
[Also in these enquiries into the wing-venation and its transformations in Diptera-Nematocera we will consider the subcostal vein (Sc) at most only occasionally. This vein, especially its terminal branches (Sc1, Sc2), is -- in Diptera -- often difficult to discern.]
Here we will consider in more detail the parallel transformational pathways leading from a protorhyphoid wing-venation, via the rhagionoid and empidoid stages, to the various muscoid venational patterns. Before we do so, however, we first expound the general theoretical background against which the venational transformations are projected.
The theory of morphoklines and the polyphyletic origin of groups and subgroups of Diptera.
A m o r p h o k l i n e is a more or less continuous series of morphological stages of a chosen organ-system (in our case wings) leading from one stage, via a series of intermediate stages, to a chosen 'end stage'. We will diagrammatize such a morphokline by a vertical line directed upwards. Certain existing (or hypothetical) wing-venations (venational patterns) are represented on such a line (morphokline) by points or nodes. The wing-venation in groups of Diptera is supposed to transform and develop along a bundle of parallel lines or morphoklines. Here we give a general scheme of such a bundle :
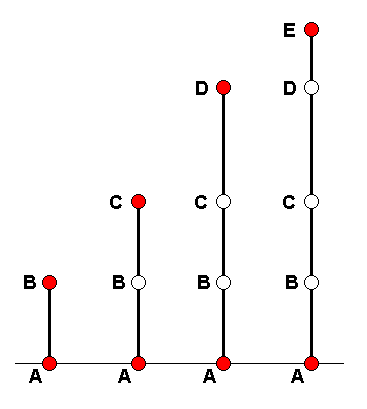
Diagram above :
A = starting venation, represented by the venation of an existing (or hypothetic) dipterous species or higher taxon.
B = second transformational stage. C = third transformational stage. D = fourth transformational stage. E = final transformational stage. The A's in all the lines represent identical or highly similar venations. The same applies to B, C, D, and E.
Vertical lines from left to right :
First line = morphokline of the wing-venation of B, where B is the venation of the wings of some existing species or higher taxon. Here the taxon represented by B has actually -- that is, historically - descended from the taxon represented by A.
Second line = morphokline of the wing-venation of C, where C is the venation of the wings of some existing species or higher taxon. The transformation has proceeded from A to C via the intermediate stage B. The taxon represented by C in this second line has not actually descended from the taxon represented by B in the first line. Only its venation (and not necessarily all other characters) has passed through the venational stage represented by B.
Third line = morphokline of the wing-venation of D, where D is the venation of the wings of some existing species or higher taxon. The transformation has proceeded from A to D via the intermediate stages B and C. The taxon represented by D in this third line has not actually descended from the taxon represented by C in the second line. Only its venation has passed through the venational stage represented by C.
Fourth line = morphokline of the wing-venation of E, where E is the venation of the wings of some existing species or higher taxon. The transformation has proceeded from A to E via the intermediate stages B, C, and D. The taxon represented by E in this fourth line has not actually descended from the taxon represented by D in the third line. Only its venation has passed through the venational stage represented by D.
As a result of the actual transformations having taken place separately and independently along the four lines, the wing-venation in the taxon E can formally be derived from that in the taxon D (third line), and this in turn from that in taxon C (second line), and this from B (first line), and B from A. However, this does not necessarily mean that the taxa themselves can be so derived, let alone that these taxa have actually, that is, historically, descended from each other. In almost all cases the representatives of such a group of taxa possess features -- other than wing-venation -- that do not allow us to assume that these taxa actually have evolved from one another.
The morphoklines of rhagionoid, empidoid, clythioid, and muscoid wing-venation.
We will now try to identify some such morphoklines that lead from a number of similar wing-venations as were present in the (fossil) nematocerous family Protorhyphidae to those (wing-venations) in Rhagionidae (Brachycera-orthorrapha), and then via rhagionoid venational stages to the venation as we see it in Empididae (still Brachycera-orthorrapha), and further to that in Clythiidae (Brachycera-cyclorrapha), and finally to the several types of wing-venation as we see it in muscoid flies (the house-fly and its --many and diverse -- allies) (all of them belonging to the large complex of Brachycera-cyclorrapha).
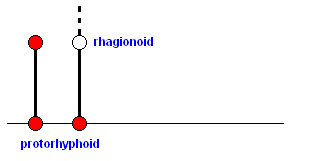
The protorhyphoid stage can be represented by the fossil Protorhyphus :

Figure 1 : Wing of Protorhyphus stigmaticus HANDL. Order Diptera, Family Protorhyphidae. Upper Lias of Mecklenburg, Germany.
(After HANDLIRSCH, 1938, from HENNIG, 1954)
The Radial Sector here is 3-branched. The Discoidal Cell is present. M3+4 splits up into M3 and M4, and the Cubitus (CuA [ = Cu1b] ) and first anal vein (1A [ = 1a] ) approach each other when running toward the wing-margin but do not meet : anal cell not closed.
In terms of morphoklines we can depict this as follows :

From here we pass over into the rhagionoid stage of wing-venation :

Figure 2 : Wing-venation of Rhagio (Asilomorpha-Rhagionidae) (Diptera). Recent.
1A = first anal vein (often denoted as A1, or An1 , or 1a).
2A = second anal vein (often denoted as A2, or An2 , or 2a).
R = radial system of veins. M = medial system of veins.
Cu1 (often denoted as CuA, or Cu1, or Cu1b) = anterior branch of cubital system of veins (the posterior branch [ Cu2 , CuP ] is in Diptera either a fold or is absent altogether).
(After RICHARDS & DAVIES, 1977, Imms' General Textbook of Entomology)
The Radial Sector is still 3-branched and the Discoidal Cell is (still) present. Medial, Cubital, and Anal system of veins still the same as in Protorhyphidae. Apart from a possible difference (evolutionary increase) of wing-size, the protothyphoid and rhagionoid wing-venations are identical. The (rest of the) morphology (and the ecology) of the respective imagines (fossil Protorhyphidae and recent Rhagionidae) (and of the larvae) can be assumed to be different.
In terms of morphoklines we can depict this as follows (where ". . . oid" signifies the venational type) :

The next rhagionoid wing still preserves a feature that points to a ( later [indeed having taken place] ) transistion to empidoid wings, that is, a transition to the empidoid stage of wing-venation :
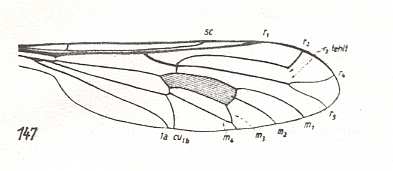
Figure 2a : Wing-venation of Lampromyia pallida MEIG. Recent. Order Diptera, Family Rhagionidae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
In this form we see the very first stage of the process that finally will result in the complete coalescence of M3 and M4, in turn (finally) resulting in M4, instead of M3, forming the posterior boundary of the Discoidal Cell.
In terms of morphoklines we can depict this as follows :
By having the upper point on the second line uncolored we indicate that, although the next venational patterns can be more or less derived from the venation as depicted in Figure 2a , this does not necessarily mean that a direct ancestor of the Rhagionid Lampromyia pallida (Figure 2a ) has actually given rise to members of the family Empididae. The uncolored point only represents a venational stage that has been passed through by species during their evolutionary transformation into species of the family Empididae. From this we pass over into the empidoid wing-venation, that is, to the empidoid stage of wing-venation :
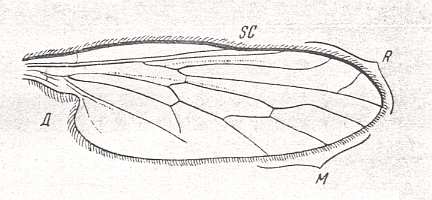
Figure 3 : Wing of Homalocnemis nigripennis PHIL. Recent. Order Diptera, Family Empididae, subfamily Brachystomatinae, Suborder Brachycera-orthorrapha. Length of wing approximately 5 mm.
(After COLLIN, from ROHDENDORF, 1951)
Here M3 has completely coalesced with M4. The latter now forms the posterior boundary of the Discoidal cell. While in the rhagionoid stage (of evolution of the wing-venation from protorhyphoid to muscoid) CuA ( = Cu1b) and 1A ( = 1a) approach each other while running to the wing-margin without actually joining up, or meet not until they are close to the wing-margin (as in Spaniopsis clelandi), in the present form the anal cell, formed in this way, is well under way of being contracted toward the wing-base as a result of a shortening of CuA.
In terms of morphoklines we can depict this as follows :
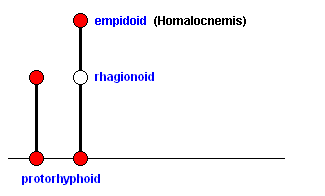
Further development of this venation consists in at least two lines (morphoklines) :
One of them leads to the advanced empidoid venation as we see it for example in Rhamphomyia :
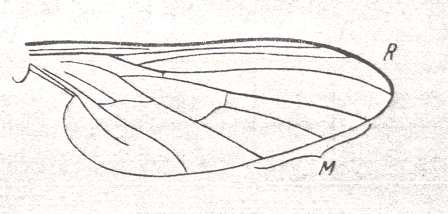
Figure 4 : Wing of male of Rhamphomyia platyptera PANZ. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha.
(After LINDNER, from ROHDENDORF, 1951)
Here (Figure 4) the Radial Sector has become two-branched and the contraction of the anal cell is in an advanced (though not final) stage. The structure of the Cubito-medial area in wings like those of Rhamphomyia and Leptopeza (see Figure 18 in previous document ) is already too advanced and idiosyncratic to give rise to the next stage in the evolution of the muscoid wing (which stage can be seen in Clythiidae), its morphokline (i.e. that of Rhamphomyia, Leptopeza, and close allies), remains within the confines of the Empididae (which means that such venation cannot develop into that of the Clythiidae, or into that as seen in other families in which the wing-venation (also) leads to that of the muscoid type of wings) :
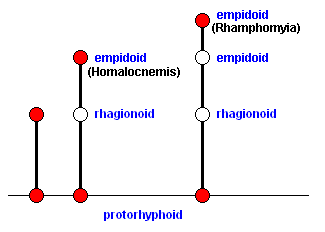
Only a wing-venation coresponding to a point on that morphokline on which the venation of Hybos culiciformis lies, but (a point) still lying somewhat before the latter, can lead to the first stage of muscoid wings -- the clythioid wings :
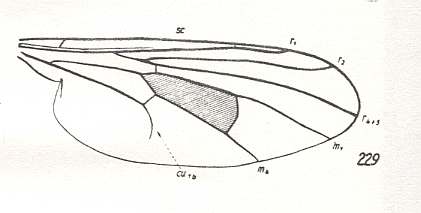
Figure 5 : Wing-venation of Hybos culiciformis FABR. Recent. Order Diptera, Family Empididae, Suborder Brachycera-orthorrapha. (After HENNIG, 1954)
As has been said, the venation of Hybos (like that of Rhamphomyia and that of Leptopeza) itself is too advanced in order to lie on the morphokline leading to the first stage of muscoid wings. It remains within the confines of the Empididae. But a little before Hybos, that is on some point lying before that of Hybos on the same morphokline, the venation (of this 'pro-hybos') is as follows : Radial Sector 2-branched, contraction of anal cell still in its early stages (first anal vein still present). The Media has become 2-branched. And as such it lies on the morphokline on which Callimyia lies, already belonging to the family Clythiidae ( = Platypezidae), that is, we enter the clythioid stage of wing-venation :
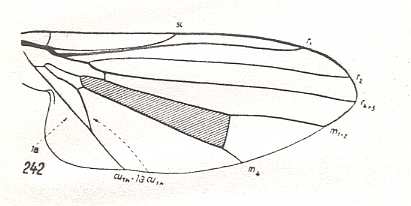
Figure 6 : Wing-venation of Callimyia speciosa MEIG. Recent. Order Diptera, Family Clythiidae ( = Platypezidae), Suborder Brachycera-cyclorrapha. (After HENNIG, 1954)
In this form (Figure 6) the Media is only 2-branched and the Discoidal Cell is still present.
In terms of morphoklines we can depict this as follows :
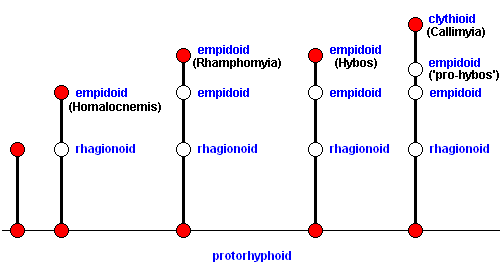
From a point lying still a bit before the above considered 'pro-hybos', a point representing a venation in which, like in Hybos (and in 'pro-hybos') the contraction of the anal cell is still in its early stages (were 1A present) and the Radial Sector is (already) 2-branched, but in which some elements (of the venation) are still in a more primitive stage : the presence of a long first anal vein (1A, 1a) (like it was already in 'pro-hybos'), and the Media still 3-branched, another clythioid venation can be derived. The venation represented by the mentioned point before that of 'pro-hybos' -- itself still empidoid -- must lie on the morphokline that leads also to the Clythiidae, but this time not to Callimyia, but to Clythia :
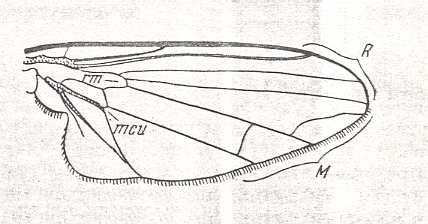
Figure 7 : Wing of male of Clythia picta MEIG. Recent. Order Diptera, Family Clythiidae, Suborder Brachycera-cyclorrapha. Specimen No. 2804. Length 4.5 mm.
(After ROHDENDORF, 1951)
In terms of morphoklines we can depict this as follows :
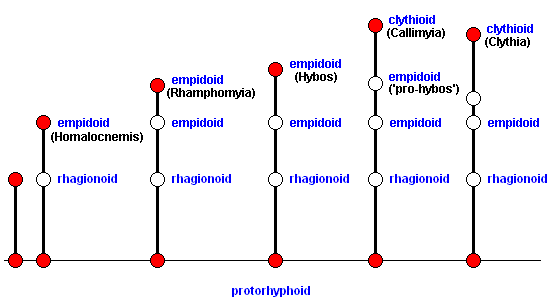
In order to have a general idea how such flies (Clythiidae) look like, we here produce an overall view of the insect :

Figure 8 : Clythia atra MEIG. General view of insect. Recent. Order Diptera, Family Clythiidae, Suborder Brachycera-cyclorrapha. Length of insect about 4 mm.
(After CHINERY, 1983, in Elseviers Insektengids, p. 236)
The clythioid wing-venation is already well within the general muscoid type of venation.
In the form represented in Figure 7 the Alula is (already) moderately developed and the Media is still 3-branched (M1, M2, M4), therefore it (i.e. that venation and wing-shape) lies definitely on the morphokline leading to the wing-venation and wing-shape (alula) as it is in the calyptrate flies (such as blow-flies, tachinids, the house-fly, etc.) -- the muscoid venation s.str. -- which (flies in their venation) all possess the more or less sharp characteristic bend of the (anterior part of the) Media :
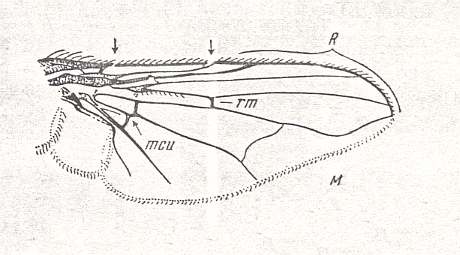
Figure 9 : Wing of Graphomyia maculata SCOP. Recent. Order Diptera, Family Muscidae, Suborder Brachycera-cyclorrapha.
Specimen No. 3770. Length of wing 9.5 mm. The arrows point to point-like interruptions of the costal vein which contribute to the elasticity of the wing.
(After ROHDENDORF, 1951)
In this form (Figure 9) the Alula is very strongly developed and the Medial system is derived from that in Figure 7 (Clythia) by having M2 reduced. It now consists of M4 and M1 only. The contraction of the anal cell is in an advanced state as compared to that in Clythia (see Figure 7 ).
In terms of morphoklines we can depict all this as follows :
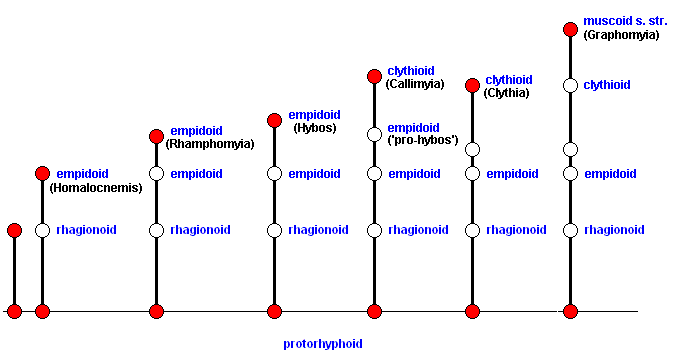
And also here it should be clear that all this should not mean that the genus Graphomyia has actually evolved from certain (anestral) representatives of the genus Clythia.
Placing the above findings in a more general and theoretical context.
The resulting final diagram above shows a parallel series of morphoklines. And a morphokline reflects a linear series of transformations historically assumed to have taken place in some particular chosen organ system, in the present case the wings of Diptera. The uncolored points on the morphoklines represent intermediate venational types. The red points represent actually living species of Diptera. The red point on the bottom of each morphokline represents some species of the family Protorhyphidae or of close relatives possessing a 'starting' wing-venation from which -- climbing up the morphokline -- other venational types have evolutionarily been derived. Each horizontal level in the (whole) diagram represents a certain stage in the development of muscoid wing-venation.
By working out some cases as they can be found in the mentioned diagram our understanding of a small segment of dipterous evolution can become still better (yielding at the same time some additional general understanding too) :
Looking at the wings of Rhamphomyia (Figure 4 ) and of Homalocnemis (Figure 3 ) we see that the 2-branched Rs of Rhamphomyia can be derived from the 3-branched Rs of Homalocnemis, so also the advanced state of contraction of the anal cell in Rhamphomyia from the more primitive state in Homalocnemis. However, it is more problematic to derive the absence of the Alula in Rhamphomyia from its beginning presence in Homalocnemis. Although such a derivation might be interpreted as a reduction of the incipient Alula, it is not very probable (to have happened historically), especially in the light of the path of evolution presently considered, a path leading to true muscoid wings, because all muscoid wings possess a strongly developed Alula. But although this is only a minor feature causing trouble to derive the wing-structure of Rhamphomyia from that of Homalocnemis, it is an indication that representatives of Rhamphomyia (that is, with all of their features, not only those of their wings) could not actually have evolved from (relatively recent ancestors of) Homalocnemis. And, what is important, experience has made us to expect the discovery of much more of such features (forbidding such a derivation) as soon as we take into account all other (morphological or physiological) structures of the respective imagines (and of the respective larvae). So we can expect that in the present case the one genus did not evolve from the other. Their venational details nevertheless preserve some features that help us to determine what transformations have actually taken place in the wings of the presently considered flies. And these details have thus helped to construct the mentioned diagram of morphoklines.
The representatives of the genus Rhamphomyia, to stick to the present example, although not actually having been evolutionarily developed from representatives of relatively recent ancestors of the genus Homalocnemis, have been developed from flies represented by the uncolored point lying just below it (i.e. below the red point representing Rhamphomyia) on the same morphokline. This point is drawn here on the same horizontal level as lies the red point representing true representatives of the genus Homalocnemis in the previous morphokline (see again the mentioned diagram of morphoklines ). The mentioned uncolored point just below the red point indicating the position of representatives of the genus Rhamphomyia, does represent a species (of Empididae) from which the representatives of Rhamphomyia have been evolved. But this species is not necessarily belonging to the genus Homalocnemis. The diagram here expresses the fact that the wing-venation of this unknown species is of the same type as that in Homalocnemis. And it expresses this fact by having it on the same horizontal level as that of the point representing actual representatives of Homalocnemis on the other morphokline.
In the same way -- see again the diagram of morphoklines -- the representatives of the genus Callimyia (Clythiidae, see Figure 6 ) did not actually evolve from (relatively recent ancestors of) the genus Hybos (Empididae, see Figure 5 ) or its close relatives, because the Subcosta in Hybos (and. presumably also in its close relatives) is already in a reduced state, while it is still fully developed in Callimyia. So the representatives of the genus Callimyia must have been developed from a species that came before Hybos (or at least before Hybos culiciformis), as shown in the diagram . There we have indicated that representatives of the genus Callimyia evolved from flies represented in the diagram by the uncolored point (dubbed "pro-hybos") just below the red point standing for Callimyia on the same morphokline. Although the species represented by this uncolored point ('pro-hybos') is supposed to have a wing-venation that is of the same type as that in Hybos, albeit a little more primitive, it will surely differ from Hybos (or 'pro-hybos' for that matter) in many other characters (of the adult or of the larva). Let this last example suffice.
According to the above considerations the vertical lines in our diagram not only represent morphoklines but also true genealogical lines. And they are parallel and genealogically independent of each other, even within smaller sections of the order Diptera (Even within the single family Empididae we find three such independent genealogical lines). Supposing such a polyphyletic development is forced upon us by the phenomenon of cross-specialization which is not only found here but almost everywhere in the system of organisms If we do not believe in the occurrence of reversed evolution (Dollow's Law, respected by all evolutionists) a given feature cannot have evolved from a formally more advanced state of it. It must have been either as such inherited from some ancestral species, or it must have been developed from a more primitive state of it as still present in some ancestral species. Consider two species A and B which are thought to have a common ancestor somewhere in the past. Now it is very common that the species A has a mix of formally primitive and advanced features -- primitive and advanced relative to the states of the corresponding features in the species B. A little reflection reveals that, depending on the degree of primitiveness of characters in A and in B, one has to locate the common ancestor of A and B very deeply back into history. And as characters in A and B are discovered of a still higher degree of (formal) primitiveness then one must project the common ancestor still further into the depths of history. As a result, the supposed ancestral species becomes more and more hypothetical and more and more ancient. The two lines originating from this supposed ancestral species then run for longer and longer times independently of each other. A natural conclusion, or at least a reasonable assumption, might be that following these lines from recent times into the depths of history they never meet at some common ancestor. The same can be said in cases involving three or more of such parallel lines. And this consideration, of course, also holds for our present case of fly-evolution. The morphologies and other characters at least suggest the type of evolution -- that is, polyphyletic evolution -- as expressed by our above diagram of morphoklines .
Evolution chiefly driven by random genetic mutations (+ natural selection) or by non-random genetic mutations?
The generally accepted framework of factors in organic evolution consists of random genetic mutations of which some, that turn out to be beneficial in a particular biological context, are favored by natural selection resulting in such a mutation to become dominant in that context while (locally) weeding out the original state of that part of the genome in which the mutation had taken place. This is standard Darwinian theory, that is, it is the theory of Darwin (1859) now updated with the many findings in cell-biology and genetics that came after him. The theory first of all was designed to give a natural explanation not only of the fact of organic evolution but also of the factors driving it. These factors are supposed to be unintelligent agents. And in fact the only way to figure this out is to assume randomness and natural selection, that is, organisms are supposed to constantly diversify themselves into (intraspecific) varieties in random ways and in this way gives natural selection something to go on. The resulting accumulation of selected small beneficial changes ultimately leads to the evolutionary emergence of complex organic systems and sophisticated and subtle adaptations -- so goes the theory. Indeed, any theory insisting in providing a material or purely physical explanation of the emergence of truly novel structures and patterns -- as we see it in organic evolution -- must come up with some sort of existing stochastic system, that is a system in which elements of the same sort are nevertheless naturally scattered over a certain range of variation, (and such a system then existing) together with some selecting mechanism. And in order for the explanation to be purely physical the selecting mechanism should not be some extrinsic agent (like it is, for instance, in the breeding and selecting of canine races) but be intrinsic to the system and its immediate conditions, that is, the selecting agent must reside within that system + its natural external conditions determined by that system itself. Said differently, such an explanation should not use the model of structured things selected or made by man because in such a model "man" is taken to be the extrinsic intelligent selector or designer. In the Darwinian theory of evolution the selecting agent is indeed not some extrinsic entity, but just a logical principle residing everywhere because it is no more than a tautology (it always holds) : survival of the fittest , which in fact merely means survival of the survivors. Such a selection agent will be active in all cases where there are entities whose enduring existence depends on the degree of their intrinsic robustness and resistance against detrimental conditions. The variations to which natural selection can fasten upon are the phenotypical expressions of genetic variations that originated as a result of random genetic mutations. Most genetic mutations are either not phenotypically expressed or, when expressed, harmful, but some of them, in their phenotypical expression, happen to be beneficial to its carriers in a given biological context. And because they are genetic they will be passed on to the next generation, and so on. During the course of successive generations these mutants (that is, the individuals carrying the beneficial genetic mutation) are going to represent the majority (of individuals) in the population in which the mutants appeared for the first time (and intially in small numbers). This is supposed to be followed by other random genetic mutations among which there are some that carry the initial beneficial phenotypic effect further until at last a new functional (phenotypic) structure has been formed giving the organism new (ecological) opportunities.
This Darwinian way of explaining organic evolution, an explanation, that is to say, in terms of random genetic mutations and natural selection (as the chief driving forces of evolution), plausible as it seems to be, is often challenged by alternative ways to explain the historical transformation and development of organisms (We do not discuss here the silly attempts to deny the fact itself of evolution on religious grounds). However, every genuinely physical explanation (using exclusively 'unintelligent' factors) of the appearance of true novelty must bring in random variation and natural selection. Random variation indeed creates novelty without being led by purpose, design, and foresight (which cannot, of course, figure in any physical explanation). But because it is random novelty, it can be of all sorts : beneficial, detrimental, insignificant, or irrelevant. So the presence of a selection mechanism must be assumed. And thus every physical explanation of the appearance of novelty must assume the enduring presence of random variation, and consequently all alternative explanations of (the factors of) evolution must involve non-physical factors (Such an 'explanation' is then in fact no explanation at all, because every genuine explanation of some given complex not-understood phenomenon reduces it to a more simple, familiar, and understood phenomenon, and such a phenomenon is always some material entity or process). But although random genetic mutation and natural selection as chief factors of organic evolution remain to represent the only genuine physical explanation (that is, an explanation in terms of physics and chemistry) of the fact and course of evolution because no alternative physical explantion can be given, it has seemed to me (and to some others) already for a long time highly improbable that random genetic mutation and natural selection of fittest mutants are the leading factors of organic evolution. It is not denied that they exist and play a role in evolution, it is only denied that they represent the main driving force of evolution. Some reasons for this denial are the following three points :
(1) It has turned out that there does not exist a straightforward and always-holding one-to-one correspondence between genes and visible (morphological, physiological, ethological) features in organisms. Genes code for proteins which are biochemical catalysts, building materials, or regulating entities (regulating the switching on or off of genes), but we don't know much about the supposed (long) chain that leads from genes or even proteins to macroscopic features of the organism. Most of these features certainly depend on certain genes and on the proteins the latter code for, but it is by far not certain that there are no other factors involved than genes and proteins in the determination of these macroscopic features. We know of organisms having the same genome but which are structurally very different. There may be factors lying outside the organism that co-determine many of its macroscopic and constant features, or even determine many of them without any genetic control whatsoever. In such cases genetic mutations (random or non-random) are simply irrelevant.
(2) All (morphological, physiological, behavioral) novelties that are associated with the evolutionary emergence of new types of organisms, are the result of highly complex, and seemingly designed, transformations of initial more or less simple states of organs or of the very 'beginnings' of them. To let this happen exclusively as a result of a series of successive random genetic mutations (and selection of the beneficial mutants) would take too much time, that is, would take much more time than is actually available in organic evolution on 'our' planet. On its way to the creation of the more or less final useful (morphological, physiological, or behavioral) structure (of some given organ) evolution will often get stuck in dead ends (i.e. ending up on local 'fitness peaks' in the evolutionary fitness landscape) long before the mentiond structure (situated on the global 'fitness peak' of the fitness landscape) would have been evolutionarily developed. And it will take unique mutations -- and thus much time -- for the organisms (meant here as evolutionary entities, species) to leave the local fitness peak and continue their evolution toward the global fitness peak.
(3) Contrary to general and wide-spread opinion, it is my belief that evolution is not about the creation of more and more fitted organisms : The fact that many so-called very primitive organisms are thriving extremely well still today (as they did in the past) (think of bacteria, coelenterates, many primitive worms, insects, etc.) supports this belief. Of course fitness -- in the sense of being well-adapted to the environment -- everywhere and always will constitute the final decisive factor in evolutionary changes, but that is not some sort of general multi-purpose fitness for life as such, but an appropriate and stable adaptation to a given particular environment in which a specific type of organism lives, while another type is living in another particular enviropnment. Indeed it is the fitness to live in a newly colonized environment that counts. And here we must concentrate our attempts to understand the nature of organic evolution not on this "fitness to live in . . ." -- because this is evident and tautological -- but on the "newly colonized . . . . Only here we find the general fitness being created, that is the fitness of life to be life at all : The surface of our planet has suffered many changes, climatological and geological. Many of such changes have wiped out whole populations, species, or even types of organisms. But the fact that life still flourishes almost everywhere on earth demonstrates that its general nature is such that it -- as a whole -- can survive even almost global catastrophes (such as significant climatic changes [drought, freezing conditions, heat] in extensive areas of the globe, big changes in land-sea relations, meteor impacts, etc.). And this it does by being extremely diverse, that is, by having colonized virtually every kind of environment in the large and in the small, and thus by splitting itself up into a myriad of different groups each specially adapted to a particular corner of the (overall) environment. So life - at least in the form as we know it from our earth -- must be such that it intrinsically seeks to diversify itself as much as possible, which here means not to create merely better organisms but to colonize as great a diversity of environmental types as possible by creating a corresponding diversity in life itself. To become different is more important than to become more fitted to an existing environment. But, of course, successfully colonizing a new environment, or a new corner of an existing environment, implies a developed fitness to live in this environment. Nevertheless we can say that Darwinian evolutionary theory is too one-sidedly engaged with the fitness of organisms and neglects the evolutionary importance of their diversity. When, on the other hand, the latter is seen as the most significant phenomenon of evolution the notion of natural selection becomes less significant.
So these three points always had convinced us that random mutation in combination with natural selection ('Darwinian evolution') cannot constitute the essence of organic evolution as it has taken place on our planet.
But be these points as they are, they could nevertheless be valuated as constituting no more than merely qualitative and speculative objections against Darwinian evolution. However, they could have been so evaluated only until recently : Genetic mutation is a change in the genome of some given organism, a change passed on to its descendants, and, depending on the phenotypic effect it has in the life of its carriers, being selected in favor of the original structure of the genome in some given biological context. Such a mutation can consist in the replacement or deletion of a single nucleotide by another in an animal's DNA, or in the replacement or deletion of two, three, or a complete series of nucleotides, or of a whole genetic unit (a gene). This is first of all reflected in a chemical change in the corresponding protein coded for by that segment of the changed genome. And because proteins are (1) either enzymes (catalysts) for important biochemical reactions, or (2) are specific building materials of the organisms, or (3) are, often in conjunction with some other proteins, specific cellular machines performing certain fundamental operations in the cell, such genetic mutations can have profound effects in the working of an organism, i.e. effects on its fundamental machinery. Most genetic mutations are supposed to be random mutations having taken place somewhere in the genome of a given organism, and they, i.e. random mutations -- where they are not noxious or even lethal -- are supposed to provide for a store of potential novelty eventually to be expressed in evolution.
But despite the fact that explaining the evolutionary appearance of organic novelty needs randomness (and selection), the chief role being allegedly played by random mutation + natural selection in organic evolution has been recently emphatically denied, not simply by considering merely the above three points, but by precise statistic reasonings concerning the detailed features of the genome and the proteins coded by it, including all the uncovered molecular control systems, responsible for the correct working of the organic cell. It has turned out that the limits of Darwinian explanation of evolution (explanation by random genetic mutation + natural selection) lie already in the molecular processes of the living cell (and not solely in macroscopic features of organisms) : Random mutation and subsequent natural selection cannot account for the origin and transformation of even these molecular processes (only for a very few of them when they are sufficiently simple). The odds are too much against such origins and transformations to ever take place. Without going into (molecular) details we note that the arguments for this contention can be read in the book The edge of Evolution (meant is the edge of Darwinian evolution), 2007, by Michael BEHE, who evidently is an expert on molecular and genetic processes in organic cells. BEHE argues not against the fact itself of organic evolution but against the widely accepted notion that the genuine driving force of organic evolution is constituted by random genetic mutation and natural selection, while not denying the frequent occurrence of random genetic mutations and the presence of natural selection of mutants. Random genetic mutations and subsequent natural selection of certain mutants can only bring about some minor evolutionary changes, and because of this randomness the accumulation of appropriate mutations eventually leading to significant evolutionary changes is out of the question. BEHE therefore argues that the genetic mutations responsible for most of evolution must be non-random genetic mutations. This case is, in the book, well-argued, and certainly convincing. But then BEHE goes on to determine what it exactly means to say that evolution is driven by non-random genetic mutation. He holds that the concept of change by way of non-random mutation is equivalent to "intellegent design" without explaining, however, what this latter should mean. Have we to do here with a scientist who has become desperate during his long quest for the true factors of organic evolution and finally falls into the trap of religion (which tries to 'explain' things (away) by simply having people to believe in the existence of a transcendent creator and ruler a of the World), or (have we to do) with a scientist who, on the basis of his findings and those of many others, should think (once again) about the very nature of Reality-as-a-whole, but, lacking the philosophical power and knowledge (of ontology and metaphysics), isn't able to do so? I don't know. BEHE sticks with the conclusion "intelligent design" and doesn't add anything to that, except a little hint that the intellegent designer is not necessarily the benevolent God of most religions, because he also created biological entities like the parasite causing malaria in humans (including innocent children) and the AIDS-causing virus. And, moreover, the argument leading from the existence of highly complicated and functional machinery (or grand pieces of art for that matter) as we indeed find them in living organisms, to the existence of some intelligent designer -- a God -- is an old one : In the eighteenth century it was given by Paley. It's true, the argument leading to non-random mutations (instead of random mutations) responsible for the origin and transformation of structure already at the cellular level of life is sound, quantitative, and (therefore) convincing. But the conceptual transition from non-random changes to intelligent design is not. I thought that using the analogy with man-made machines to explain the organic world had been (rightly so) abandoned already for a long long time (especially since Darwin) !
So we must continue where BEHE left sound reasoning, that is, we must try to understand what "non-random" in "non-random mutation" should actually and truly mean (Of course in a specifically human context a non-random change is a change performed on the basis of intelligent decision or design, so here it has acquired a special meaning). To answer this question we must first know what "random (genetic) mutation" is. Here, that is, in the present context, the term "random" is not supposed to express that there is no knowledge of the cause (of that what is random), that is, that we don't know what the cause of the mutation is. The term also is not intended to express that there is no cause.
The term "random" in the expression "random (genetic) mutation" expresses the very uniqueness of the cause (of the mutation). Not every cause is necessarily unique, but some are. Such a (unique) cause is probably tied up with the total collocation of things and events in the whole world (or in a large part of it) at a single given point in time (At another, even at the next or previous) point in time in the history of the Universe, this total collocation is different). And such a collocation (how it precisely is at a given point of time) itself is undoubtedly unique. And a unique collocation generates unique causes, among which the cause of a given genetic mutation. Such a mutation is then called a "random" mutation.
Having said this, what then is a non-random (genetic) mutation? Let's try to answer this question. Like a random mutation also a non-random mutation has a definite cause. But the general nature of the cause is different from that of the random mutation. It is not unique. And this here means that it is such that it can in principle be repeated (that is, take place again) because it is not dependent on the unique total collocation of things and events in the whole world at a given point in time. And when the cause is not unique so is its effect -- the (genetic) mutation -- not unique, which means that we can expect such a mutation to take place more than once in the course of time. In a human context a mutation -- which here is, of course, not a genetic mutation but just some change of a given initial state of some limited and definite collocation of things into a different collocation (for example when something is purposefully designed and constructed) -- can be repeated by re-creating the determining conditions. And only here the result of non-random change can be equated with the result of "intelligent design". In a natural and general context, however, we should equate the result of non-random change (genetic or otherwise) with the effect of a non-unique cause. And the effect is then also non-unique. Only an uncritical look at such an effect tempts us to invoke "intelligent design" and lets us forget that the correspondency between (1) purposeful design and construction of machines by humans and (2) the generation and transformation of biological devices and structures in organic evolution, is no more than an analogy until it is proved to be otherwise. However, the problem is far from solved. It cannot be denied that the nature of almost all organic structures and processes is very functional indeed. And of course only such structures have a chance to survive in the course of time. But how were they (evolutionarily) generated in the first place? We now know that the answer "random mutation" is wrong precisely because of this very functionality and specifity of such structures and processes, that is because of the narrow range for them to be functional in a given specific biological context. To rely on pure chance would demand too much time (than there is actually available) for such a structure ever to appear (in the right organism at the right time). Before we will adress this problem and its solution we will first turn to the implications of our discovery of the genetic mutations being non-unique.
A genetic mutation being non-unique means that its cause is non-unique. Such a cause could be, for instance, the presence of short-wave radiation. We do not think that is the right place to find such causes, because the genetic mutations in which we are interested here take part in specific evolutionary transformations in organisms resulting in the diversification of functions among organisms. This means that the (non-random) genetic mutations playing a part in evolution are very specific. And indeed, only then they can be functional. So the cause of a given genetic non-random mutation must be non-unique but nevertheless biologically functional. Well, a cause that can present itself on several different occasions, which might mean that it is present as a spatially and temporally extensive factor, and a cause that is at the same time functional (i.e. a cause that can take part in the creation or transformation of certain organic functions), what could that be? Expressed in general terms it could be the environment of the organisms in question. The environment of a given organic species "S", living in it, is itself specific and at the same time not necessarily unique. Like the individuals of the species "S" are not unique, but nevertheless specific, the environment, insofar as it is the environment of the species "S", is delimited by this species. Said differently, while the environment simply as being the "external world" is a broad segment of the bio- and abiosphere, the organic species "S" narrows it down to a very specific sector of it, which is precisely "its environment". The rest of the outside world contains some general necessary conditions not only for the species "S" but for all organic species. So the precise qualitative content of a given organic species "S", the latter's very essence or nature, delimits its specific environment or ecological niche. It collects, from the complete set of environmental elements, precisely (and only) those qualitative elements of the existing outside world that in some way constitute a qualitative extension of it, that is an extension of the nature of the species "S". Said differently, the nature of the species "S" picks out only those elements of its external world that are relevant to it, or form some sort of qualitative extension of it. The total collection of these chosen elements then constitutes the specific environment or (equivalently) the ecological niche of the species "S".
In the same way in which the actual specific environment or ecological niche of a given organic species is delimited by the qualitative nature of that species, also a p o t e n t i a l ecological niche of this same species is so delimited. A potential ecological niche of a given species is, like its actual niche, also an extended qualitative sector of the external world, in which, however, the species cannot as such live, but could live after having acquired one or two additional adaptations. As long as such a qualitative sector of the external world does not exist, the species does not have a potential ecological niche but only an actual one, namely the one in which it resides and to which it is fully adapted. And like the actual ecological niche of a given species is a spatially and temporally extended qualitative sector of the existing external world, so also the potential ecological niche is such a sector. And if we assume the potential ecological niche of a given organic species to harbour in some way a possible cause of genetic mutations in individuals of this species, then we have our sought-for non-unique cause taking part in the formation of adaptations to new environments. How precisely a potential ecological niche can cause a genetic mutation will be discussed later (on the basis of our metaphysical theory established earlier), where we discuss the functionality of the mutations. Here we will concentrate on the non-uniqueness of the cause of the genetic mutations, where this cause is the potential ecological niche. The point is that wherever and whenever this potential niche of a given species happens to exist the potential cause of a particular genetic mutation (or a set of sequential causes of respective mutations) is present. And such a genetic mutation could spread among the members of the population and is then partaking in the development of an adaptation to the potential niche. And in this way the potential ecological niche of a given species changes into the actual ecological niche of a new species. And because of the fact that a potential ecological niche is a spatially (and temporally) extended section of the general environment (the external world) it can be a cause repeatedly on several different occasions, that is, causing the same or at least similar genetic change independently in several different geographic locations or at several points in history.
Like an organic species, an ecological niche as such is a more or less abstract entity, but it can occur concretely in the form of several individual instances throughout the overall environment. Let us diagrammize all this.
If necessary, increase the brightness of your monitor in order to obtain well-distinct colors in the diagrams that follow.
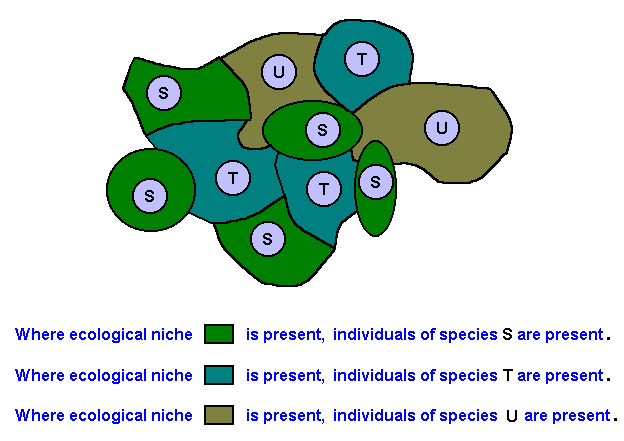
Of course it is possible that one or more individual instances of a given ecological niche happen not (yet) to be colonized by its corresponding organic species, but they can in principle become so colonized at any time.
The next diagram expresses the supposed fact that species S has, in addition to its actual ecological niche (green), a potential ecological niche (individual instances [orange] of which might be scattered throughout the overall environment) :
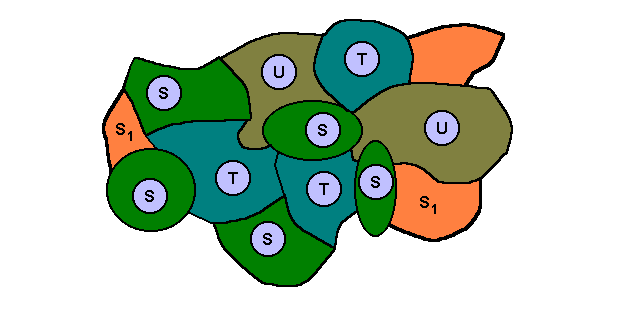
This potential ecological niche can become the actual ecological niche of a new species S1, that is, new as a result of the transformation (acquisition of an adaptation to this new niche) of the invading species S. But this will only happen when individual instances of this new niche are accessible at all by individuals of the species S. In the diagram this accessibility is indicated by the patches (orange) -- of the overall environment (patches) representing the potential ecological niche -- to actually being adjacent to the actual ecological niche of the species. In places where this is not the case the relevant individuals of the species are not supposed to be able to invade the potential niche.
[We here are describing things -- occupation of new ecological niches, the appearance of adaptations to these niches, etc. -- as we 'see' them (happening) in the Explicate Order, the Order that is, where things are unfolded along the space and time dimensions. And for the time being we will continue to do so.
Earlier -- Part IV of the Nomological Theory -- we have argued that these things we 'see' (observe) are in fact the result of projections of immaterial noëtic (that is, thought-like) reactions and their products from the Implicate Order (where they originally take place) into the Explicate Order. By having this -- noëtic reactions, Implicate Order, projection (and injection) -- in the back of our mind we can allow ourselves happily to speak about factors of the new ecological niche evolutionarily causing an adaptive change in the members of the species which is invading this new ecological niche. We can say this without us having to worry about the problematic issue of the change of parts of the genome by the action of non-random environmental factors, because our ultimate description will be in terms of noëtic reactions and the Implicate Order.]
Let's now apply all this to the evolution of muscoid flies as it was depicted in the above diagram of morphoklines . This is supposed to have been an evolution along parallel lines, where each line started off from some different member of the family Protorhyphidae, and where some lines got stuck, being not able to evolutionarily further develop, while other lines continued, finally producing the muscoid stock. We can reproduce this supposed evolution by setting up a consecutive series of diagrams in terms of species invading their potential niches :
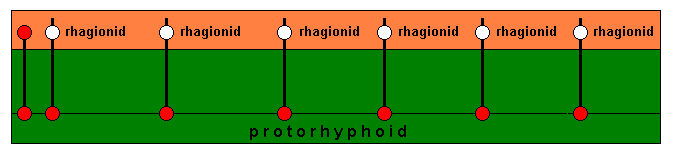
The green zone in the above diagram is the actual 'ecological niche' (so to speak for a moment) of the Protorhyphidae. We can call it the actual ecological zone of them. In fact this zone is subdivided into a number of slightly different subzones, which we call ecological niches, each corresponding to a different species of Protorhyphidae, that is, because these species differ a little from each other their respective ecological niches also differ. In addition to the actual ecological niche each species also delimits a potential ecological niche, which is different from the actual niche. These potential niches (one for every species) collectively form the potential ecological zone of the Protorhyphidae (orange zone in the diagram).
[In the case of the Protorhyphidae we do not here consider cases of certain existing species that cannot delimit a potential ecological niche. Such species do, of course, each for themselves, d e f i n e a potential niche, but because one or more elements of this defined niche are lacking in the existing overall environment it is impossible for such a species to d e l i m i t a potential niche].
In the course of evolution this potential ecological zone (orange) is invaded by members of the (at the time existing) Protorhyphidae.
[For a number of individuals of certain such species of Protorhyphidae no individual instances of their potential ecological niche can actually be reached by them (because of, say, geographical barriers). So these individuals will not participate in the invasion and colonization of (individual instances of) this potential ecological niche, and, consequently, will not develop an adaptation to this niche. At least at this point in history they remain unchanged (that is, they keep belonging to the original species). Other individuals of such a species do manage to reach and gradually colonize one or more individual instances of the species' potential ecological niche. They will evolutionarily develop the necessary (and initially lacking) adaptations to the potential ecological niche. They thus change and become a new species. The potential ecological niche of the original species becomes the actual ecological niche of the new species.
Based on such considerations we can assume that in many cases of such a transgression into a new ecological niche this transgression does not only result in the appearance of a new species but also in the preservation of the original species (represented by those individuals that have not been able to reach any individual instance of the potential niche). We do not, however, consider the latter (phenomenon) any further in our exposition.]
So, as has been said, in the course of evolution the potential ecological zone (orange) (consisting of the slightly different individual potential ecological niches of the respective species) is invaded by members of the Protorhyphidae. See diagram above . The latter thereby develop the necessary adaptations to this ecological zone causing each one of them to become a new species. These new species are supposed not to belong anymore to the (morphologically and ecologically defined) protorhyphoid type, but to the (morphologically and ecologically defined) rhagionid type, see next diagram :
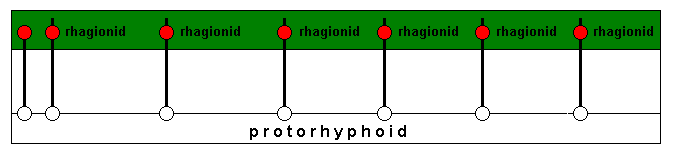
and compare it with the previous situation depicted earlier in the diagram above . We call it "rhagionid type" because all these new species have identical or similar wing-venation, a wing-venation as we see it for example in Rhagio, see Figure 2, above .
As soon as the new species have been formed and live in their respective actual ecological niches (and all of them, taken together as the 'rhagionid type' -- which [type] strictly is only about the wing-venation -- living in the common actual ecological zone (green) ), they delimit each for themselves a potential ecological niche, and as a collective a potential ecological zone (orange) :
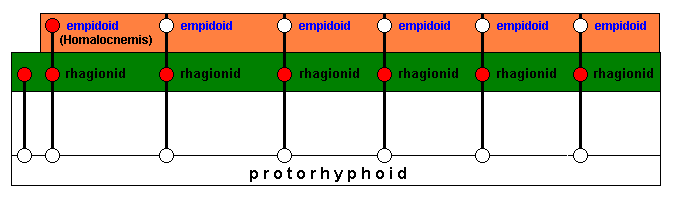
The species indicated at the extreme left is supposed to be a 'rhagionid' species that cannot delimit a potential ecological niche. Consequently this particular species remains the same while the other 'rhagionid' species will change in the course of evolution as soon as they have successfully invaded their respective potential ecological niches (and as a whole the potential ecological zone) and have acquired the necessary adaptational changes :
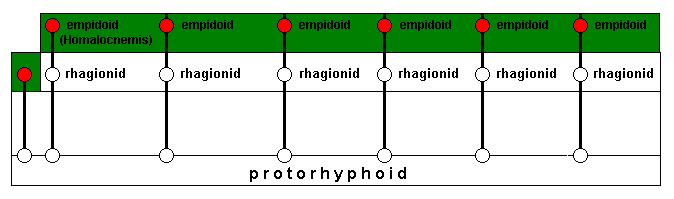
As soon as the new ecological zone is conquered, which means adapted to, by the rhagionid invaders, having those invaders changed into empidoids, they each for themselves (or most of them) delimit a new potential ecological niche (while collectively delimiting a new potential ecological zone (orange)) :
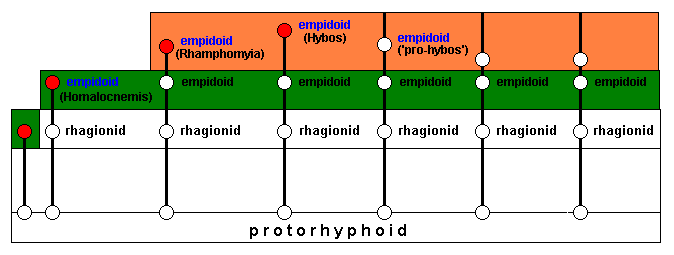
The line having resulted in Homalocnemis is such that it cannot delimit a potential ecological niche, because some necessary environmental factors are absent.
Invasion and successful adaptation of the new ecological zone (next diagram) results in representatives of the genera Rhamphomyia and Hybos (both still belonging to the family Empididae) which do not delimit a potential ecological niche. Other lines, while also still giving empidoids, do define, each for theselves, a potential ecological niche while collectively a potential ecological zone (orange) :
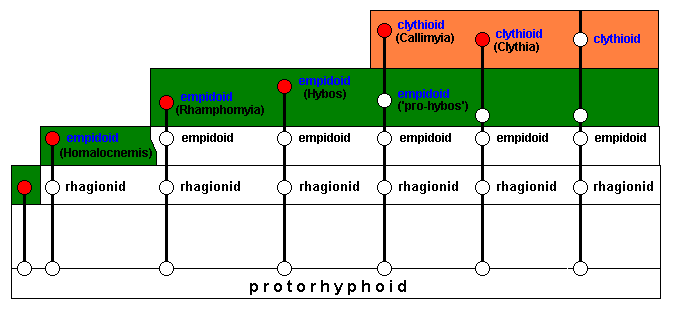
The new ecological zone will be invaded (next diagram), resulting in the transformation of empidoid into clythioid species. Among these latter are the genera Callimyia and Clythia (two lines). These two genera do not delimit a potential ecological niche, and do not, accordingly, develop any further evolutionarily. The last line, on the other hand, does result in a clythioid species that does delimit a potential ecological niche for itself (orange) :
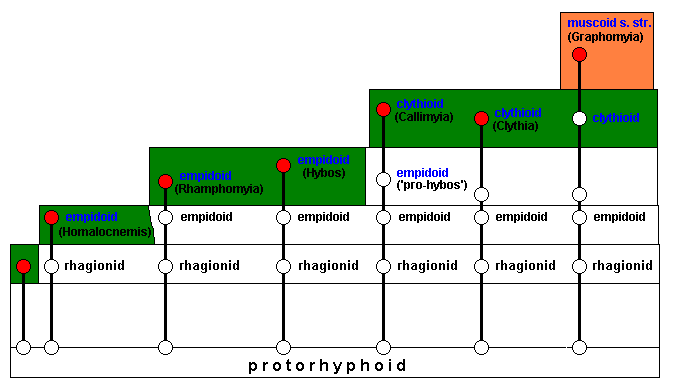
After invasion of this potential ecological niche the muscoid genus Graphomyia has been formed, with its members carrying the typical muscoid (s.str.) wing-venation :
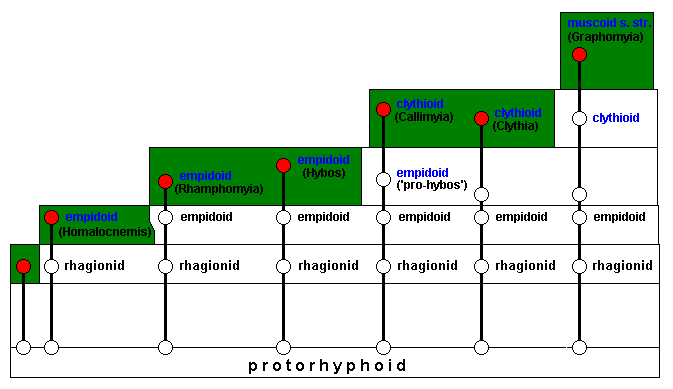
So now we have produced our diagram of morphoklines in terms of the successive invasions of a number of ecological zones by members of a bundle of several evolutionary lines, leading from protorhyphoids, through rhagionids, empidoids, and clythioids, to muscoids s.str.
Ecological niches, ecological zones and the evolution of the muscoid flies from protorhyphoid flies.
It is perhaps helpful to once again expound the concepts of (a) actual or potential ecological niche of a given species, (b) actual or potential ecological zone of a given higher taxon, (c) adaptation of the individuals of a given species to its ecological niche, (d) the colonization of new (potential) niches, (e) the formation of new species, etc., that is, helpful (as these things are) to actually begin to understand the chief factors of organic evolution.
What determines the course of evolution? Generally there are two chief factors, namely (1) the special qualitative content of the taxon (species, genus, family, etc.) and (2) the environment. And so it is also in the present case of the evolution leading from protorhyphoid to muscoid flies. Colonization of new habitats -- here nutritive media or substrates -- leads to the formation of new forms (because they develop new adaptations). But members of one or another given [higher] taxon cannot fan out to every kind whatsoever of new habitat. They can only start to colonize, and adapt to, a habitat that is, or is part of, the potential ecological zone of the taxon. So members (species) of any given taxon cannot explore and successfully colonize every kind of ecological zone whatsoever (which is meant to mean that the taxon cannot explore from its base, that is, from its proper and regular ecological zone, any arbitrary new zone). And for any given individual of this species (of that [higher] taxon) the new habitat [in the newly invaded ecological zone] must be one of the species' potential ecological niches. And for such a niche or such a zone to be a potential niche or zone for the taxon depends on the qualitative content of that taxon. In all this we can confidently assume that a given ecological niche (in the strictest and narrowest sense) can harbor only one species. This is just a matter of definition of the concept "ecological niche". ( Later we will meet cases where an apparent single particular ecological niche -- as always, being not a concrete habitat but a precisely delimited t y p e of habitat -- has been successfully occupied by two different, even taxonomically widely separated, species. But also in such cases there must exist, albeit subtle, differences.). An ecological zone, on the other hand, can harbor many species, depending on how many ecological niches it contains. While the concept "ecological niche" can, at least in principle, be precisely defined, the concept of "ecological zone" will -- apart from it having to accommodate more than one ecological niches -- remain more or less vague (some "zones" might be sub-zones, others might be super-zones).
Any particular given species possesses its actual ecological niche, the niche in which it lives. The precise qualitative content ( = special collection of environmental factors) of this actual ecological niche depends on the qualitative content of the species, and is made up of those parts or factors of the (overall) environment with which the individuals of the species are fully integrated, which here means that 'copies' of these environmental parts or factors are, as it were 'internalized' into the body of these individuals. These internalized environmental parts or factors are of course the special adaptations evolutionarily acquired by the individuals of the species.
In addition to this actual ecological niche such a given species also possesses its potential ecological niche (or even more of such niches). No individuals of the species actually live in such a niche, but after having evolutionarily acquired certain adaptations, provided the qualitative content of the species is such that it can indeed produce these adaptations, they can live in it. But because of now possessing these adaptations they have changed into a new species.
Ecological niches and ecological zones are present in the form of one or more actually existing individual instances of such a niche or zone.
A series of ecological niches, potential or actual, present in a particular type of environment, forms an ecological zone. An example could be the muddy bottom of fresh water basins. The muddy bottom of an existing particular water basin is an individual instance of that ecological zone. Another example is the shoreline of such a water basin ( The shoreline of an existing particular water basin is an individual instance of that ecological zone).
Generally, the majority of the smaller taxa (for example species) of a higher taxon (for example a family) lives in the actual ecological zone of that taxon, and each species of it lives in that species' own particular actual ecological niche, these niches together composing that ecological zone. This actual ecological zone, insofar as being the ecological zone of the taxon, only contains ecological niches in which species of that particular taxon live. But the ecological zone as such (not specifically that of a given [higher] taxon) contains, in addition to the niches mentioned, also niches of species of other [higher] taxa. So such an ecological zone can harbor species of different, say, families. How many different families have representatives living in this zone depends on the (relevant) qualitative content of this zone and on how broadly this zone is taken (for example 'woodland' is broader than 'oak forest').
In the same way the potential ecological zone of a given higher taxon (such as a family), only insofar as it is a potential zone of this particular taxon, only contains potential ecological niches of species of precisely that taxon. But understood just as ecological zone it can and will contain potential ecological niches of species of several different [higher] taxa.
An ecological niche of a given species, is, as has been said, precisely that particular set of environmental factors that are internalized into the body (in the broad sense, that is, including behavior) of the representatives of that species. For a large part the essence of the ecological niche is biochemical : the individuals of the species are biochemically well-matched to their direct nutritive substrate. But this essence is also made up by fine-tuned behavioral and, especially, fine-tuned morphological features. Because of lack of proper data we cannot as yet discuss evolution in terms of ecological niches (as defined here). We therefore concentrate on ecological zones (which contain a definite set of ecological niches). And because the ecology of Diptera is largely determined by their larval stage the term "ecological zone" of a given dipterous species refers to the living conditions of the larvae.
At first sight we could think of, say, "woodland", "oak forest", "rivers", "stagnant water-basins", "banks of rivers or of stagnant water-basins", "soil", etc. as representing ecological zones. [For a more general account of environments and nutritive substrates of dipterous larvae, see Organic Evolution, Part V, Intermezzo : Biology of the larvae of Diptera]. It is, however, more appropriate to see these 'zones' not as ecological zones but as general environments that can harbor true ecological zones. These true ecological zones are -- in the case of dipterous larvae -- in fact nutritive substrates. And as such these substrates determine classes of (larval) feeding habits. For the concise and provisional exposition of the evolution of muscoid flies from the ancient protorhyphoid flies the following larval feeding habits (determined by the respective nutritive substrates -- ecological zones) are important :
Zone of saprophagia
The larvae of the family Rhyphidae are polyphagous and rapidly developing. The larvae live in extensive moist substrates rich in decaying vegetable substances, substrates that, although widely distributed, are relatively less permanent (shoreline of streams or the soil of temporary water-basins in tropical forests, etc.) [ROHDENDORF, 1964 ].
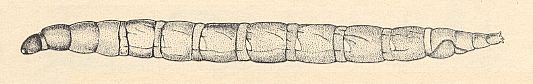
Figure 10 : Larva of Sylvicola fenestralis SCOP. ( Anisopodidae [ = Rhyphidae, Phryneidae] ).
(After KRIVOSJEINA, 1969)
Whether the larvae of the (Jurassic) Protorhyphidae lived this way we cannot, of course, know. There is nothing, however, that forbids us to assume that they were saprophagous in their larval stage.
Zone of zoophagia
The larvae of Empididae [Asilomorpha] are in their appearance and behavior less agressive (than those of Rhagionidae and Therevidae), but that does not mean that they do not hunt various invertebrates, especially earthworms. In the soil one can often encounter larvae, of which the intestine is filled with blood of these invertebrates. Generally, however, we are forced to say that the nature of feeding of many larvae of Empididae is unclear. [KRIVOSJEINA, 1969 ].
Generally, the imago is carnivorous. Many run about swiftly, and seize their prey (small insects, mites, spiders) with their legs. The larvae of this family (and also of the family Dolichopodidae) are essentially carnivorous, though they make do at times with carrion food, such as dead insects, and even with wholly vegetable material. Though the Empididae and the Dolichopodidae are essentially terrestrial, and are propery linked with the terrestrial Asiloidea, some members of both families have begun to stray towards the water. This may not be unexpected, since the larvae look for animal food, and have not found it in the parasitic or semi-parasitic ways of the Bombyliidae, Nemestrinidae, and Cyrtidae. Water has much to offer for carnivores. [OLDROYD, 1964 ].
Zone of mycetophagia
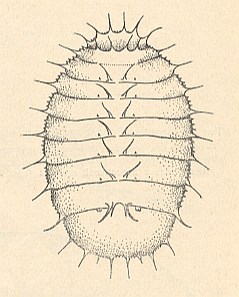
Figure 11 : General view of larva of Platypeza dorsalis MEIG. [Myiomorpha-Platypezidae].
(After KRIVOSJEINA, 1969)
So all this gives some impression of the ecological zones 'colonized' by the families that play a role in the above diagrams of morphoklines and successive colonizations of different ecological zones by representatives of a number of families. It does not permit to reconstruct the actual history of these families.
In the above discussions we have dealt with the ecological factors of evolution (that is, we have found out that THE factors of evolution are ecological). We have found them to lie in the environment (of the individuals of a given species whose role in evolution is being discussed). And, accordingly, we developed the concept of potential ecological niche as being a specific part or aspect of the environment of a given species. In some way such a potential ecological niche acts on (a part of) the (set of) individuals of that species (itself living in its actual ecological niche). How this can take place will be explained later on when dealing with the Explicate and Implicate Orders of Reality (next document).
In the next document we will once again describe the evolution of muscoid flies from (jurassic) protorhyphoid flies -- still, as compelled by necessity, in general terms only -- but now wholly in (general) terms of the Implicate/Explicate Orders and noëtic reactions.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXIII.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII