDORF, 1951).
Bottom image - Wing of Nemestrellus abdominalis Oliv. Nemestinidae. Length about 20 mm. (After SACK)
(In ROHDENDORF, 1951)
But totally differently structured is the venation of the wing-blade. The posterior Radial branches are, as a rule, peculiarly curved, unified one with the other by means of characteristic cross-veins (usually one in number, more rarely two). Of Medial veins there always are no more than three, whereby one of the middle branches is reduced. Features of costalization are only apparent by the strengthening of the marginal vein (Costa) and the base of the Radius. This subtype is, as is the previous one, known only from the beginning of the Tertiary. Study of Tertiary representatives of the Bombyliidae does not add much to our knowledge of the history of this subtype.
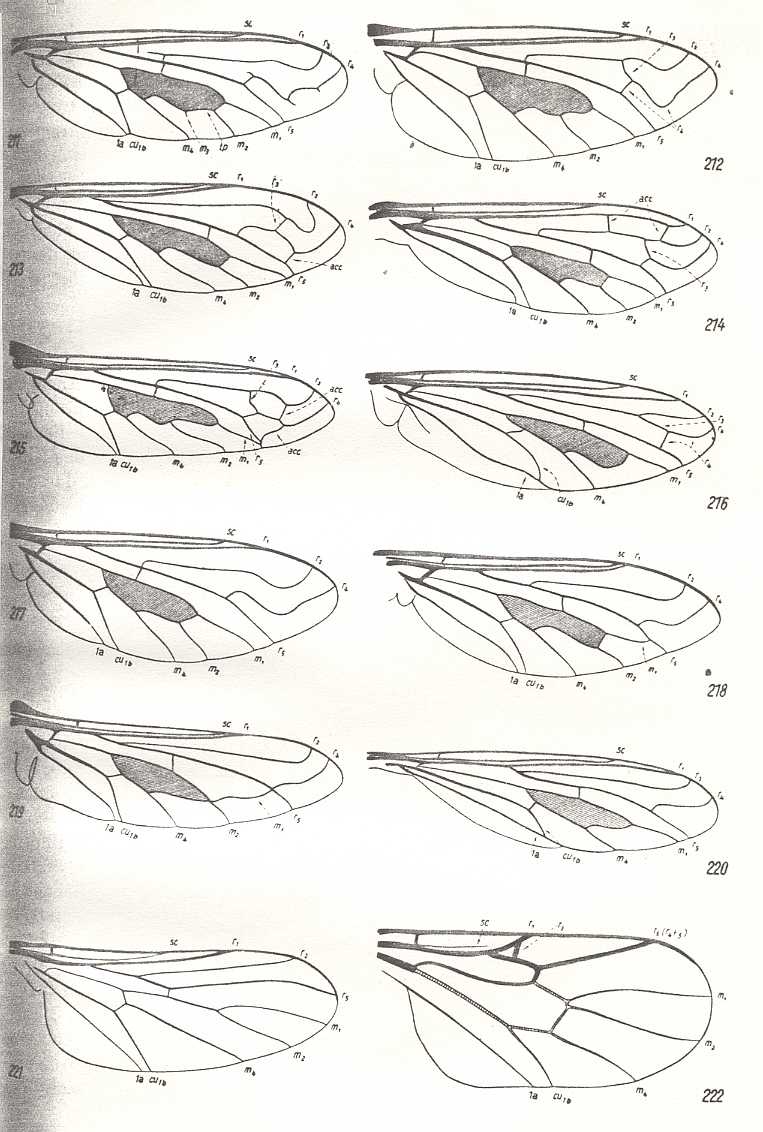
Figure 16 : Wings of Bombyliidae. From left to right and from top to bottom :
| 211 - Wing of Argyramoeba oedipus Fabr. | 212 - Wing of Exoprosopa stupida Rossi. |
| 213 - Wing of Ligyra lugubris Rond. | 214 - Wing of Henica longirostris Wied. |
| 215 - Wing of Hyperalonia bombyliformis Mac Leay. | 216 - Wing of Toxophora maculata Wied. |
| 217 - Wing of Thyridanthrax afer Fabr. | 218 - Wing of Callistoma fascipennis Macq. |
| 219 - Wing of Heterostylum robustum O.S. | 220 - Wing of Systropus excisus End. |
| 221 - Wing of Cyrtosia meridionalis Rond. |
222 - Wing of Glabellula nobilis palaestinensis Engel. (After ENGEL, 1933) |
Sometimes the venation of a representative of a given family can be the product of a transformation of the f a m i l y-prototype such that the latter is hardly recognizable anymore even to the extent that the product-venation is almost identical to the typical venation of a non-aberrant representative of another suborder! The next Figure shows this for the venation of the bee fly Empidideicus :
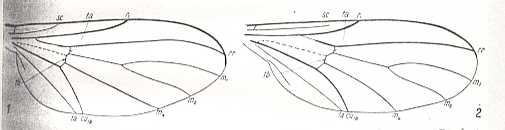
Figure 17 : Convergence of wing-venation.
Left image - Wing of Diadocidia spec. (Suborder Nematocera, Diadocidiidae).
Right image - Wing of Empidideicus spec. (Suborder Brachycera-orthorrapha, Bombyliidae).
Both wings evidently do not belong to the tabanoid type, but probably to the ancient lifting venationally-attenuated (fungivoroid) wing type (discussed in Part LXIX )
(After EDWARDS, 1926, in HENNIG, 1954)
9. A number of forms of the family of bee flies (Bombyliidae), namely the genera Usia, Cyrtosia (221 of Figure 16), Systropus (220 of Figure 16), Apolysis, Oligodranes, Alloxytropus, and species of the peculiar genus Omphrale (Omphralidae) (next Figure), show a characteristic reduction of the venation, its costalization. These wing-forms must be taken apart to constitute the special cyrtosioid subtype.
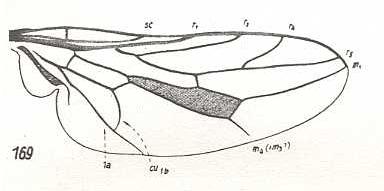
Figure 18 : Wing of Omphrale fenestralis Lw. Omphralidae. Compare with Cyrtosia (Bombyliidae) ( Figure 16, 221 ).
(After HENNIG, 1954)
The basiala of the wings of this subtype is to a lesser extent strengthened than it is in the anthracoid subtype, lacking a phragm or only with a weak phragm or fold. The wing-venation of the wing-blade is markedly reduced. Of the Radial branches there are four, sometimes only three (Cyrtosia (Fig. 16-221). Of the Medial veins there are almost always only two branches, and only sometimes a third branch is seen (genera Alloxytropus, Cyrtosia). Especially characteristic is the weakening of the hind-margin of the wing, lacking the Costal vein. This subtype is established exclusively on the basis of recent forms. There is no fossil material.
10. The chiefly tropical and subtropical diptera, the representatives of the families Mydaidae and Apioceratidae, possess wings that form a special transformation of the tabanoid type and which may be called the mydaoid subtype. These forms stand more closely to those of the asiloid subtype and distinguish themselves by the all-out peculiar structure of the venation of the apical part of the wing, and by the nature of its hind-margin. Mydaoid wings are, as to the structure of their basiala, similar to asiloid and tabanoid-proper wings (see Figure 7, Part LXXI ). In them there is a well developed strong phragm, a large alula and wing-scale. The Subcostal (SC) and anterior Radial vein (R) are very long, where the latter, just as in many representatives of the asiloid subtype, ends up, not at the wing-margin, but unifies at its end with other Radial veins. See Figure 15, middle image . But most remarkable is the structure of the posterior Radial and all Medial veins, which do not end up at the wing-margin, but unify at their ends one with the other, forming peculiar closed cells, or, on the contrary, partly arch-wise curve to the wing-apex and end up at the Costal vein on the anterior wing-margin or even at the anterior Radial vein. The Costal vein only extends up to the wing-apex and no farther. The whole posterior margin[al area] of the wing is without veins, and only the apices of the Cubital vein, of the posterior, more rarely also of the middle (Apioceratidae) Medial veins, reach the wing-margin. Sometimes also these veins do not reach the margin. The wing of the representatives of this subtype is very peculiar indeed by its bow-like veins in the apical part of the wing, and by the free, lacking veins, membrane that fringes in the form of an edging the posterior wing-margin. This subtype is most elegantly expressed in the representatives of the family Mydaidae. The species of the family Apioceratidae are distinguished by the lesser degree of development of an attenuated hind-margin. The Mydaoid subtype stands closely to the asiloid subtype, but is its derivative only partly. The connections of these subtypes are undoubtedly more complex and consist in their origin from the rhagionoid subtype, whereby the asiloid wings branched off from a derivative of the latter, namely from the tabanoid-proper subtype (from wings of the type as they are seen in Therevidae ( Figure 12 )), while mydaoid wings were formed not only from the tabanoid-proper subtype (also via Therevidae to Apioceratidae), but also directly from rhagionoid wings. There are practically no fossil documents that could throw light on the history of mydaoid wings, if we do not count the single finding of not-determined forms of Mydaidae from miocene deposits of North America.
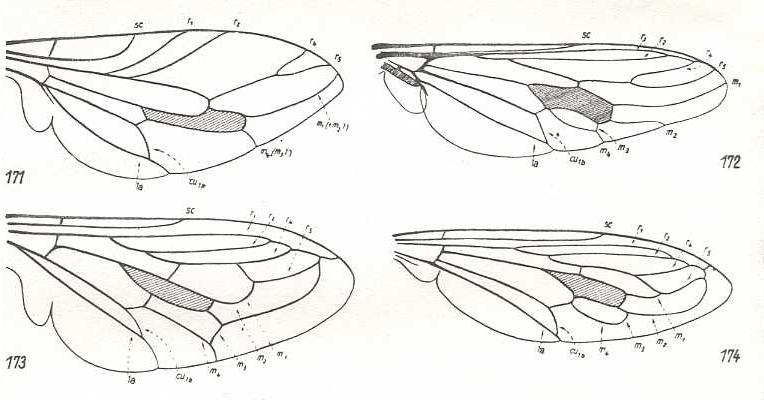
Figure 19 : Apioceratidae (= Apioceridae).
171 - Wing of Apiocera maritima Hardy. (After CAZIER, 1941).
172 - Wing of Apiocera bigoti Macq. (After HENNIG, 1954).
173 - Wing of Megascelus nigricornis Phil. (After CAZIER, 1941).
174 - Wing of Neorhaphiomidas hardyi Norr. (After CAZIER, 1941).
(In HENNIG, 1954)
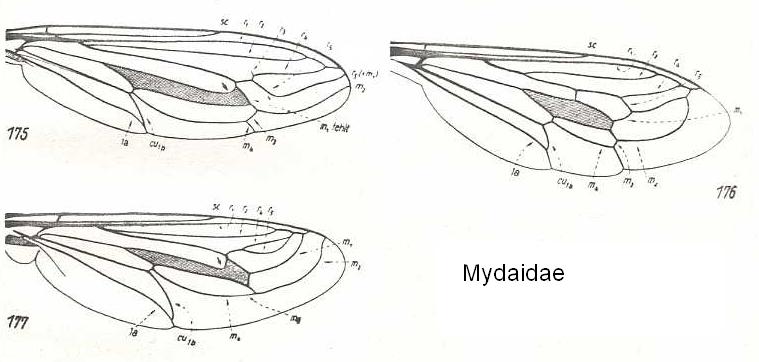
Figure 20 : Mydaidae.
175 - Wing of Rhopalia paulseni Phil.
176 - Wing of Dolichogaster nigricornis Phil.
177 - Wing of Mitrodetus dentitarsis Macq.
(After HENNIG, 1954)
11. The wings of the most peculiar diptera, Nemestrinidae ( Figure 15, bottom image ), together make up a special subtype of tabanoid wings, which might be called the nemestrinoid subtype (see also the Figures below ). This subtype may be best characterized by the almost constant presence of "supplementary veins" (in reality undoubtedly remnants of an ancient rich venation!), chiefly in the form of many cross-veins, positioned between the branches of the Radial and Medial systems. Another characteristic feature of this subtype is the development of a special so-called diagonal vein, made up by sections of radial and medial veins, and positioned obliquely across the wing from the basal part of the anterior margin down to the middle of the termen (see Figure 15, bottom image ). This complex additional structure delimits the apex from behind [i.e. forms the posterior-proximal border of the apical region of the wing), and probably has a certain mechanical function or significance. Nemestrinoid wings are, in addition, characterized by the bend of all Radial and anterior Medial veins to the apex of the wing. These branches run parallel to the hind margin and end up at the anterior margin of the wing-apex (see Figure 21, below ). This similarity with the mydaoid wings is supplemented by a certain attenuation of the whole hind-margin[al area] of the wing, which sometimes lacks the Costal vein, and has a free membraneous zone (for instance in the genera Atriadops, Fallenia [193 in Fig. 21] ) [This, is, by the way, also seen in many hymenopterous wings.]. Very characteristic of this subtype is the great enlargement of the basiala, reaching a large size indeed, sometimes up to more than one fourth of the length of the whole wing. The phragm is almost always very strong [forming a structure that is analogous with the Radio-Medial T-vein in many Hymenoptera-Apocrita] in the form of an oblique firm vein between the base of the radial and cubito-anal veins (see Figure 2, above ). Anal lobe, as a rule, well developed, seldom narrow (in the genera Nycterimorpha [197 in Fig. 22], Cyclopsidea [196 in Fig. 22] ). Alula almost always large, seldom being absent (in the genera Cyclopsidea, Nycterimopha), or rudimentary (in the genera Fallenia [193 in next Figure], Symmictoides [187 in next Figure], Symmictus). The wing-scale is rudimentary.
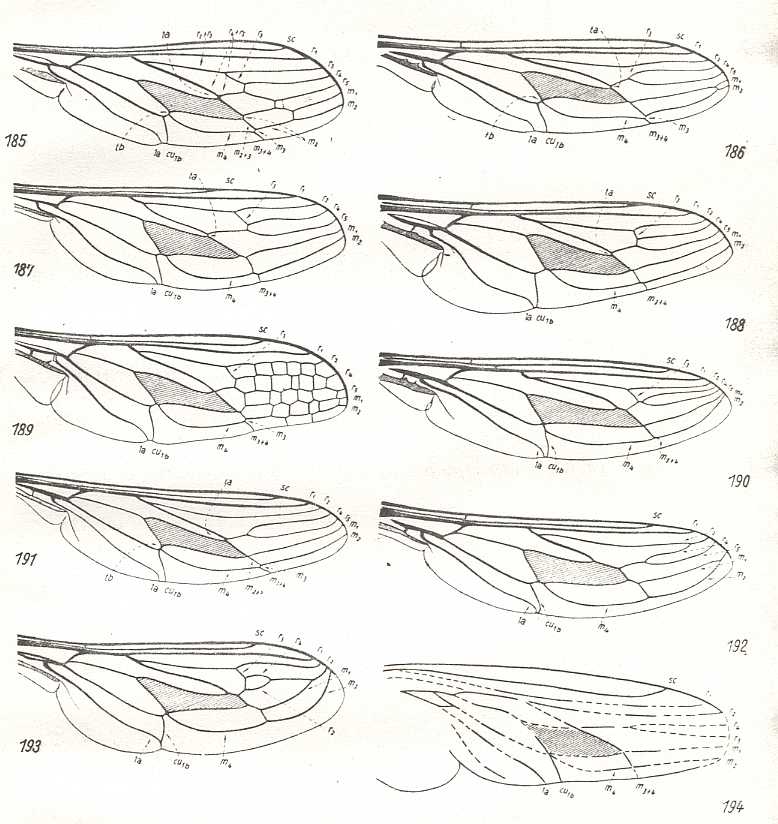
Figure 21 : Nemestrinidae.
|
185 - Wing of Hirmoneura bellula Phil. |
186 - Wing of Hirmoneura obscura Meig. |
|
187 - Wing of Symmictoides simplex Lw. |
188 - Wing of Rhynchocephalus fasciatus Macq. |
|
189 - Wing of Nemestrinus aegyptianus Wied. |
190 - Wing of Neorhynchocephalus tauscheri Fisch. |
|
191 - Wing of Trichophthalma barbarossa Big. |
192 - Wing of Stenopteromyia bolivari Strobl. |
|
193 - Wing of Fallenia fasciata Fabr. |
194 - Wing of Prohirmoneura jurassica Handl., from the upper Jurassic (Malm) of Bavaria, Germany. (After HANDLIRSCH, 1908) |
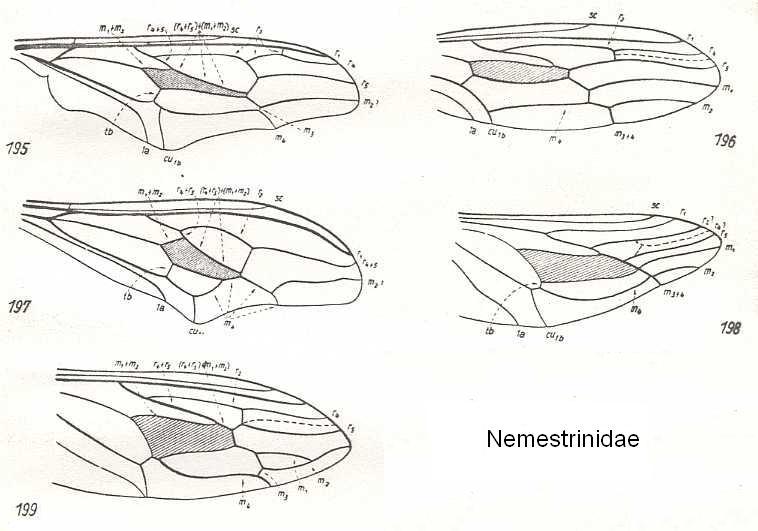
Figure 22 : Nemestrinidae.
195 - Wing of Nycterimyia horni Lichtw. (after the type in Deutsches Entomologisches Institut)
196 - Wing of Cyclopsidea spec. (After HARDY, 1946)
197 - Wing of Nycterimorpha speiseri Lichtw. (after the type in Deutsches Entomologisches Institut)
198 - Wing of Atriadops spec. (After HARDY, 1946)
199 - Wing of Trichopsidea spec. (After HARDY, 1946)
(After HENNIG, 1954 (except 196, 198, 199))
Of fossil documents on the history of the nemestrinoid subtype there are only a few. The sole mesozoic representative of the family Nemestrinidae is found in the upper Jurassic deposits (Malm) of Bavaria (Germany) and was described by Handlirsch under the name of Prohirmoneura jurassica Handl. (see 194 in Figure 21 ). This form is very insufficiently studied, and to judge about its features from the existing description and drawing of Handlirsch is not possible. Only its being close to the family Nemestrinidae is clear : the absence of a long snout [in the complete fossil] was evidence for the author to assume it to be close to the genus Hirmoneura, which has little basis. The venation of the wings is not described and not drawn in an exact way. Other fossil nemestrinids are known only from the cenozoic. Such are a few species of recent genera from the Miocene of North America and from the upper Oligocene of France. All fossil material, as to the history of the nemestrinoid subtype, does throw almost no light on the way of its formation.
General considerations concerning the tabanoid type
We have now concluded the description of the tabanoid functional wing-type and its subtypes.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXXIa.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII
Back to Evolutionary Part LXVIII
Back to Evolutionary Part LXIX
Back to Evolutionary Part LXIXa
Back to Evolutionary Part LXIXb
Back to Evolutionary Part LXXa
Back to Evolutionary Part LXXb