
In the present Phase the digger wasps (Sphecidae) play an important role. Therefore, before we expound this Phase, we will give two diagrams, one depicting the evolution of the Sphecinae (a subfamily of the Sphecidae), while the other depicting the kinds of arthropods utilized as prey by the representatives of the Sphecidae. The interpretation and data are from EVANS and EBERHARD, 1973. Further we give, from the same source, a schematic drawing of a more or less typical burrow (nest, cell) of a sphecoid wasp, but with a far from typical content.
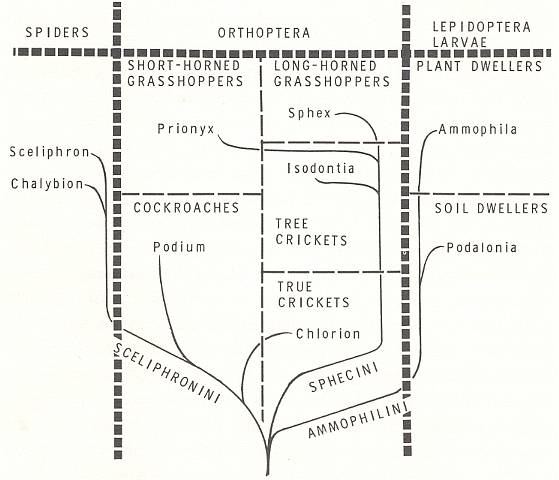
Relationships of the tribes and genera of the digger wasp subfamily Sphecinae. Although only prey type is indicated here, this arrangement is also supported by adult and larval structure, by type of nest, and by still other features. [For a magnification, click (twice) on the image]. (After EVANS and EBERHARD, 1973)
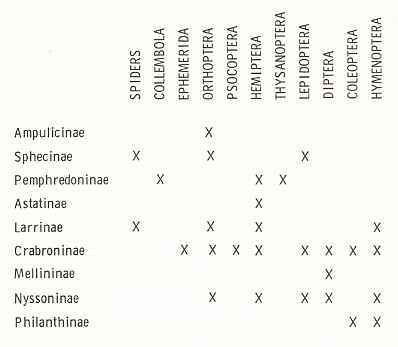
Table showing the subfamilies of digger wasps (Sphecidae) in the left column, and the kinds of arthropods utilized as prey (top). (After EVANS and EBERHARD, 1973)
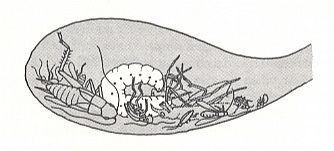
A larva of Microbembex monodonta (Sphecoidea) in the nest cell, feeding on a variety of dead arthropods (drawn from a photograph). (After EVANS and EBERHARD, 1973)
Assuming a new work-order -- first the construction of a nest accomodation, and then the hunt [for a prey] -- was an enormous step forward along the path of development of the maternal instincts of some wasps. Thus, the well-known digger wasp Ammophila sabulosa L., of the family Sphecidae, first digs out its characteristic burrow widened at its end, and [only] then (usually on the next day) brings to it a paralyzed caterpillar of a night owl. Here a number of illustrations :

Figure 1 : Ammophila sabulosa. 16-28 mm. Family Sphecidae.
(After SEVERA, in Thieme's Insektengids voor West- en Midden-Europa, 1977)
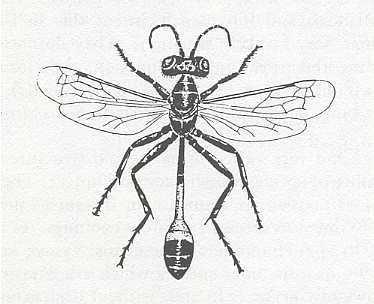
Figure 2 : Ammophila sabulosa. Female, ca 20 mm. Family Sphecidae.
(After RICHARDS and DAVIES, in Imms' General Textbook of Entomology, 1977)


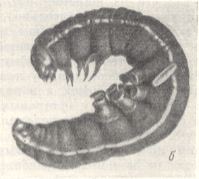
Figure 4 : Paralyzed caterpillar of a night owl (paralyzed by Ammophila ) lies at the bottom of the cell (Left image). It is removed from the cell (Right image).
(After MALYSHEV, 1966)
As we said, Ammophila sabulosa first digs out a nest, and only then catches a prey, paralyzes it, and brings it into the nest [With this change of work-order it is now effectively prevented that the prey will be stolen or infected by a foreign insect while the wasp is constructing a nest or searching for a proper place for it].
Primary-vespine (pompiloid) phase of development (discussed in the previous document).
A = Hunt :
a1 = Finding a prey.
a2 = Attacking the prey and paralyzing it.
a3 = Bring it to a certain place for it to stay temporarily.
B = Constructing or adapting a nest (= definite quarters of the prey) :
b1 = Finding an appropriate place for the nest.
b2 = Burrowing of the nest, adapting it, etc., usually in connection with visiting the prey.
C = Transporting the prey to, into, and in the nest.
c1 = Placing the prey before the nest.
c2 = Inspecting the nest.
c3 = Bringing the prey into the nest.
D = Laying of the egg.
E = Closure of the nest (from the outside).
e1 = Closure of the true cell.
e2 = Closure of the nest itself.
The original work order belonging to the previous Phase was :
And because this series is repeated a number of times (given by n ) by the wasp, because it makes several nests, each one supplied with a prey, the work-order is in fact serial :
or, equivalently :
In the new work-order the wasp, instead of first hunting the prey, first constructs a nest (thus reversing the acts A and B).
Let us explain (and, if necessary, see HERE for the meaning of the symbols).
As has been said, the wasp will generally provide for more than one descendant, meaning that several such series, that is originally : A + B + C + D + E, A1 + B1 + C1 + D1 + E1, A2 + B2 + C2 + D2 + E2, A3 + B3 + C3 + D3 + E3, etc. are executed. Said differently, the wasp will end up with a number nests each one of which provided with a paralyzed prey for a larva to feed on.
Now back to our new series B + A + C + D + E.
As regards B (construction of the nest) there is now no doubt that it is a residue of the first series (A + B + C + D + E). But from where was A (hunt) taken? And how must we interpret the actions C, D, and E ?
If the first act of the first series were indeed cancelled, then the insect could essentially not finish those acts connected with A, namely C (bringing the catch into the nest) and D (laying of the egg), when we take into account the presence of the instinct to lay-the-egg-on-the-prey. In such circumstances the whole original series would read (where "0" = zero) :
It is not hard to understand that the last act, E (closure of the nest), now becoming useless, also might be cancelled, and that the wasp now continues with the first act (A1) of the second series (A1 + B1 + C1 + D1 + E1), but now cancelling act B1 (constructing the second nest), which is now not necessary because a nest is already constructed (B).
So we obtain the following combination :
that is, precisely what we were looking for. From this it follows that the catched prey (A1) originally belonging to the second nest (built in the second series) is now placed by the wasp in the first nest. So in this way we have, not an exchange of the first acts (A and B), not a displacement in time of those acts, but just the combination of them out of two successive series ( B from the first series, while A1, C1, D1, and E1 from the second series) which is indeed expressed by our symbols. Therefore, A1 of the new series is not essentially that, what A is of the first series. (If necessary, see HERE for the meaning of the symbols).
That things went precisely that way, that is, by combining two successive series, is convincingly demonstrated by the Ammophila's. Thus, A. sabulosa L. and A. argentata Lep., after having first burrowed a nest, and having closed it while it is still empty, hunt down a prey, and then place it in the nest, lay their egg and finish their work as usual.
The above established series of acts can be further explained : Here we in fact have from the originally first series : 0 + B + 0 + 0 + E, and in addition to it from the second series : A1 + B1 + C1 + D1 + E1, where B1 -- the building act of the second series -- is here the removal of the temporary cap (the temporary closure of the nest was act E ). If we add up these two acts (E and B1) we get zero, and thus we indeed end up with our new series B + A1 + C1 + D1 + E1. And because here the hunt (for the prey) A1 and bringing it into the nest (act C1) are closely connected and not separated anymore by the construction of the nest, we can express this as :
In connection with this, it is not superfluous to note that Psammophila hirsuta Kby., still working under the original order, that is, first taking care of the prey, and only then constructing a burrow, does not make a temporary cap [of its nest] (see Figure 17 in previous document [Psammophila hirsuta and Ammophila hirsuta are probably the same species. The author-name of hirsuta is stated as "Scopoli" both in the Elseviers Insektengids as well as in MALYSHEV, 1966, p. 150, although on page 154 MALYSHEV writes "Kby." for this author-name (of hirsuta)] ).
It is interesting that temporary closure of the nest is sometimes also observed in other wasps that provide their cell with only a single prey. So do behave, for example, Sphex albisectus Lep. and Tachytes panzeri v.d. Lind., preparing grasshoppers [Orthoptera-Caelifera, family Pamphagidae].
However all this may be, it is totally clear that the cancellation of the first act (hunt) could not remain indifferent to the species and that it directly had to evoke a corresponding change of the whole course of the work, [the change] taking over to (as far as we can trace) the next series [originally connected with the second nest] first. In the present stage of development the plasticity of the vespine instincts was 'put to a hard test', but the ability to self-regulate (a similar correlation in the organism), as we already have told, allowed the instinct to overcome the difficulty and led it [the instinct] onto a broad path : All higher wasps, and all, without exception, independently nesting bees, first prepare a cell, and only then go to obtain provisions.
The new work-order in the present phase of evolutionary development, in those cases where into the preliminarily constructed cell is placed only one prey, is today observed only in a few, partly already mentioned, digger wasps (Sphecoidea), and as an exception also in [some] spider wasps (Pompilidae). In the remaining wasps this new work-order, as it is expressed in the present Phase, is either almost not at all expressed or its beginning stage has disappeared without a trace.
We will soon see why precisely the wasps in their great majority did not stick to the present Phase but developed further. Let us, for the moment, note that already in this Phase the new work-order was beneficently expressed in the development of building instincts in wasps. First of all, for them opened up the possibility to concentrate on building of nests in definite places, that gave birth to the habit to be present colonially, from one year to the next, at a well-selected site. And the very methods of construction-work became more subtle and complex. As a result, individual sections of the nest received a definite shape or form, and from time to time also became larger, whereby in one single nest it became possible to construct not just a mere one cell anymore, but several cells.
Thus, one of the largest wasps of North America -- Sphecius speciosus Dr. (Stizidae) digs burrows, reaching a depth of almost half a meter, of which each ends up with a spherical cell with a diameter of about 36 mm. In further work, at the end of the chief burrow, consecutively are dug out several side-channels each ending up with such a cell. See next Figure.
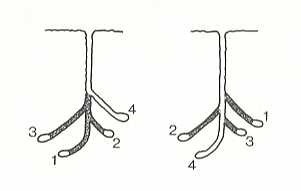
Figure 5 : Two major types of multicellular nests in digger wasps.
On the left, the initial cell is deepest and additional cells constructed from side-burrows back toward the entrance. On the right, the initial cell is closest to the entrance, and additional cells are added by deepening the burrow.
(After NIELSEN, 1933, in EVANS and EBERHARD, 1973)
Having constructed a cell, the wasp finds a large cicada, knocks it from the tree, and stings it into the thorax at the base of the forelegs. After that, having grabbed the prey firmly with the middle legs, the wasp climbs with it up into the tree in order to find a proper place to descend and lets itself, together with the paralyzed cicada, fall down in the direction of the entrance of the nest, to which it then drags the prey. The egg is deposited onto the thorax of the cicada, between its legs. See next Figure, depicting the transportation.

Although the established new work-order (preparing an accommodation for the prey before the hunt for it) lies at the base of the development of the higher forms of vespine life, among the present group of wasps still those primitive features were preserved which we saw in their ancestors. Thus, larvae of wasps, also here, as in the previous Phases, each develop at the expense of only one prey, and moreover usually in a unicellular nest. To this primitivity of the present group of wasps definitely points also the selection of preys by them. In the above given examples we again see orthoptera -- crickets, grasshoppers, and also cicadas -- as prey, that is, still insects with incomplete metamorphosis. To this choice are added caterpillars, especially those that live in the soil or above it close to its surface. But larvae of beetles we do not find here. And in the present Phase we do not find preys which are adult insects with complete metamorphosis -- beetles, butterflies, and flies. They did not figure as prey in the past, neither in the present.

As preys we here [in the present Phase] again encounter crickets (Gryllidae). Thus, the Australian wasp Diamma bicolor Westw. attacks the mole cricket Gryllotalpa coarctata Wek., paralyzes it, and then drags it to the burrow, prepared preliminarily. The paralysis is also here temporary, but longer lasting, taking an hour.
With all this we conclude our exposition of the Secondary-vespine (sphecoid) Phase of hymenopterous evolution.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part L.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII