

(Description of stratiomyoid wing-type according to ROHDENDORF, 1951)
Representatives of the type
Lifting costalized (stratiomyoid) wings are typical to species of the brachycerous families Stratiomyidae, Chiromyzidae, many Cyrtidae and some Bombyliidae (the genera Empidideicus, Glabellula).
Size of the wings
The absolute size of the wings of this type is medium, more rarely small, from 3 to 15 mm, seldom larger. The relative size of the wings is short, almost always less than the length of the body, more rarely longer than it. The size of the surface area of the wings is known for some species of three genera of the subfamily Stratiomyinae, viz., of the genus Eulalia (0.151-0.287 cm2), of the genus Hirtea (0.416-0.713 cm2), and of the genus Stratiomys (0.529-0.769 cm2). Weight of the wings insignificant, reaching in the only species studied in this respect, a species of the genus Stratiomys, 1.34 percent of the weight of the whole insect. The load is studied in several species of the family Stratiomyidae and varies between 0.072-0.135 gr/cm2 in the small Eulalia and 0.116-0.204 gr/cm2 in the large Hirtea and Stratiomys.
Shape of the wings
Wings with a straight anterior margin, a small apex, individualized termen and a large anal lobe. The basiala is sharply distinguished from the wing-blade by the border of the anal lobe, and often carries a large alula which sometimes is absent (genus Glabellula and some others). Wings moderately elongate, usually 2.5-3 times longer than wide, more rarely a little longer. Seldom are the wings short, 2-2.33 times longer than wide (some Beredinae, and the genus Glabellula).
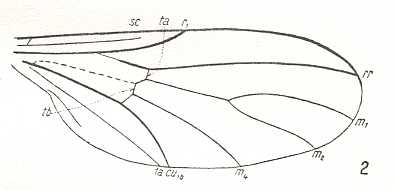
Skeleton of the wing
In resting position the wings are folded one above the other over the abdomen, sometimes obliquely so. Very characteristic of this type is the strong degree of costalization, consisting in strengthening, shift, and coalescence of veins in the anterior half of the wing, and reduction or weakening of the middle-part veins and a widening of the wing-membrane of the posterior half of the wing, free of veins. The Costal vein strong, almost never reaching the wing's apex. The Radial veins running closely together and thickened, all ending up at the anterior wing-margin at the Costal vein. Only sometimes the last Radial vein directed backwards (in the genera Oncodes and Corononcodes (see next Figure, third image))
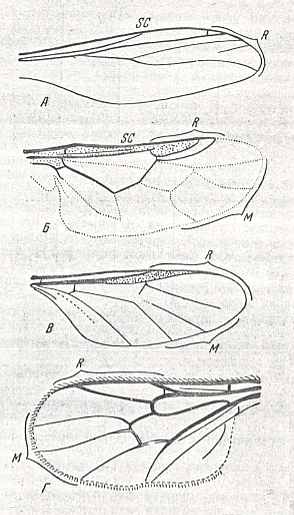
|
Figure 1 : Wings of representatives of the lifting costalized (stratiomyoid) type. From top to bottom :
|
The number of Radial veins [including the Radius proper] varies from 4 to 2. The Subcostal vein is weak, sometimes ending in the membrane of the costal field. Medial veins thin, in the form of 3 or 4 branches, usually forming the intermedial cell. Sometimes the number of Medial veins is reduced to two and the intermedial cell absent (in the genera Oncodes, Corononcodes, Philopota). The Cubital and first Anal vein are delicate, thin, and long, often coalesced at their end, forming the closed cubital cell (Stratiomyidae, Figure 1, second image), sometimes only converging or parallel (in the genera Glabellula and Oncodes). The cross-veins r-m and m-cu, as a result of costalization processes, sometimes completely disappear, or sharply change their position. Only r-m lies between the last Radial branch and the anterior Medial branch in the majority of Stratiomyidae.
The next Figure presents some more wings of the present type :
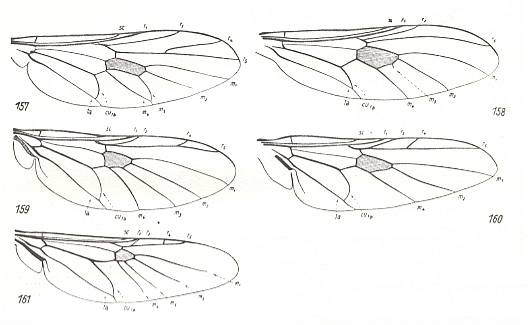
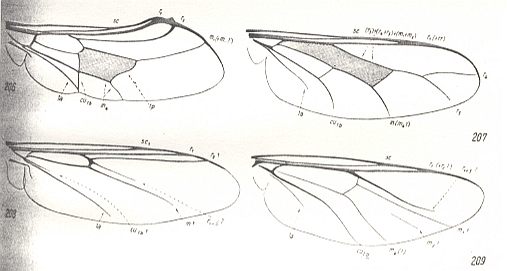
Figure 3 : Some more representatives of the Lifting Costalized (stratiomyoid) functional type of wings.
Cyrtidae [= Acroceridae = Oncodidae].
206 : Pterodontia misella O.S. 207 : Acrocera convexa Cole. 208 : Philopota aenea Meig. 209 : Oncodes zonatus Erichs. (After HENNIG, 1954)
Coverings of the wing
Functional characteristics
Only the general aspects of the way of life of these peculiar diptera are known. The chief group of the type, the Stratiomyidae, is connected with water-basins and rotting material, in which their larvae live. The winged insects are characteristic visitors of flowering plants, where they eat nectar. Other representatives of the type, Cyrtidae [= Acroceridae], develop in the body of spiders or in their egg-cocoons. The winged flies are encountered on flowers, and apart from this these flies are partly aphagous, having a reduced proboscis.
Finally, representatives of the family Bombyliidae, namely the species of the genus Glabellula, undoubtedly belonging to the present type, are very peculiar, distinguishing themselves by their extreme smallness (from 1.25 to 2.5 mm), and are found on flowers. The individual development of these bee-flies is not known [ROHDENDORF, 1951]. In all probability we may assume that their larvae are parasites of certain insects.
History and transformation of the type.
Fossil remains of the lifting costalized (stratiomyoid) type are few in number and until now [1951] limited to only two jurassic finds of single species of the families Archisargidae Figure 1, first image ) and Protocyrtidae, when not taking into account the many Tertiary forms of recent families and genera. To the latter belong many remains from the Oligocene of western Europe and North America, and from the Miocene of Eurasia and North America, belonging to species of the genera of the subfamilies Beridinae, Geosarginae, Stratiomyinae. The known mesozoic faunae of flies (diptera) (Karatau) include true representatives of the type, namely the highly specialized Archisargidae Figure 1, first image ) and Protocyrtidae. Certain jurassic forms from Karatau, namely species of the genera Palaeostratiomyia and Rhagionempis (See Figure 4 in Part LXXI ) possessing wings with rich venation of the tabanoid type, do show such features that point to their belonging to a special subtype, and together with this point to the evolutionary direction of the wing-venation directly leading to the stratiomyoid type. Another source of this type undoubtedly was formed by the representatives of the cyrtosioid subtype of tabanoid wings, in which costalization developed, which distinguished it from the traction-lifting many-veined (tabanoid) type and led to the formation of the stratiomyoid type. These forms are known as various, for a great part relict, genera in the recent fauna.
The different sources of the stratiomyoid type and its phylogenetically diverse content determine the existence of subtypes reflecting the systematic content of the type.
The largest of these, the stratiomyoid-proper subtype, includes species of the family Stratiomyidae and the relict Chiromyzidae [or Chiromyzinae, a subfamily of the Stratiomyidae, see Figure 2, above]. Despite the manifold of genera distributed among the many subfamilies of Stratiomyidae, there is no reason to further subdivide this subtype into subordinated categories. The variability of the wing-venation in Stratiomyidae only shows the different stages of costalization, chiefly connected with the absolute size of the insects and the larger or smaller intensity of flight. Precise analysis of the features of this subtype is a special task which I shall not perform here. We only may note that the group Beridinae could be characterized as having the least costalized wings.
The other, cyrtoid subtype, includes phylogenetically alien, very few forms, and is characterized by the extreme degree of costalization of the wing-venation (see Figure 1, third image and Figure 3 ) which has lost the majority of cross-veins (and with it having lost the closed cell in the middle of the wing!) and is markedly strengthened at the anterior margin. The larvae of the representatives of this subtype are highly specialized internal parasites of spiders.
The last, glabelluloid subtype consists of only two genera of Bombyliidae, Empidideicus and Glabellula (see Figure 1, fourth image ), which are minute insects (not larger than 2.5 mm), possessing extremely costalized venation, in which the Costal, Radial, Cubital, and cross-veins are significantly thickened and partly shifted distad. [Rohdendorf does not give a drawing of the wing-venation of Empidideicus. The only drawing we have so far seen is taken from HENNIG, 1954, see HERE. As one can see, this wing not at all belongs to the stratiomyoid type, because there is no trace whatsoever of any costalizaton of the venation]. The shape of the wing is truncated, reminding of certain Nematocera, namely various Oligoneura -- Scatopsidae and many fungivoroids. Until now [1951] the way of life of these peculiar diptera is unknown : the winged insects are found on flowers of various plants, and their larvae undoubtedly parasitize in whatever insects as is the case in all other species of the large family Bombyliidae.
A general evaluation of the evolutionary directions of the stratiomyoid type was already succinctly given earlier. We only may, concludingly, add that all features of this type evidently bear witness of the progressive development of the flight-function. Flight in the representatives of this type is a very important function in their life-activities, apparently connected with obtaining food and with reproduction. The chief causes, however, of the origin of this costalized type remain more or less unclear, i.e. the very significance and role of the development of costalized wings. As is known, such a wing-structure is widely distributed among diptera, and, probably, its origin was determined by certain conditions of perfection of the flight-function, which included concrete relationships between the weight of the insect and the demands of its way of life. These changes of the living-conditions also determined the formation of costalized wings.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXXIIa.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII
Back to Evolutionary Part LXVIII
Back to Evolutionary Part LXIX
Back to Evolutionary Part LXXa
Back to Evolutionary Part LXXb
Back to Evolutionary Part LXXI
Back to Evolutionary Part LXXI
{kind=link}