
Introduction
Hennig's [1954] phylogeny of the Infraorder Bibionomorpha of the Order Diptera, as we have it (further) worked out in Part XXVIII, from the Section The cladistic evolution of the Bibionomorpha onwards and Part XXVIII-A , was (primarily) based on wing-venation. And, especially, it was based on formal derivations found to be possible between the various venations of existing species. But these formal derivational relationships mainly concerned the number of branches of the chief longitudinal veins, and the presence, absense, or transformation of the particular cross-veins as they are presumed to be present in the diptera-order-prototype (the cross-veins ta [= r-m], tp [= intermedial cross-vein], and tb [= tb2 = m-cu] ). On the other hand, differences as to the general 'look' of the wing (its absolute and relative size, its shape, the spatial position and distribution of the main veins and their branches, presence of hairs, bristles, or scales, etc.) were not considered (they have been considered when functional types were described). So although the derivations of the venational branchings were formal, the wings -- each one as a whole -- were not completely, and that is not strictly (formally) derived from one another (in order to establish phylogenetic relationships). Further, in the mentioned phylogenetic investigation of the Bibionomorpha, although, of course, species were considered, they only functioned there to represent families, that is, the wing-venation of the representatives of a particular species, or some of them, was taken to represent the venational characteristics of the family to which that species belongs. So in the mentioned phylogenetics of the Bibionomorpha wing-venations of families were derived from those of other families of the infraorder. And in such an approach it is to be expected that specialization-crossings will be encountered (and even more so when also features other than wing-venation are being involved to demonstrate phylogenetic kinship). So, in deriving one family-groundplan from another, there may exist, in the groundplan of the original family, a character whose state is more derived than that of the corresponding character in the groundplan of the to-be-derived family, violating Dollo's law of irreversibility. Thus, as a result, virtually not any single family of Bibiononorpha could directly be derived from some other family, but only from more or less remote ancestors of the latter in order to escape from specialization-crossing :
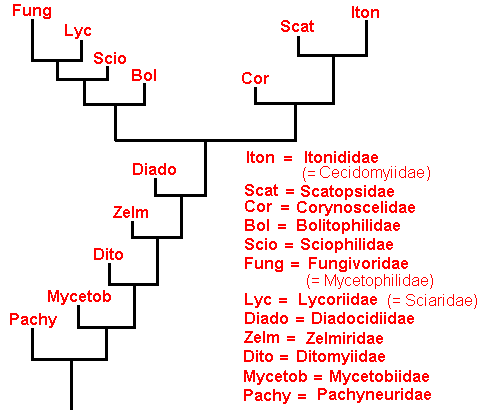
But because the more specialization-crossings will be discovered between any two compared families, the more backwards in history (and in our theory that is noëtic history) their point of phylogenetic separation (point of bifurcation) has to be pushed, -- it is safest to not acknowledge the actual existence of such a point anywhere in history, and hold, in contrast, that the two families have developed independently of each other. So if we have two families, X and Y, where Y can roughly be derived from X, we now say that Y has originated from a precursor of it [i.e. of Y] that looked like X. In Part LXIXb we have done this, not at the family level, but at the species level, while having (also) confined ourselves to wing-venation (And also in our phylogenetic investigations [Parts XXVIII and XXVIII-A] we have confined ourselves to wing-venation).
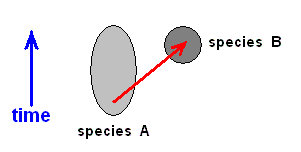
where the time difference can, phylogenetically, still be considered to be zero, meaning that A and B are contemporaneous.
And such a way of origination can only be assumed when species A and B belong at least to the same genus, and are, within it, most closely related morphologically. So also in the Implicate Order (where the derivation actually and timelessly took place) it must be assumed that the corresponding descriptions of the strategies (s.l.) must be so related to each other.
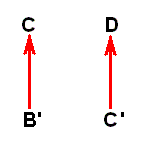
If, on the other hand, two species, C and D, belong to different genera (or even to different families), and D is (nevertheless) more or less derivable from C, then its derivation is more indirect, that is, more remote, than the derivation of B from A was. It cannot be contemporaneous (in the above sense) anymore. So, according to phylogenetic systematics species D must have been evolved from a more or less remote precursor of C. Accordingly, here we then have, not species D directly having evolved from species C, but from species C', meaning that we here have to do with an assumed splitting up of an original species C' into the species C and D :
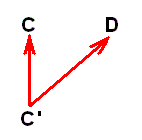
where -- it must be stressed -- species C' is different from species C, and that this species C' is to some extent hypothetical. And, moreover, now we have the possibility of specialization-crossing between species C and D, which may cause us to project the hypothetical common ancestor of these species farther back in time. So, as a result, it may, after all, be safest to assume -- contrary to phylogenetic systematics -- that species C and D ( [morphologically] belonging to different genera) each have their own historic background : Their ancestral lines independently (i.e. independent from each other) extend into the depths of history (and in our theory this history is noëtic history), so that species D has evolved, not from species C', but from D' :
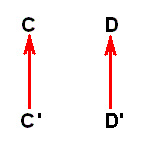
where D' is assumed to be more or less similar (but not identical) to C. And although all this is also hypothetical, it is less hypothetical than the assumption that D has evolved from C'. We assume species C and D having each their own bundle of morphoklines (one morphokline for venational transformations, another for, say, chaetotaxic transformations, etc.).
In our ealier diagrams ( Part LXIXb ) of venational morphoklines we have expressed the assumed similarity between (1) the independent precursor of D and (2) species C by means of related signs (C and C') :
In the earlier documents on Bibionomorpha the cladistic course of phylogeny, established by the methods of phylogenetic systematics, using primarily wing-venation, was -- in our theory -- interpreted as the course -- in the Implicate Order-- of the noëtic trajectory (which is a course of consecutive [noëtic] derivations) through the noëtic stability fields of families and genera (i.e. every family-groundplan, resp. genus-groundplan, corresponds, in the Implicate Order, to a definite noëtic stability field, determining where in noëtic space the noëtic description of that family- or genus-groundplan is noëtically stable). That is to say, the noëtic trajectory runs from one stability field to another and displaying thereby a branched pattern.
In Part LXIXb, the present Part (LXXa), and the parts to come, it is, however, assumed -- in the sense of merely exploring an alternative possibility -- that the organic world has evolved more or less completely polyphyletically, and polyphyletically already on the species-level (and thus not developing in a branched fashion), and that noëtic counterparts of ecological factors (existential conditions) are active already in the Implicate Order, as described earlier.
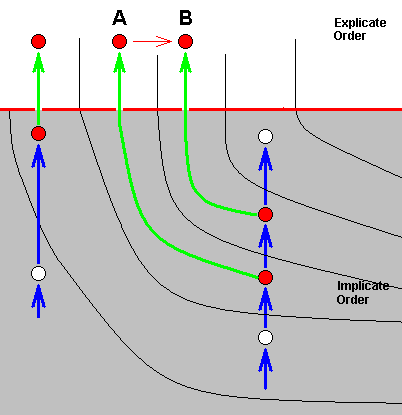
In this polyphyletic picture of things it is, of course, not assumed that already in the Implicate Order every species has appeared ready-made and completely independent from one another. Species there evolve one from the other, forming lines of noëtic descent, each made up of a series of noëtic descriptions of strategies, that may or may not be projected into the Explicate Order and when they do, then appear there as true species. And such a line is the noëtic history of the strategy that did project. That is to say, the noëtic history of such a strategy is the complete series of strategies that derivationally precedes it. And this noëtic history of a given strategy-that-is-projected, that is, of a given existing organismic species, is in fact a bundle of morphoklines in which each constituent morphokline represents the linear series of consecutive transformations of a given organ-system (or part of it) having taken place in this noëtic history. So strategies do, in the Implicate Order, (noëtically) evolve from one another. And there are many cases of two consecutive strategies having both been projected into the Explicate Order, where we see them as two closely related organismic species, A and B, one of which can be derived -- as we have them in the Explicate Order -- from the other (indicated by a red arrow) :
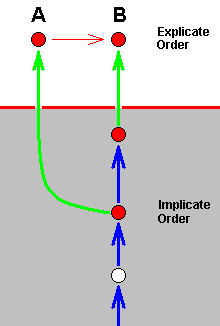
The species A and B belong to the same morphokline, that is, in fact, to the same bundle of morphoklines. And in addition we can say that this situation here displays the classic example of the splitting up of a given species : A transforms into B, in which process A is not exhausted. This results in the presence (in the Explicate Order) of not only B, but also of A. Translated into conventional theory one says that one of the several populations of the original species A has ended up for some reason in different ecological or geographical conditions preventing further contact of this separated pupulation with the rest of A's populations, finally leading to the (physiological and morphological) impossibility of cross-breeding as a result of independent genetic drift on both sides of the (ecological or geographical) barrier. So a new species B has evolved from A, or, equivalently, the original species A has split up into (1) the original species A, and (2) the new species B.
In noëtic terms we can picture this once again but now in a bit extended context and with the ecological zones (in the Implicate and Explicate Orders) explicitly given, and again emphasizing the fact that A and B lie in the same noëtically ancestral line :
And of course it is also possible that B is itself in turn transformed -- in the Implicate Order -- into yet another strategy which is not projected :
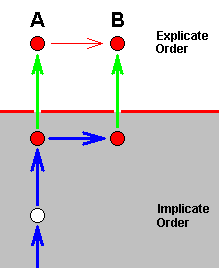
In monophyletic theory, on the other hand, this is pictured as follows :
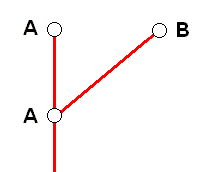
That is, it is pictured as a genuine species-cleavage event, while, according to our noëtic theory it is in fact not.
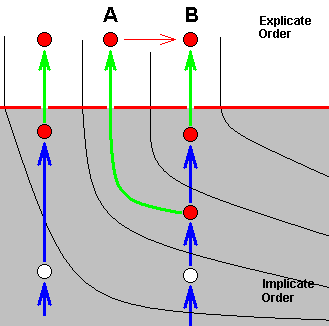
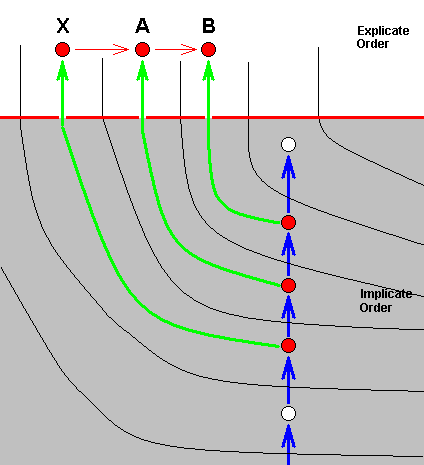
Another type of alleged splitting-up of a species consists in the fact that both daughter-species differ from the original species. The next diagram shows how this looks like in terms of our theory :
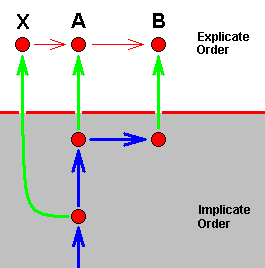
Here strategy X, in the Implicate Order, has been projected into the Explicate Order (without exhausting X in the Implicate Order), but at the same time has, in the Implicate Order, transformed into strategy A (without exhausting X ), which is also projected, while at the same time A has transformed into strategy B (without exhausting A) which is then also projected. So in the Explicate Order we see three species, X, A, and B, where X turns out to be the most primitive of the three, and where B is in a derived state with respect to A (and, by implication, also to X ). Or, in other words, we have an ancestral species (X) which has given rise to two new species, A and B, both different from X. And still we have one single morphokline, ..., X, A, B, in which we see the noëtic history of B (where at least two of its noëtic precursors have also been projected into the Explicate Order), as well as the noëtic history of A (where at least one of its precursors have also been projected).
In conventional theory it is then said that the original species X had two of its populations isolated (from one another and from the rest of X ) developing into respectively the new species A and B (as a result of different genetic drift and subsequent loss of the ability to cross-breed).
In noëtic terms we can picture this once again, with more details, that is, with the ecological zones (in the Implicate and Explicate Orders) explicitly given, and emphasizing the fact that X, A, and B, all lie in a single noëtically ancestral line :
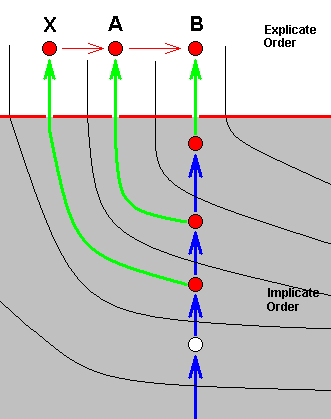
And also here it is possible that B is itself in turn transformed -- in the Implicate Order -- into yet another strategy which is not projected :
In the monophyletic theory, on the other hand, this is pictured as follows :
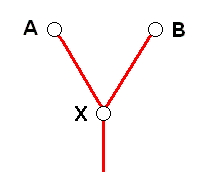
This cladistic diagram depicts the phylogenetic relations existing -- according to monophyletic theory -- between the species X, A, and B, and this independent of the fact whether these species still exist (in the biological-material world) or not.
In phylogenetic systematics (monophyletic theory) it is assumed that in the past large groups (i.e. each time two of such) of organisms -- so called sister-groups -- are each a descendant of one of the (two) species that had resulted from a species-splitting event. And it is the continuous splitting-up of species that results in the ancestral tree of organisms (according to monophyletic theory). This is a very large tree which contains, in addition to fossils, many recent species despite the fact that many other species have become extinct.
As a result, when the spliting-up of the first ancestral species -- the so-called stem-species -- has happened a long time ago, often the two daughter-species have themselves (as a result of subsequent repeated species-cleavages) given rise to two large groups of organisms (such as reptiles and birds) that differ considerably from each other ecologically as well as morphologically.
We, however, cannot accept such an idea, that is, we take it to be very improbable that one single species -- the stem-species -- would be able to evolutionarily give rise to such divergent groups of organisms. Indeed, monophyletic theory assumes, for instance, that all the diverse groups in the Order Diptera have ultimately originated from a single mecopterous (scorpionfly-like) species (while the other mecopterous species have not done so). Such a state of affairs is argued on the basis of the common possession by all the diptera (to stick with this example) of certain derived characters such as the transformation of the mecopterous hind-wings into halteres (specialized organs having to do with the stabilization of flight) leaving the insect with only one pair of wings. It is, in monophyletic theory, assumed that this transformation has happened only once, in a single species, as a result of a single random genetic mutation (or a coherent series of such mutations) and subsequent natural selection, and then genetically passed on to all the descendants of that stem-species. So the mentioned transformation of the hind-wings is taken to be a unique event in the history of insects.
It is, however, difficult to prove this uniqueness without having recourse to the phenomenon of random genetic mutations. And this is why monophyletic theory heavily relies on neodarwinian evolutionary theory. We, on the other hand, take it to be more probable that the mentioned transformation was a regular process, a process that is, that has taken place independently in a relatively large number of mecopterous insects as a consequence of their special intrinsic qualities naturally leading to that transformation in the course of their noëtic history. Arguments like this lead us to a denial of species-cleavage being a crucial factor in organic evolution. And that's why we investigate the possibility and consequences of a thorough and complete polyphyly in evolution, all the way down to the species level.
Species-cleavage, even as it may take place in the way depicted in the above diagrams of morphoklines, is not the rule however. It is just a particular configuration of noëtic derivations and projections of which the result, as seen exclusively in the Explicate Order, may be interpreted as cases of species-cleavage. And if we assume -- just as an alternative theory of which the implications are being explored -- complete polyphyly in the evolution of organisms, meaning the absence of any monophyletic groups, except those of the types just described, and thus meaning the presence of all-out polyphyly not only of genera, families, etc. [i.e. not only the independent origin and development of genera and of families, etc.] but also [the independent origin] of species, that is, of the lowest unit of strategy that can exist in the Implicate Order, if we assume all this, then species-cleaving (apart from the cases mentioned) cannot take place. Each species, existing or having existed in the Explicate Order, has its own morphokline [in fact a bundle of morphoklines], i.e. has its own noëtic history independently extending into the depths of the Implicate Order, and not merging there with other such morphoklines. So in the Implicate Order we have the many 'masses' of parallel morphoklines, i.e. parallel bundles of morphoklines (each such a bundle consisting of, say, a venational, chaetotaxic, neurophysiologic, etc. etc. morphokline, together -- as a bundle -- constituting the series of consecutive noëtic transformations of one strategy into another), -- masses of parallel morphoklines that is, that do not show branches, but are just independent parallel lines [All this should be placed in the context of exploring the possibilities, implications and merits of a more or less alternative theory -- complete polyphyly of organic evolution --, alternative that is, with respect to our earlier theory of a b r a n c h e d noëtic trajectory (visiting stability fields in the Implicate Order), and subsequent projections, and thus contrary to monophyletic theory (which was an assumption of Phylogenetic Systematics)]. The results of phylogenetic systematics are presented in the form of a cladogram depicting the phylogenetic relations between all the species of some given group of organisms. The cladogram is supposed to display the phylogenetic history of the group, and this history is in fact held to be a succession of species-cleavages, as we ourself have done with the Bibionomorpha in Part XXVIII and Part XXVIII-A. The arrangement of the species in such a cladogram or phylogenetic tree [species that is, even when only families are mentioned in it] is based on the morphological, etc. characters of the representatives of these species. For example in Bibionomorpha it was based on wing-venation. So this arrangement itself must, in one way or another, truly reflect the transformational history of the group [where we place this 'history' in the Implicate Order]. What we now, i.e. in our assumption of complete polyphyly, are going to do is to preserve the arrangement of species (by displaying them as derivational lines), but to do away with any form of monophyletic development, and that means to do away with species-cleavage (in every cladogram supposed to have taken place).
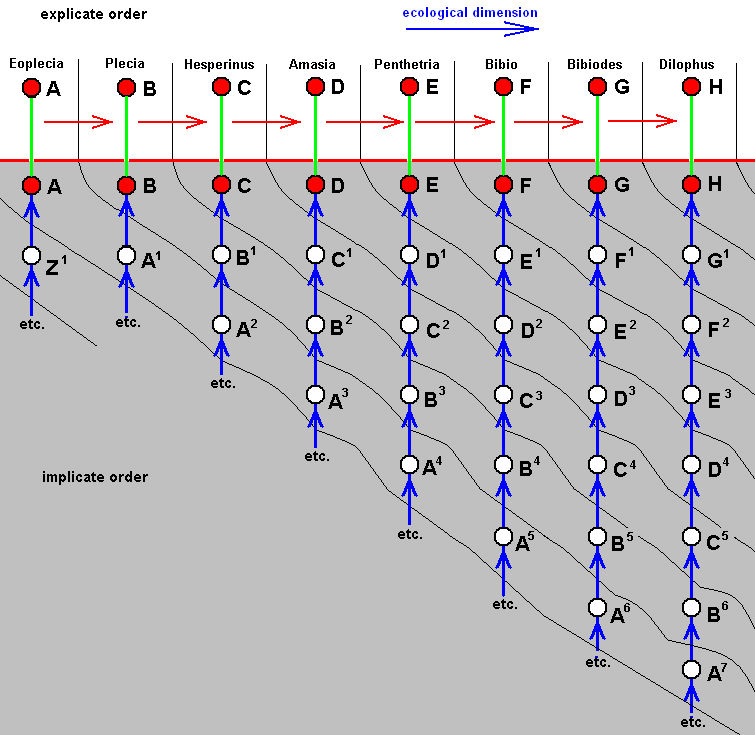
So we're going to express the cladogram (such as that obtained in Bibionomorpha), and that is, to express the arrangement of species in it, in terms of complete polyphyly (by deriving from each derivational line, obtained in that arrangement, a series of morpholkines [in fact a series of bundles of them] see, for instance HERE for the derivational line (B, C, D, E) and the four corresponding (venational) morphoklines. This means that every species representing a bifurcation-point in the cladogram, which species is -- in the majority of cases -- in fact merely a hypothetical species, must be replaced by a number of more or less closely related species.
Already in each morphokline we do not let a given existing species originate from the previous one in the corresponding derivational line, but let it originate from its own immediate noëtic precursor which itself is not, but only looks like, the mentioned previous species in the derivational line. This 'looked like', this similarity, here is supposed to be the result of their being associated with the (noëtic counterpart of the) same ecological zone, see, for instance, again the derivational line of the species B, C, D, E , and the series of venational morphoklines derived from it HERE , where, for instance species D is not supposed to have originated from its previous species C in the derivational line, but from species C' which (venationally) looks like species C, and where C' and C are (noëtically) associated with the same ecological zone.
On the other hand, the similarity we spoke about earlier, i.e. the similarity of the several species together representing a given bifurcation-point in a cladogram in phylogenetic systematics (based on monophyletic theory), is supposed to be of a more or less different kind (of similarity), i.e. not a similarity as, for instance there is between the species C and C' just mentioned, a similarity that is in some way associated with a particular ecological zone common to both species, but a similarity in morphological characters, not directly associated with some particular ecological zone. Indeed the various species together representing such a bifurcation point are supposed to differ morphologically, such that their respective descendants will go their own way, leading in the end to the origin of different groups in the sense of different morphological types.
Let us now -- to offer a general example -- show how we should translate a cladogram of a given, but still imaginary, allegedly monophyletic group of species of organisms, a cladogram supposed to be empirically obtained by the methods of phylogenetic systematics, into the terms of derivational lines and morphoklines, that is, in terms of complete polyphyly :
To begin with, we here give the (imaginary) cladogram that phylogenetically arranges the species A, B, C, D, E, F, and G :
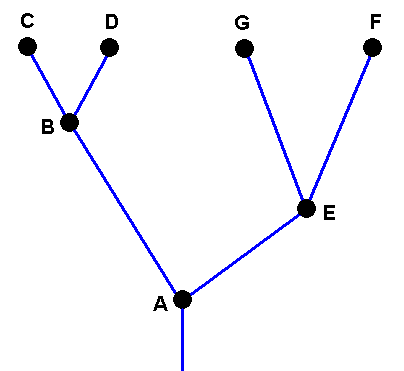
Cladogram of the group (A, B, C, D, E, G, F) depicting the phylogenetic relations between its species.
Species A is supposed to have bifurcated into the species B and E, the species B is supposed to have bifurcated into the species C and D, while the species E is supposed to have befurcated into the species G and F.
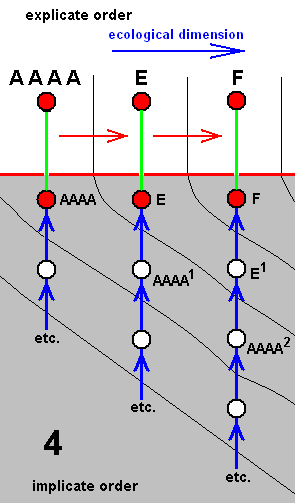
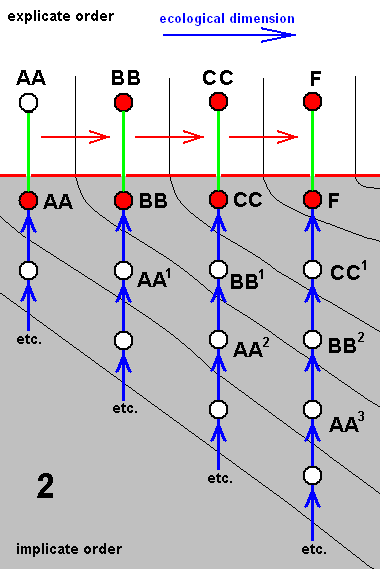
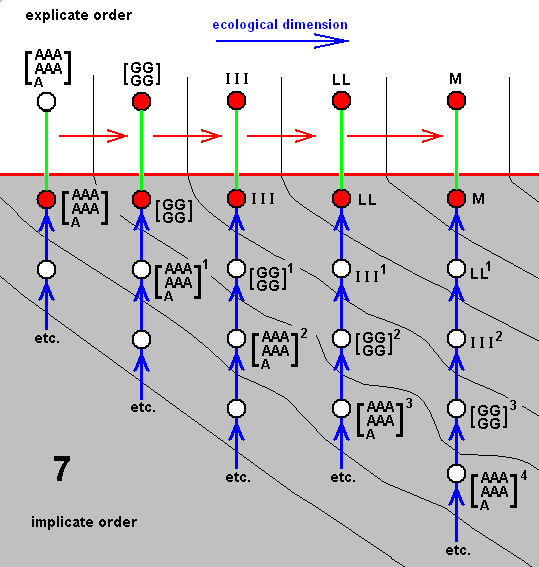
In our theory we suppose that all species, and thus also the species C, D, G, and F, each have their own independent history, i.e. a derivational history in the Implicate Order, which means that the species B did not bifurcate, simply because the bifurcation-point in the cladogram is in fact represented not by the single species B, but by two morphologically similar species B and BB. The same aplies to the species E. Also there the bifurcation-point is represented by two species E and EE. And with respect the species A we say that there the bifurcation-point is represented by four similar species, viz., A, AA, AAA, and AAAA :
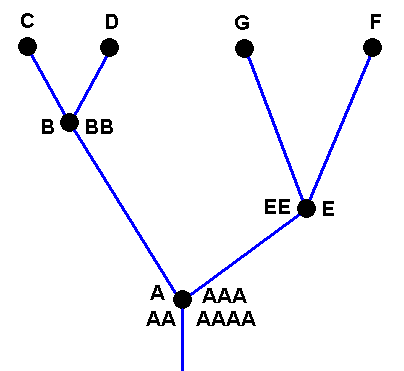
Cladogram of the group (A, B, C, D, E, G, F) now having its three bifurcating species A, B, and E replaced by three clusters each containing independent but similar species : (A, AA, AAA, AAAA), (B, BB), and (E, EE).
Accordingly, the cladogram will now be changed into the following diagram of derivational lines :
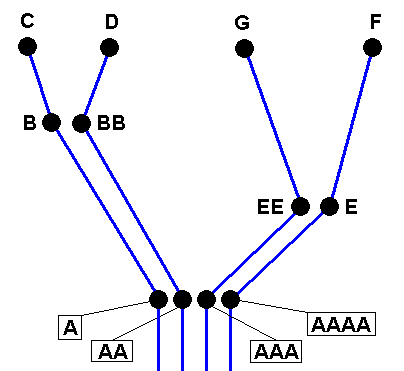
Four derivational lines extracted from the cladogram of the group (A, B, C, D, E, G, F) :
A==>B==>C,
AA==>BB==>D,
AAA==>EE==>G and
AAAA==>E==>F.
The possible derivations expressed by these lines essentially were the basis on which the original cladogram was determined. But there they were, of course, A ==> B ==> C, A ==> B ==> D, A ==> E ==> F, and A ==> E ==> G.
In the present diagram depicting derivational lines, as well as in the above diagrams cladistically depicting monophyletic groups, all mentioned species, i.e. all species figuring in these diagrams, whether as actually existing or fossil, are supposed to exist or having existed in - what we call -- the Explicate Order.
From each one of these derivational lines we will now construct the corresponding group of morphoklines, each morphokline representing the noëtic history of the relevant species figuring in the derivational line. And still, the order of the species in a given derivational line suggest (and no more than that) in what way the transformations might have taken place in the noëtic history of the species of the derivational line. Transformations of what? Well, first of all the transformation of that particular morphological structure of which the morphoklines are about, that is, venational transformations when the morphoklines are venational morphoklines, ans chaetotaxic transformations when the morphoklines are morphoklines of chaetotaxy, etc. But although in all cases we deal with morphoklines that are about the transformation of a single character (such as wing-venation), we have in fact in mind the whole bundle of morphoklines, together dealing with the transformation of all characters of the species-in-question from the derivational line.
Let us now then set up the corresponding groups of morphoklines, to be constructed from the just determined four derivational lines :
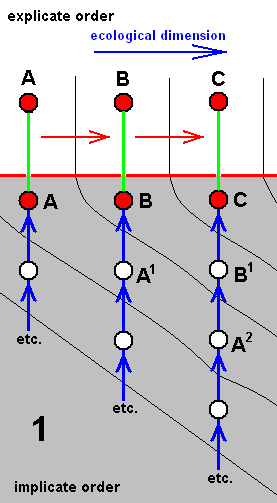
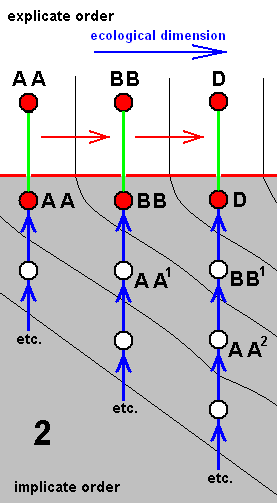
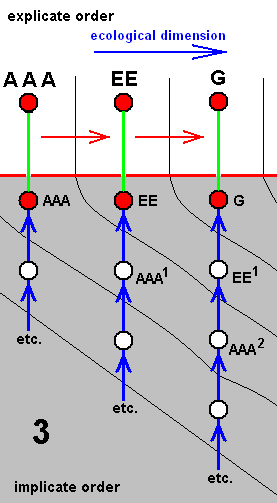
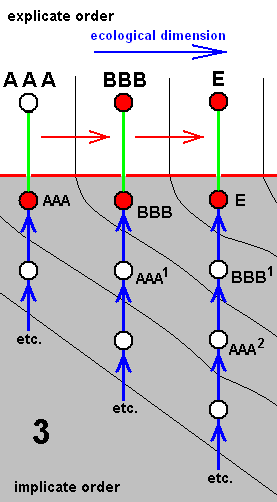
Each species of a given derivational line is the last member of a morphokline. And because all species figuring in a derivational line are supposed to exist or having existed in the Explicate Order (in addition to their existence as immaterial patterns -- noëtic descriptions of strategies -- in the Implicate Order), such a species is placed in the diagram of morphoklines first of all into its explicate-order-part (as a red disc). As such it is then seen as the projection, from the Implicate Order, of its noëtic counterpart. So this counterpart -- also indicated by a red disc -- is placed into the implicate-order-part of that same diagram of morphoklines. Its projection into the Explicate Order is indicated by a green line [In fact symbolizing the incessant projection, injection, re-projection, etc. of such a strategy]. And the mentioned noëtic counterpart of the species is then the top member of the morphokline, which extends 'downwards' into the 'depths' of the Implicate Order (noëtic space). The morphokline consists of 'hypothetical species' -- indicated by uncolored discs -- which in fact are, of course, not exactly species, but hypothetical noëtic descriptions of strategies. So here the "hypothetical" refers to existence in the Implicate Order only. They are supposed to be strategies that have not been projected, and so do not figure in the Explicate Order.
For instance in the second (2) of the above four diagrams of morphoklines BB1 is a hypothetical strategy in the Implicate Order. It is, when indeed existing in that Order, not projected into the Explicate Order. However it, as a noëtic description, is similar to BB, which is the noëtic counterpart of the explicate-order-species BB, that is, BB is a species existing or having existed in the Explicate Order. And the (hypothetical) strategy BB1 is supposed to noëtically having given rise to strategy D (which is non-hypothetical with respect to existence in the Implicate Order), which has then projected into the Explicate Order as the species D (which is not hypothetical, neither with respect to existence in the Implicate nor with respect to existence in the Explicate Order). The species D (second (2) diagram) is the terminal member of the derivational line ( AA==>BB==>D ). The transformation of the character (such as wing-venation) with which the morphokline deals, that is, the transformation of the corresponding noëtic description in the morphokline of species D, is suggested by the sequence of the three species of the derivational line AA==>BB==>D. Correspondingly, the morphokline of species D (i.e. its supposed noëtic history) is : ..., AA2, BB1, D.
We should thereby read AA1 as (AA)1, or, equivalently, as (AA)1, or as AA' for that matter. Likewise we should read AAA2 as (AAA)2, or, equivalently, as (AAA)2, or AAA'', and read EE1 as (EE)1, or, equivalently, as (EE)1, or EE', etc.
We have given much attention to the splitting-up of species (generally into two daughter species) because it plays an important role in the monophyletic theory of evolution. It is presupposed by Phylogenetic Systematics.
But all this has not yet been enough to transform monophyletic theory into polyphyletic theory.
In monophyletic theory every taxon above the species level is considered to be a monophyletic group of species. Especially a family is supposed to be such a monophyletic group, meaning that all members, and only these members, have been descended from a single stem-species. And we, following Hennig, 1954, have treated families as units in the phylogenetic system of the Bibionomorpha (meaning that the result is a highly compressed phylogenetic system, because its real units are species, not families). And this has been done on the basis of the wing-venation (of course with the assumption that characters other than wing-venation will follow the same derivational pattern [and not causing specialization-crossings!] ).
But if we assume -- as an alternative theory -- that every organismic species, and thus also every species in the Bibionomorpha, has its own independent noëtic history, its own morphokline, then every taxon above the species level cannot be phylogenetically defined anymore, i.e. cannot be interpreted as being a monophyletic group of species, but as being a morpho-biological type. And the boundaries of such a type are not expected to fully coincide with those set by phylogenetic systematics, thay is, the criteria to distinguish such types are partly different from those that distinguish phylogenetic groups (with respect to the latter the criterium is monophyly, demonstrated by the common possession of derived characters (synapomorphy), derived that is, with respect to species closest to those of that phylogenetic group).
A family, then, is, in our theory, not a monophyletic group, but a type (in the sense of typological systematics, as we see it, for example, in the works of Rohdendorf). And because in the practise of phylogenetic systematics monophyly it is often hard to establish anyway, phylogenetic systematics often merely assumes that a family -- as it was already earlier established as such, i.e. as a taxon, often on the basis of typological criteria -- is also a monophyletic group as long as it is not demonstrated not to be so.
So although the members of a family form, in our theory, a polyphyletic assemblage of species, each with its own independent morphokline (noëtic history), they nevertheless form a unit, not phylogenetically, but typologically. And although a given type cannot so neatly be defined as can a monophyletic group, it can be approximately circumscribed in terms of morphological and ecological properties. We assume that all members of a family possess some common kernel in their qualitative content, that is, some common elements in their essence or whatness, elements different from those in other families. And these common elements are not inherited from some single 'stem-species', but have independently evolved in the noëtic history of each species/strategy of that family, that is, the common possession of these elements is not the result of common ancestry.
Let us work out a more or less concrete example.
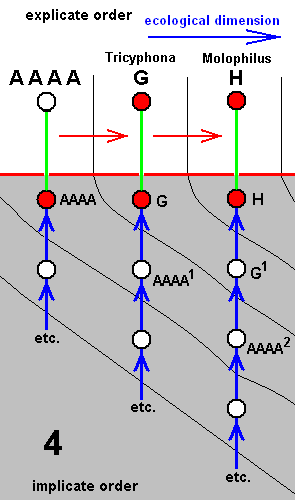
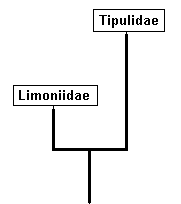
Phylogenetically thinking we may suppose that the dipterous families Tipulidae and Limoniidae are sister-groups, and that both, of course are monophyletic. The next diagram depicts a very simplified version of the phylogenetic system of ( Tipulidae + Limoniidae).
For Tipulidae we enter a species of the genus Pales (coded as B) while for the Limoniidae we enter species of the genera Tricyphona and Molophilus, coded as G and H respectively. The species A is hypothetical, it is supposed to be the stem-species of Tipulidae + Limoniidae, but will, despite its hypothetical status, be entered in our derivational lines and morphoklines.
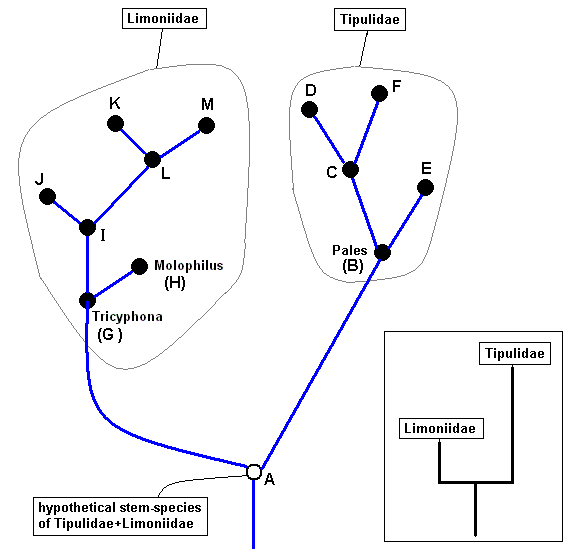
Simplified cladogram of Limoniidae + Tipulidae.
Tipulidae and Limoniidae are supposed to be sister-groups, i.e. both having evolved from a single ancestral species (A), the stem-species.
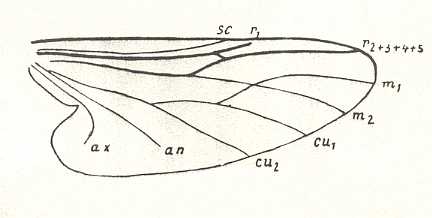
The insert summarizes the phylogenetic origination of the family Tipulidae from the family Limoniidae, that is, the family Tipulidae originates from some primitive member, or direct ancestor (A), of the family Limoniidae, based on the fact that the " capture of R4 by R2+3 " (i.e. the origination of the fork of R2+3 from R4 instead of from the main trunk of RS (see HERE ) ) is present in all Tipulidae, but not in Tricyphona and Molophilus in Limoniidae, meaning that it was not yet present in the stem-species of Limoniidae + Tipulidae (and from this stem-species (also) the Tipulidae originated).
In the same way as we did ABOVE , we can -- as a result of denying species-cleavages -- distinguish seven derivational lines :
And from these derivational lines we can set up seven diagrams of morphoklines, i.e. a morphokline for each species of the group.
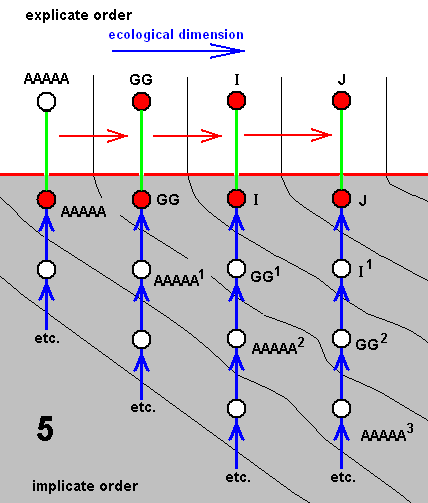
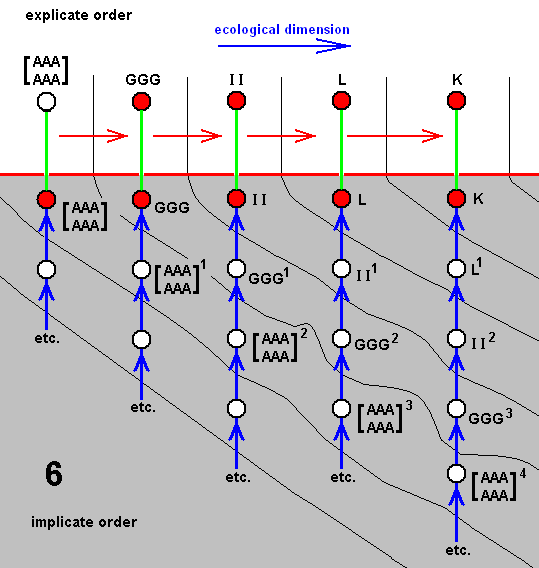
All these seven series of morphoklines (and in reality much more of course) are contained in the compressed cladogram
as soon as this cladogram is translated into the picture of polyphyletic theory.
Returning to our general noëtic theory, we now assume that the aforementioned fields of noëtic stability with respect to given noëtic patterns (such as descriptions of strategies), that is, fields in noëtic space in which particular noëtic patterns are stable while others are not, do -- especially in all the cases of organic strategies -- coincide with the noëtically-represented actual and potential ecological existential conditions described earlier. Indeed, from these latter the fact of noëtic stability directly follows. And these fields of existential conditions in turn more or less coincide with the (noëtic description of the) groundplans of taxonomic categories such as species, genera, families, superfamilies and orders within the Class of Insecta (and generally with the groundplans of taxonomic categories of organisms as such), because these taxa embody strategies-to-exist and are associated with special ecological conditions. And although there is probably no complete correspondence between taxon, strategy, and ecological condition (niche or zone), it generally is as such present, as is shown by writers such as Rohdendorf.
Venational Derivational Lines and Morphoklines in Bibionomorpha (Diptera-Nematocera).
On the basis of derivational relations in the wing-venation of representatives of the genera and families of the Bibionomorpha, as we had found these relations in Part XXVIII and Part XXVIII-A , we will now establish a number of derivational lines, from each one of which we directly derive the diagram of morphoklines (i.e. the corresponding set of venational morphoklines). Into these we will enter fossil as well as recent species.
But before doing so we first present here the results obtained in Part XXVIII and XXVII-A as to the phylogeny (family level) of the Bibionomorpha according to the methods of phylogenetic systematics (= monophyletic theory of evolution). In the Bibionomorpha we distinguish the Rhyphiformia, the Bibioniformia, the mesozoic Parafungivoriformia, and the Fungivoriformia. The phylogenetic series begins with the Rhyphiformia.
The reader may click on the family names to check the decisive characters :
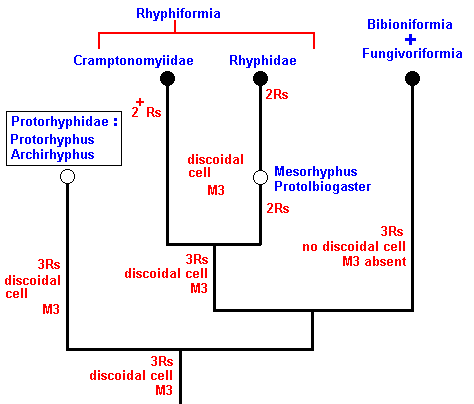
Figure 7 : Phylogeny of the Rhyphiformia and Protorhyphidae. Indicated are the venational transformations on which this phylogenetic system is based. The evolution leading from the Protorhyphidae to the Rhyphiformia consists in the reduction of the number of branches of the Radial Sector (from 3 to 2 [or 'almost' 2] ) while the discoidal cell remains. The evolution that leads to the Bibioniformia + Fungivoriformia consists in the initial preservation of a 3-branched Radial Sector, the disappearance of the discoidal cell, and the vanishing of the vein M3.
Here, "3Rs" means : Radial Sector is 3-branched, while "2Rs" means : Radial Sector is 2-branched. And "2+Rs" means : Radial Sector with three branches, of which one is very short, that is, almost vanishing. "M3" means : The vein M3 is present.
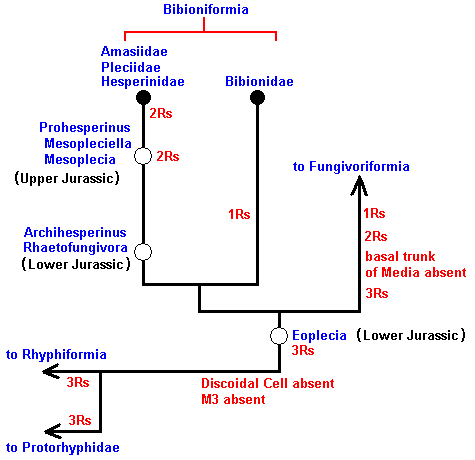
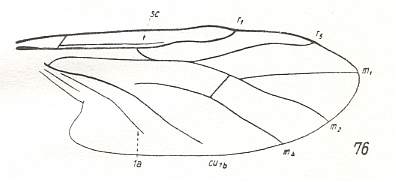
Figure 16 : Phylogenetic tree of the Bibioniformia, and the places in this tree where the fossils should belong.
To see these fossils, click on their names : Prohesperinus Mesopleciella Mesoplecia Archihesperinus Rhaetofungivora Eoplecia
Having showed the structure, i.e. the cladistics, of the first divisions (Rhyphiformia and Bibioniformia) of the Bibionomorpha according to the monophyletic theory of evolution (and based on wing-venation), we will now investigate the venational derivational lines and their corresponding diagrams of morphoklines, showing the polyphyletic evolution of these first divisions [and in the next document we will occupy ourselves with the largest and central group of the Bibionomorpha, the Fungivoriformia].
Derivational LINE 1 ( Protorhyphidae + Rhyphiformia, for phylogeny see cladogram above ) (see for the original configuration of the Medial-Cubital region in Bibionomorpha HERE ):

|
Protorhyphus stigmaticus, Protorhyphidae. Upper Lias (= upper lower Jurassic) of Germany. Radial Sector (RS) 3-branched. M1, M2, M3, and M4 present. Discoidal cell present. CuA and 1A separate. Cross-veins ta [= r-m] and tb2 [= m-cu] present. |

|
Cramptonomyia spenceri, Cramptonomyiidae (or just Rhyphidae). RS 3-branched, but one of them only visible as a crossvein-like veinlet. M1, M2, M3, and M4 present. CuA and 1A parallel. |
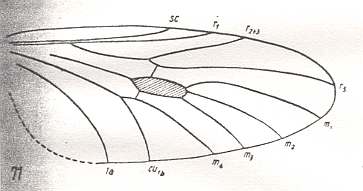
|
Mesorhyphus nanus, Rhyphidae. Upper Lias (= upper lower Jurassic) of Germany. RS 2-branched. M1, M2, M3, and M4 present. CuA and 1A parallel. |
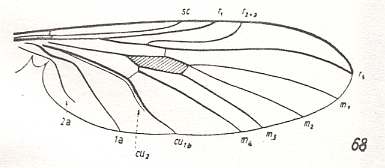
|
Phryne fuscipennis, Rhyphidae. RS 2-branched. M1, M2, M3, and M4 present. CuA and 1A parallel. Venation virtually the same as in Mesorhyphus. The here depicted wing-venation of the recent Phryne being virtually identical to that of the mesozoic Mesorhyphus is normally seen as evidence of phylogenetic descent (see cladogram above ). But this need not be so : First, of Mesorhyphus only its wing-venation is known to us. Secondly, an identical morphological character (here the wing-venation) may be part of a similar adaptation as a result of similar ecological conditions, an adaptation independently obtained by different forms, not phylogenetically related to each other. |
Although the above four venations are pretty well derivable, one from the other, it is nevertheless safest to assign to each one of them its own separate morphokline, because (1) two of the four species are only known as wing-impressions (in a sediment) implying that, even if they were assigned to the same genus we could not know that for certain. It is, therefore safest to assume them to belong to different genera, to which indeed they have been assigned, different not only from each other but also from the other genera in this derivational line, and (2) the other two wings of this line (wings of recent flies) have been assigned to different genera as well, viz., Cramptonomyia and Phryne.
The corresponding diagram of morphoklines :
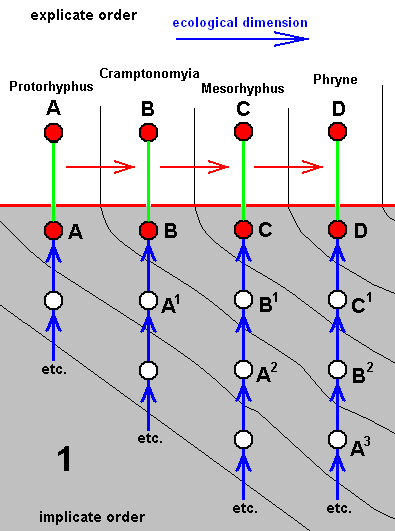
Short theoretic intermezzo
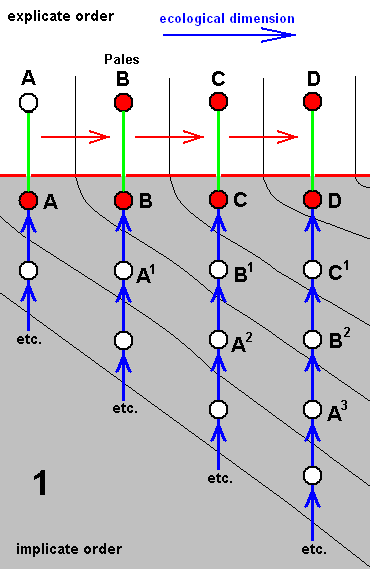
The above diagram of morphoklines depicts the separate development -- in the Implicate Order -- of four species of rhyphoids. In what way precisely this development has taken place [i.e. what its initial stage 'was' and what the intermediate stages 'were'] is supposed to be more or less reflected in the derivational relations that turn out to exist between the existing species A, B, C, D, together forming a derivational line. Generally the corresponding diagram of morphoklines, concerning four species looks like this :
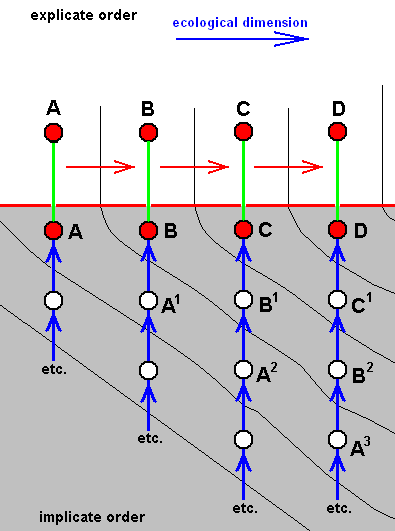
The existing species A, B, C, and D, differ from each other by the one being in a derived condition with respect to the previous one, and thus forming the derivational line A==>B==>C==>D. But they are not strictly and in all respects so derivable from each other. It is only a reflection of the true sequence of strict derivation A3==>B2==>C1==>D (while A==>B==>C is a reflection of the sequence of strict derivation A2==>B1==>C, and A==>B of A1==>B ).
And in fact the morphoklines are here supposed to be first of all venational morphoklines, representing the derivationability of one [noëtic description of] venation from another in these mentioned sequences of strict derivation. But in a diagram like this we subsequently assume that A1, A2, A3, B1, B2, C1, etc. represent complete species (as also do A, B, C, and D in the Implicate Order) [i.e. they are, descriptions of true and complete strategies (s.l.) ] implying that we here have to do with sequences of strict derivation not only of wing-venation but of all characters of these species [indeed, when the species as a whole are derivable from one another then all characters of them are so derivable].
In such a diagram we, for instance say that existing species C, although more or less derivable from existing species B, is strictly derivable (and actually derived) from B1 which is similar to B (this similarity is expressed by their being associated with the same ecological zone). And if we look to a pair of adjacent morphoklines, say those of C and D, we see that while developing [i.e. while being derived] they cross ecological zones [i.e. their noëtic counterparts] one after the other, whereby at each derivational level [each such level representing a derivational step and being horizontal in the diagram] the noëtic precursors of D are 'ecologically' one step ahead of C, that is, having 'arrived' in the next ecological zone. So this is how things are in the case of a derivational line, i.e. in the case of a series of species that are more or less derivable from one another (implying, by the way, that they strictly are neither identical nor similar to each other).
On the other hand, there are also cases of a number of existing species being in such a degree similar to each other that between them there does not exist a one-way derivability. In the context of our theory they are species that have developed in parallel, in exact parallel that is. The nature and degree of similarity between them is the same as that between C and C1, or, say, between A1 and A2, in the above diagram, but without one species/strategy being ahead of another. See next diagram, concerning the species O, P, Q, and R.
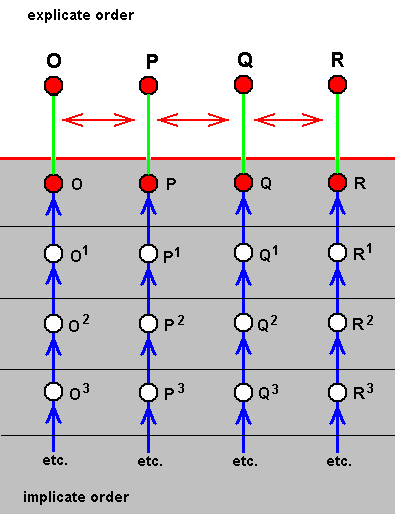
Figure above : Diagram of parallel morphoklines.
These four morphoklines go together -- in step -- from one ecological zone to another, i.e. from the noëtic description of one ecological zone to that of another. So they are strictly parallel derivational sequences in the Implicate Order, beginning from similar (but not identical) stages and leading, not to identical, but to similar end-products.
The part of the morphokline of species O actually depicted here is O3==>O2==>O1==>O.
The part of the morphokline of species P actually depicted here is P3==>P2==>P1==>P.
The part of the morphokline of species Q actually depicted here is Q3==>Q2==>Q1==>Q.
The part of the morphokline of species R actually depicted here is R3==>R2==>R1==>R.
The series of existing species O, P, Q, R is not a derivational line, but a series of products of parallel noëtic development. Not any morphokline in this diagram of parallel morphoklines is reflected by the series of existing species O, P, Q, R.
A strict polyphyly, i.e. polyphyletic development all the way down to the species-level -- the species being the ultimate elements of the polyphyletic scheme -- demands a taxonomic classification (systematics) fully compatible with it. Indeed such a taxonomic system should not reflect common descent, because there is no such a descent. It reflects the qualitative relationships existing between these species. And these species are the ones existing in the Explicate Order. In fact the proper taxonomic system reflects the relationships existing between the qualitative intrinsic contents of them -- the species' qualitative kinship -- in exactly the sense of the pre-darwinian rational morphologists, the discoverers of homology. Indeed, to them the morpological relationships existing between organisms were ideal relationships, not the result of material genealogical descent, but of idealistic derivations. And the species of the taxonomic system [this system being] compatible with polyphyletic development are the true material species of the Explicate Order, that is, strategies actually existing or having existed materially in the Explicate Order. There they appear as individuals -- individual organisms in time and space -- of the different organismic species as soon as the appropriate ecological conditions are actually satisfied in the Explicate Order. So our taxonomic system of organisms should, first of all, reflect the similarities in qualitative intrinsic content of the species of the system, this content being reflected in the species-constant biochemistry, morphology, physiology, and behavior. Further it should reflect the order of actual appearance in the Explicate Order, as is documented by fossils. So defined, our taxonomic system essentially will be very close, in structure and intent, to typological systems such as we see in, for example, Handlirsch, Rohdendorf, Sharov, and many others, but without their monophyletic completions, i.e. without their lines finally converging in common ancestors. And of course our taxonomic system stands, as to its general structure and intent, in contrast to the taxonomic systems set up by phylogeneticists working strictly along hennigian lines.
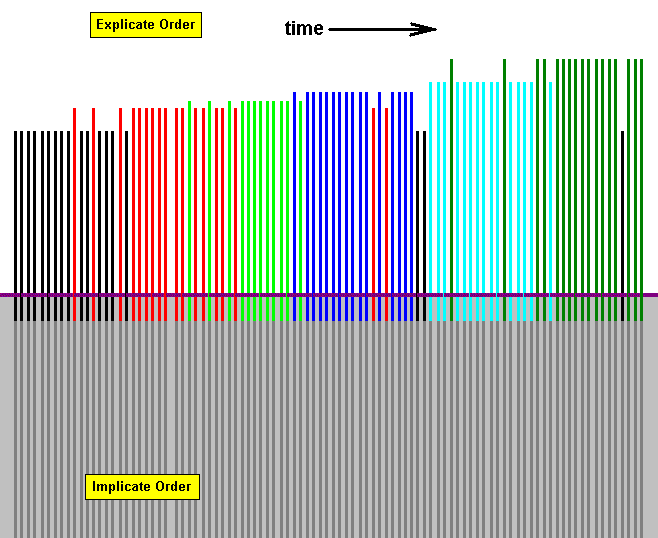
Figure above : Diagram of parallel morphoklines of species of a number of different families (indicated by different colors).
While in the Implicate Order there is no time dimension, in the Explicate Order there is such a dimension as indicated by the arrow in the explicate-order-part of the diagram. Depending on existing proper ecological conditions in the Explicate Order, the strategies, at last noëtically completed in the Implicate Order, will project into the Explicate Order and appear there in the form of individual organisms of various species, each species/strategy symbolized by a vertical line, ascending from the Implicate into the Explicate Order. The nature of the qualitative content of these species is symbolized by the color of the lines.
The present diagram symbolizes the successive appearance in the Explicate Order of, to begin with, species of a particular family indicated by black lines. During the course of time species of another family (red) appear contemporaneously with certain later species of the first family. Then this first family (black lines) becomes extinct while species of the second family (red) continue to appear. Then, long before this latter family (red) gets extinct, another family (green) appears, and keeps on appearing also after the red family has become extinct. Then just before the time of extinction of the green family, a new family appears (dark blue) and its species keep on appearing for a considerable long time, during which, however, a few species, and not more than a few species, of the red family appear. Then, immediately after the blue family has become extinct, a few species of the first family (black) appear, and after them yet a new family (light blue) appears and keeps on appearing for some time during which yet another new family (dark green) begins to appear intermittently. And after the light blue family has disappeared, species of the dark green family continue to appear, and -- unexpectedly -- during the continuous appearance of this dark green family, one single species of the very first family (black) appears, but soon extinguishes.
And it should be clear that any case of the 'reappearance' of some earlier family must not be interpreted as a phenomenon of genealogic descent of the later from the earlier species. Such a reappearance must have to do with the reappearance of certain earlier ecological conditions, the later version of them may contain some new elements (organic species that did not yet exist at the time of those earlier conditions). And when these new elements do not oppose the qualitative content of species of the earlier family, it will indeed reappear and may then be found in the ecological conditions still compatible with it.
Further, in the present diagram, the height of the colored lines may represent the degree of anagenesis (= the degree of organizational complexity and integration of body and behavior).
So all this indeed represents the general structure of the taxonomic system compatible with strict polyphyletic development of organisms. This development, however, does not (as, for instance, Lichtig 1938 [see below], assumes) take place in the Explicate Order, but in the Implicate Order in the form of independent morphoklines.
While we assume that the derivational line of existing species (i.e. existing or having existed in the Explicate Order) more or less reflects the noëtic histories of those species (having 'taken place' in the Implicate Order), it (i.e. the derivational line) does not necessarily reflect the order of (first) appearance of these species in the Explicate Order, because projection depends on the presence of the appropriate existential conditions in the Explicate Order. For example, a more derived strategy (being placed more to the right in the derivational line) may be projected earlier than a less derived strategy in that same derivational line (and to polyphyletic theory this doesn't pose a problem!). However we, for the time being, assume that cases like this are rare exceptions (accounting for certain fossils [of a given group] being more derived than their fellow species in the recent fauna or flora). We might theorize that generally the projection of more derived strategies presupposes the earlier projection of less derived strategies, which conforms to our general paleontologic evidence of more ancient (groups of) species being less derived (more primitive) than less ancient ones. And we also theorize that generally "more derived" coincides with "higher developed", i.e. having taken a step forward in organizational height, that is, in anagenesis. Also this generally conforms with paleontologic evidence.
Whether these supposed relationships between derivational line, morphokline, order of appearance (projection) in(to) the Explicate Order, and organizational height, are actually present, and if so, precisely in what way, is a very interesting question, and indeed attempting to answer it should constitute the very core of evolutionary research. At least we, on this website will so attempt.
( End of theoretic intermezzo)
Derivational LINE 2 ( Bibioniformia, for phylogeny see cladogram above ) :
|
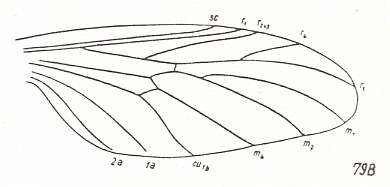
Eoplecia primitiva, Upper Liassic (= upper lower Jurassic) of Germany. Radial Sector (RS) 3-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. CuA and 1A separate. Cross-veins ta [= r-m] and tb2 [= m-cu] present. 2A present. |
|
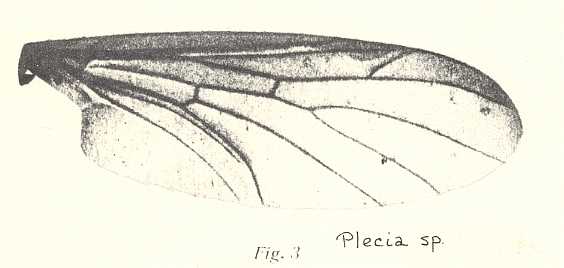
Wing of Plecia sp. Pleciidae (= Penthetriidae). RS 2-branched. First branch relatively short. The vein running just behind CuA is CuP. Vein 1A present, separate from CuA. Vein 2A absent or strongly reduced. The venational correspondence seen to be present between forms like the recent Plecia and the mesozoic&n,,,,,,, althoughbsp; Mesoplecia should not a priori be seen as evidence of genealogic descent (see cladogram above ) , but just as a same adaptational or morphological element having independently appeared in different forms (genera) at different times. |
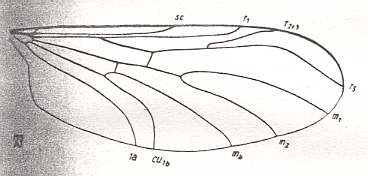
|
Hesperinus imbecillus, Hesperinidae. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. CuA and 1A parallel. Origin of first branch of RS shifted a bit basad. |
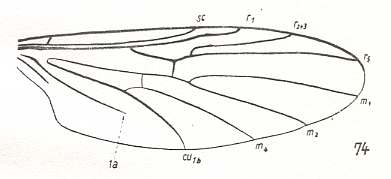
|
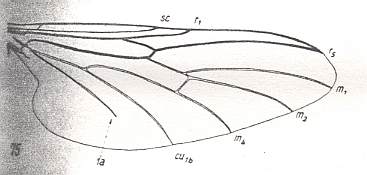
Amasia funebris, Amasiidae. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. 1A shortened, not reaching wing-margin. Origin of first branch of RS shifted basad still more. |
|
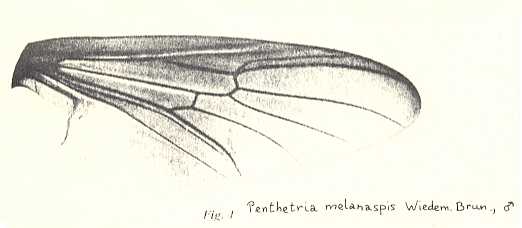
Wing of Penthetria melanaspis, Pleciidae (= Penthetriidae). RS 2-branched. First branch long. In this photograph the vein CuP (posterior Cubitus) is clearly visible. It runs closely along CuA (anterior Cubitus). The cross-vein ta [= r-m] lands on M1 instead of on the common stalk of M1 and M2. Origin of first branch of RS still more shifted basad. It is now precisely at the level of the cross-vein ta. |
|
Bibio marci, Bibionidae. Radial Sector (RS) unbranched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. Main trunk of Media still strongly present, at least until the cross-vein tb1 (which is placed rather distally). CuA and 1A separate. 1A shortened. Cross-veins ta [= r-m] and tb2 [= m-cu] present, but the latter is strongly drawn out. |
|
Bibiodes spec. Bibionidae. Radial Sector (RS) unbranched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. Also here the main trunk of Media is still strongly present. It has coalesced along a short distance with the Radial Sector. As a result the cross-vein ta [= r-m] is suppressed. CuA and 1A separate. both shortened, not reaching wing-margin. |
|
Dilophus albipennis spec. Bibionidae. Radial Sector (RS) unbranched. M1, M2, and M4 present. M3 absent. (Cu1 in the figure is M4). Discoidal cell absent. While in Bibio the veinlet ta is still clearly manifesting itself as a genuine cross-vein, in Dilophus it lines up with the Radial Sector, rendering this region of the venation look very specialized. The same configuration is shown in the region of the cross-vein tb1. Cross-vein tb2 [= m-cu] drawn out. CuA (Cu2 in the Figure) separate from 1A (an in the Figure). The latter not reaching wing-margin. The vein 2A (ax in the Figure) apparently present, not reaching wing-margin. The primitive condition of ta does in fact not allow Dilophus to be derived from Bibiodes. This adds to the fact that existing species generally are not strictly derivable from one another. The venation of Dilophus may be derivable from that of a more or less close relative of Bibiodes. |
The corresponding diagram of morphoklines :
A number of fossil Bibioniformia centering around the recent genus Hesperinus form a series of parallel lines of noëtic development (see also their place in the phylogenetic system of Bibioniformia ) :
|
|
Hesperinus imbecillus, Hesperinidae. Recent. Length 8.5 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. CuA and 1A parallel. |

|
Mesopleciella minor. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 3.75 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. |

|
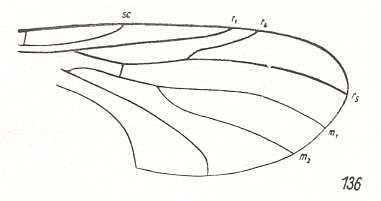
Mesoplecia jurassica. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 7.0 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. |
|
Prohesperinus abdominalis. Upper Jurassic of Karatau, southern Kazachstan. Length of wing 3.25 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. |
|
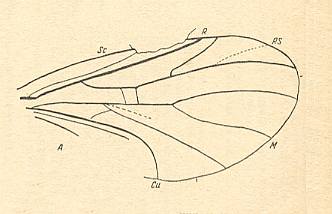
Rhaetofungivora subcostalis. Lower Jurassic of Issyc Kul, central Asia. Length of wing 2.3 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. |
|
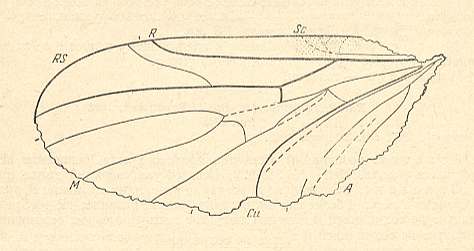
Archihesperinus phryneoides. Lower Jurassic of Issyc Kul, central Asia. Length of wing 2.65 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. CuA and 1A more or less parallel. |
|
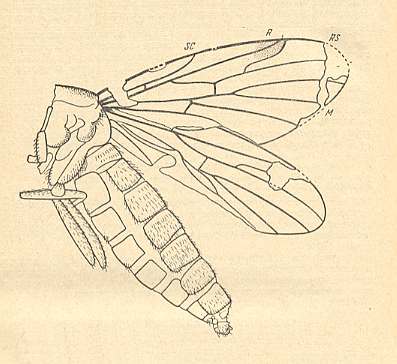
Mesoplecia stigma. Upper Jurassic of Karatau, southern Kazachstan. Length of remains about 8.0 mm. RS 2-branched. M1, M2, and M4 present. M3 absent. Discoidal cell absent. |
The diagram of morphoklines of this parallel series of species is then accordingly :
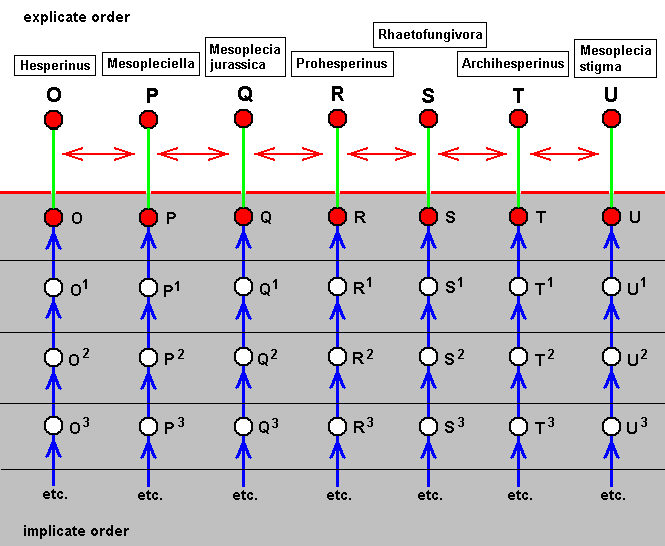
The diagram depicts these seven species as having (in the Implicate Order) independenty developed, that is polyphyletically. Of course one might, on the contrary, think that they have inherited all their common characters from a single ancestor, that is, one might think this collection of species to be (part of) a monophyletic group. And because they are so much alike, their common ancestor must be nearby in terms of time. Of course this supposition is opposed by the fact that the occurrences of these species are scattered over a large period of time : from early Jurassic, via late Jurassic, all the way up to the present time. So it is more reasonable that the common characters of these species have been acquired independently, and at different times. And with those characters the species themselves have evolved independently, although of most of them we do not, apart from their wing-venation, know how their representatives looked like.
Although we here discuss groups of species that are qualitatively more or less closely related [but not necessarily genealogically so related!], species that is, whose corresponding strategies (s.l.) or qualitative contents are more or less similar, a derivational line of such species may nevertheless already reflect very small steps, not of mere change, but of anagenesis, that is, increments of so-called "higher-development" (in the sense of increasing complexification and integration of the body and of behavior, that is, in the sense of an increase of different functions in the organism). This anagenesis will play an important part in our exposition of the general polyphyletic theory of evolution to which we now turn our attention (before, in the next document, continuing with derivational lines and morphoklines in Bibionomorpha).
General Polyphyletic Theory of Evolution
LICHTIG, in his 1938 book on the polyphyletic [all the way down to the species-level] evolution of organisms, "Die Entstehung des Lebens durch stetige Schöpfung" = The origin of life by continuous creation -- not in the theological sense], discusses the difference between anagenesis and genealogy in evolution. In the evolutionary development of the organic world he distinguishes between (1) the "vertical row" which coincides with our "derivational line", (2) the "horizontal row", where such a row is the chronologically ordered row of all living and fossil organisms of the same organizational height or level, and, finally, (3) the "genealogical row", that is the sequence of organismic species that have actually evolved one from the other. The latter more or less coincides with our "morphokline" [which, of course, differs from his genealogical row by the fact that in our theory the descent of organisms from each other does not take place in the physico-biological world (our "Explicate Order") but in noëtic space, that is, in our "Implicate Order" from which, moreover, not all strategies forged there will actually be projected into the physico-biological world.
While evolution proceeds from lower (i.e. simple) forms to higher (more complex) forms, there is also a continuous change within one and the same organizational level, i.e. new forms develop which are different from earlier ones, without, however, having increased their organization.
Let us translate (from German into English) a number of pages (pp.220) from Lichtig's book while supplementing it with commentary and Figures :
From the just given statement, succinctly signified as "horizontal rule", another one follows, which might be called the "vertical rule". To avoid confusion, it is expressly stated that the terms "horizontal" and "vertical" do not refer to, say, geological layers, but are used as terms in natural philosophy to distinguish differences in organizational level. While calling the chronologically ordered row of all living and fossil organisms of the same organizational level a "horizontal row", we will call a "vertical row" a row ordered according to [increasing] organizational level of all organisms simultaneous living in the same definite period of the earth history. Further, every row ordering living and fossil organisms according to genealogic descent may be called a "genealogic row". See Figures.
Horizontal Row
Vertical Row
Genealogic Row
[ If we would rotate the diagrams 900, then the terms "horizontal" and "vertical" could be more easily memorized. We, however, stick to LICHTIG's terminology to avoid confusion with his text. Afterall, these terms refer to organizational height, and a "horizontal row" consists of forms having reached the same organizational level, they organizationally "level out", where "level" is horizontal.]From the "horizontal rule" (reading : there is gradual change even within one and the same organizational level) follows the "vertical rule" : Vertical rows and genealogic rows are incongruent.
This means that if one orders organisms living in the same period according to their organizational level in a row, then this row can never be a genealogic row, that is, its memebers never can be connected by direct genealogic descent.
The way in which the vertical rule follows from the horizontal rule is very simple. Let us take as an example the Mammals that have developed from reptile-like ancestors. The vertical rule then expresses the fact that it is impossible to hold that the recent reptiles are the ancestors of the recent mammals. With "reptiles [still] living today" is meant only the g r o u p s living today, not the individuals. The fact that a given reptile individual cannot be the ancestor of the mammals is evident. The sense of the vertical rule really consists of the fact that the reptile-like ancestors of the mammals are not to be found among the forms -- the reptiles -- still living today , that is, could not have belonged to any the groups Tortoises, Crocodiles, Lizards, and Snakes. And while the most ancient mammals are known already from the Triassic, then their formation from reptiles must have takem place at least at the beginning of the Mesozoic. And were these ancestors of the mammals belonging to recent reptile orders, then these orders must have been preserved from Triassic times up until today. However, according to the horizontal rule (gradual change also within the same organizational level), also within the organizational level of reptiles (the reptile-level) a continuous change of forms has taken place since Triassic times. Consequently, the recent descendants must differ from those reptiles of the Triassic which were the ancestors of the mammals. That this difference turns out to be so big that the triassic and recent forms have to be placed in different reptile orders, cannot, it is true, as such follow from the horizontal rule. But how this may be, all forms of reptiles must have been changed since Triassic times. And that is sufficient to the conclusion that among the recent reptilian forms the ancestors of mammals cannot be found.
Let's diagrammize this, in which F = fish-stage of anagenesis, A = amphibian stage of anagenesis, R = reptilian stage of anagenesis, and M = mammalian stage of anagenesis :
Diagram depicting "vertical rows" (horizontal lines in diagram), "horizontal rows" (vertical lines in diagram), and "genealogic rows" (the four red oblique lines in the diagram). The genealogic rows may be seen as compressed morphoklines.
The recent groups of mammals have not descended from recent groups of reptiles, but from ancestors that looked like reptiles but were more primitive. That is to say, the mammals have, in their evolution, not descended from reptiles, but have passed through a stage that was reptile-like.
And in the same way, the recent groups of reptiles have not descended from recent groups of amphibians, but from ancestors that looked like amphibians but were more primitive. That is to say, the reptiles have, in their evolution, not descended from amphibians, but have passed through a stage that was amphibian-like.
And, again, the recent groups of amphibians have not descended from recent groups of fish, but from ancestors that looked like fish but were more primitive. That is to say, the amphibians have, in their evolution, not descended from fishes, but have passed through a stage that was fish-like.Depicted in terms of noëtics, derivational lines, and morphoklines we obtain :
The derivational line (= "vertical row" sensu Lichtig) F==>A==>R==>M gives us some indication what stages (in the Implicate Order) the recent mammals might have gone through during their noëtic history, that is, what stages figure in their morphokline : a fish-like stage F3, an amphibian-like stage A2, a reptile-like stage R1, and the mammalian stage M (which latter is projected into the Explicate Order).
In the same way the derivational line (= "vertical row" sensu Lichtig) F==>A==>R gives us some indication what stages (in the Implicate Order) the recent reptiles might have gone through during their noëtic history, that is, what stages figure in their morphokline : a fish-like stage F2, an amphibian-like stage A1, and the reptile stage R (which latter is projected into the Explicate Order).
And, finally, in the same way the derivational line (= "vertical row" sensu Lichtig) F==>A gives us some indication what previous stage (in the Implicate Order) the recent amphibians might have gone through during their noëtic history, that is, what stages figure in their morphokline : a fish-like stage F1, and the amphibian stage A (which latter is projected into the Explicate Order).(continuing with Lichtig's text)
The incongruence of vertical row and genealogic row, logically implied by the horizontal rule, can empirically be confirmed. Just as little as the ancestors of the mammals can, the ancestors of birds can be found among recent [groups of] reptiles. The recent amphibians, consisting of Urodela (salamanders), Gymnophiona, and Anura (frogs), are in no way candidates of representing the ancestors of reptiles. And from none of the recent fish the amphibians could have originated.
Were the conventional monophyletic view of the origin of the organismic world right, then vertical rows and genealogical rows had to be congruent, in the sense of :
The vertical row -- which can morphologically be found -- of the recent organisms, ordered according to their organizational height, had to coincide with the phylogenetically-to-be-constructed genealogic row. Precisely this -- i.e. the congruence of vertical and genealogical row -- is the view held in the first days of the victory of evolutionary theory. The systematic series from simple to complex forms normally was interpreted such that the simpler animals and plants still living today were held to be the stemforms of the higher ones, resulting in the whole natural [classificatory] system reflecting the real genealogic tree of the higher organisms. This is just the picture naturally implied by the monophyletic view of evolution. Dürken, it is true, objects to it by the fact that this view is false already by the fact that the ancestors of recent animals must be sought for in those that lived a long time ago, and one should not hold one part of recent forms to be the ancestors of another part of recent forms. This objection does, however, not go to the bottom of the matter. Insofar as it is about the i n d i v i d u a l s, Dürken is undoubtedly right. The transformation of organismes from lower to higher takes a long time indeed, and therefore the lower organized ancestors of the recent organisms must have lived in more ancient times. This is evident, and was apparently also clear to those who which interpreted the systematic sequence from lower to more complex forms of the recent organisms -- which sequence we had called a vertical row -- as to represent a real genealogic row. They, however, assumed that the recent forms of lower organizational height corresponded, as to their qualitative content -- biologically expressed, as to their species -- with lower forms of earlier epochs of the Earth history, which earlier forms are the ancestors of the recent and higher organisms. And that is the decisive point. If one takes one's point of departure with the view that Life has originated from dead matter only once [i.e. only a single time in history] and that the organisms have monophyletically developed from one single ancient living cell, then, because the amount of time at their disposal to evolutionarily develop is equal for all simultaneously living organisms, further development of all organisms of lower organization must have been halted. This stagnation of development must then have ocurred already at that time when higher life-forms developed from them for the first time. When, for instance, already at the latest in carboniferous times the simplest reptiles orginated from amphibians, all amphibians who did not undergo this development halted further development. And then one has to assume that these retarded forms show them themselves in the same form [the same stage] also today as they were already in carboniferous times. This thoroughly natural assumption was held by all who took the morpho-systematic row of recent organisms ordered according to organizational level as a genealogic series.
So what was to be expected in the monophyletic view is the contrary of what the horizontal and vertical rules say to us in reality. Both rules are utterly opposed against the idea of depotentialization, which in the monophyletic view would naturally follow as an explanation of the fact of the contemporaneous existence of the most diverse complexificational stages in the organismic world.
See next Figure.
Phylogenetic development of mammals from certain reptiles according to monophyletic theory. That is, some reptiles have (gradually)changed into mammals, while others remained unchanged.
[ We, on the other hand, hold that evolutionary development takes place, not in the Explicate Order, but in the Implicate Order. In this view primitive forms (forms of low organizational height), initially having become extinct, may, much later, be projected again into the Explicate Order when appropriate ecological conditions reappear.](Continuing with Lichtig's text)
Another important phenomenon, which was, from diverse sides, already rightfully brought against monophyletic theory, consists in the fact that, if one one looks back in time, one does not see a steady decrease in organizational height of the organisms, while according to monopyletic theory such a decrease can be expected. It is true, the picture of the nature of fauna and flora does change the more we descend into history. The forms we see there are more different from recent ones the more ancient are the geological layers from which they are taken. However, according to monphyletic theory it is to be expected that the organizational height of living beings has steadily increased since the single event of the origin of Life, so that, when descending into history , one would not merely see things changing but especially see a gradual decrease of organizational height of living beings. In fact, however, already in the oldest fossil-bearing deposits all animal phyla, except the vertebrates, are represented. The only new event since these most ancient times, new in the sense of the appearance of higher organization levels -- anagenesis -- is the origin of vertebrates, and in the plantworld that of the terrestrial plants.
[ So apart from the eventual appearance of the vertebrates as the highest stage of anagenesis, we cannot, in these ancient times, see a gradual increase of organizational height in the living beings of these times.] [ Also in insects, all the main types were already present early on. See HERE ]
According to monophyletic theory one should further expect to find in the older deposits not yet specialized original forms (archetypes) which must be seen as the starting forms of the forms later to appear. The assumption of archetypes is very significant in monophyletic theory. Mammals are supposed to have evolved from some archetypic mammal, birds from some archetypic bird, reptiles from some archetypic reptile, amphibians from some archetypic amphibian, molluscs from some archetypic mollusc, etc. Such an archetypic form, if it is the ancestor of all later representatives of the group, must fail to possess all special characters that distinguish individual divisions of the group from other such divisions. An archetypic mammal, for example, may not have possessed any of the one-sided specializations of a monotreme (duck-billed platypus), of a marsupial, or of a placental mammal. It rather must be in such a degree undetermined that all the special [later] forms could still originate from it. In reality, however, we don't see such archetypic forms. The life-forms are already in ancient times precisely as specialized as they are today. The Trilobites of Cambrian times, as to their specialization and organizational height, are not inferior to the modern representatives of Crustaceans, and even significantly surpass many of them. [It must further be realized that genuine archetypic forms as they are constructed by phylogeneticists do not seem to be viable, that is, they do not seem to represent true strategies to exist.]
Still more clearly, however, difficulties appear, difficulties speaking against the conventional monophyletic theory, in the way in which Phylogenetic Systematics must draw genealogic trees of organisms designed by it. To best understand the essentials of such constructed trees one should distinguish between that what one expects and that what is obtained from experience (i.e. observation). From the monophyletic view one would expect that orgamisms can be arranged in a straightforward genealogic tree in which always the group that is placed higher up in the tree originates from the group placed below it. However, the genealogic trees actually drawn -- although also they are hypothetical, depicting the real state of affairs only schematically -- generally show parallel lines, that only deep down begin to converge a little, and eventually come together in a single point far down in the depths of history. As an example here the genealogic tree may be given that Handlirsch, 1906-1908, has designed of the beetles (Coleoptera) :
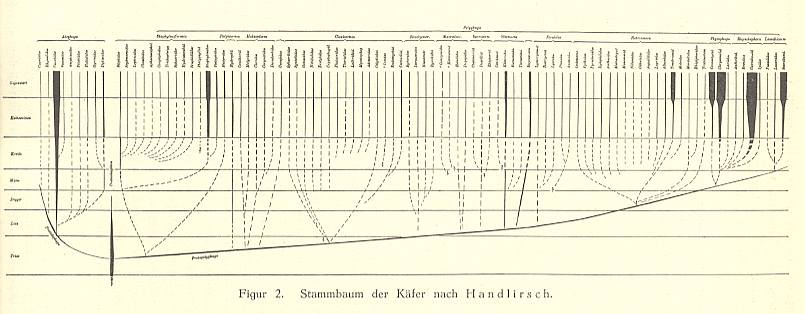
Genealogic tree of the families of the insect Order Coleoptera. The geological periods are, reading from top to bottom : Present, Tertiary, Cretaceous, Malm (upper Jurassic), Dogger (middle Jurassic), Liassic (lower Jurassic), Triassic.
Although Handlirsch arranges the families in a typological way (and thus not according to strict phylogenetic systematics) the tree is definitely meant to represent genealogical relations.
We see in this genealogic tree a large number of parallel lines, further down ending up in dashed weakly converging lines. What does this picture mean, and what is its bearing on the question of monophyly and polyphyly? The continuous straight lines are the stem-lines of the various beetle families, as far as paleontology and morphology are able to determine things. The dashed lines, on the other hand, are hypothetical completions of these stem-lines backwards. The common point of intersection of all the lines,- which in the diagram -- evidently on technical grounds, because it would fall far outside the picture frame -- is represented by a more or less horizontal continuous curve in which the various lines connecting individual points of intersection come together,- represents the hypothetical archetype, that is the archetypical beetle, from which all beetles are supposed to have evolved.
Relinquishing all that is hypothetical, and limiting our consideration to the facts -- which are in reality, as has been said, because of the nature of phylogeny [taking very long time], also already in a high degree hypothetical -- we see incomplete rows (ancestral lines) in the phylogeny of Coleoptera. We never see complete ancestral lines, because these should have to reflect the complete beetle-genesis, beginning with the concerning origin-of-life-act, and then all the way up to the origin and development of beetles. What we are able to determine are always no more than mere sections of phylogeny which can be traced back to a certain point, after which things become foggy. And here hypotheses begin, making up the uncomplete picture. And because most phylogeneticists adhere to monophyly, the hypothetical completion will proceed in monophyletic fashion, where one assumes that the ancestral lines, after the point where they cannot be traced farther back, converge and eventually will meet one another in some hypothetical point of convergence. If one removes from this picture all things that were hypothetically added, then polyphyletic rows remain that run parallel to each other. It must be noted that in reality also these individual straight lines are in fact seen to be composed of many parallel lines when instead of families, as is the case in Handlirsch' drawing, the genera, or even the 300000 species, were displayed in this picture. [ . . . ]To this we may add two more ancestral trees, one of the Order Diptera, and the other of Hymenoptera, both published in 1980. To see them click HERE .
(Continuing with Lichtig's text)
What has been expounded with the help of the example of beetle-phylogeny [and also of dipterous and hymenopterous phylogeny] holds for every phylogenetic genealogic tree. [ . . . ]Generally in science it is such -- and we have also seen this in the science of Phylogeny -- that a theory, generally held to be correct, wil not easily be abandoned when the expectations inherent in that theory are not confirmed by empirical data. One rather attempts to accomodate the theory to these data by changing it in some respects, i.e. by adding special assumptions. And this is also done in the present case (where phylogenetic lines appear to be parallel). Different approaches have been taken which will be further elucidated.
[ It is important to know that Lichtig, when dealing with monophyly, polyphyly, origin, derivation, etc. has in mind mainly the larger animal and plant taxa, i.e. the higher taxonomic categories such as Orders and Classes. And this is understandable because the phenomenon of "higher-development" (anagenesis), and thus true evolution, is best detectable when following the fate of these larger groups. Further we know that Lichtig, as do all biologists, accept only one Order of Reality, the Order that we have called the Explicate Order. So in his view phylogenetic development, and all other processes for that matter, takes place in the Explicate Order, while in our view it [i.e. phylogenetic development] takes place immaterially in the Implicate Order. Lichtig further assumes that not only the origin of species is polyphyletic but also the origin of Life itself, implying that the event of the origin of life has occurred many times and in many places, where each such origin gives rise to a 'vertical' unbranched genealogical sequence of ever higher developing forms. And the cause of anagenesis he finds in the continuous action of the environment exerted on every organismic individual from generation to generation, forcing new functions and adaptations to arise. In our view the origin of life takes place in the Implicate Order. And if it took place there repeatedly, the acts of origin must be qualitatively different from each other as well as their results [because repetition of qualitatively the same cannot be in the Implicate Order]. The mentioned results [of the acts of origin of life] are the most simple strategies for immaterial patterns to exist (as material organismic species) in the Explicate Order (and to obtain ontological completion). In fact they can hardly be called strategies at all, but look rather like snow-crystals whose particular shape and size guarantee their more or less sustained existence in given meteorological conditions. The more complex immaterial patterns, on the other hand, must turn into real strategies in order for them to be able to exist in the Explicate Order. They do not correspond to the origin of life, but to the products of evolution.]
(Continuing with Lichtig's text, concerning changes that have been made in monophyletic theory, mainly to remedy difficulties caused by specialization-crossings : (1) the alleged original (primitive) state of a character --rendering it non-derivable -- is assumed to be secondary, and thus being derived character state afterall, (2) backwards projection of point of convergence of phylogenetic lines, (3) hypothetical mediating forms.)
A chief difficulty often opposing the straight phylogenetic connection of organic forms as this is expected in monophyletic theory, is the specialization-crossing. As soon as in two forms one of them in one character, and the other in another character shows a higher development no one of them can be the ancestor of the other. This holds at least as long as one supposes that the higher organization of a character is a derived condition with respect to the simpler organization, because generally the more complex originates by a transformation of the simple. It is, it is true, also conceivable that the simple has resulted by reduction of the complex. In this case the simple condition of a character is a derived condition, derived that is, from a complex condition, beause it has secondarily evolved by reduction of the latter condition. Thus, winglessness in an insect may be primary (i.e. original) in the sense that its ancestors never had wings. There are, on the other hand, also cases of secondary winglessness, having resulted from the fact that the winged ancestors have lost their wings. Here we thus have the possibility of two interpretations [original condition, derived condition]. Because every complex organization ultimately must have originated from a less complex organization, every reduction means a zig-zag course of evolution. [ This course refers to the evolutionary fate of single characters, not to species. When an insect evolutionarily loses its wings, this is part of an overall transformation of it with respect to other morphological characters as well as to its behavior, meaning that the resulting overall strategy may nevertheless have become more complex.] In the mentioned case of secondary winglessness phylogenesis proceeds from an originally wingless state to a winged state and from there back to a wingless state. When, accordingly, in this way a simple condition is not necessarily always original, a difficulty as a result of specialization-crossing [species A is simple with respect to character X as compared to that of B, while B is simple with respect to character Y as compared to that of A] may be solved by assuming an alleged simple character, [say X of species A] to be merely seemingly simple, while in fact it is derived [now allowing A to be derived from B]. If it is, for example, impossible to consider a given wingless insect as a descendant of some winged insect as long as one considers winglessness as the original condition -- because then one would have the impossible result that the descendant is, as to its winglessnes, more primitive than its alleged ancestor --, the case directly changes when one considers the winglessness of this particular insect to be secondary. Now the present case of winglessness is a derived condition and presupposes a winged ancestor.
It is known that reductions abound in phylogenesis. They can, however, always be explained biologically. If it is only a single organ that has been reduced, one may assume that the cause is to be found in an altered way of life in which the organ is not used anymore and therefore disintegrates. If, on the other hand, the reduction involves essential parts of the whole organism, then the biological cause generally lies in the transition from a free-living mobile way of life to a sedentary or parasitic one. In all cases the zig-zag course of phylogeny, as a result of reduction, is rather rare, and must therefore be especially proven for every individual case. Generally one may hold that the simple condition is at the same time the original condition.
The way in which difficulties, coming from paleontology or morphology, to phylogenetically connect groups, are being solved by assuming a phylogenetic zig-zag course, is shown in the phylogeny of the lowest vertebrates. The most ancient fossil vertebrates are the Armoured Fish, Ostracodermi, known from upper Silurian and Devonic deposits. The investigations of Stensiö have shown that the oldest Ostracodermi have possessed, in their exterior skeleton as well as in their interior skeleton, well-developed bone tissue. On the other hand, in elasmobranchs that for the first time appear in Devonic times, typical bone tissue is not present. Because of this it is apparently impossibe to derive the elasmobranchs from armoured fish, because armoured fish already stand -- as to their possession of an ossified interior skeleton -- above the elasmobranchs. In order to establish here a genetic connection, Stensiö assumes that also the ancestors of the elasmobranchs did possess a bone skeleton, and that its absence in elasmobranchs has resulted from a secondary reduction, in which the bony skeleton was replaced by a cartilaginous one.
We will not consider the difficulties that oppose the assumption of Stensiö, which, for example, have led Sewertzoff to dismiss as improbable the hypothesis that the bony skeleton of the vertebrates was already developed very early on in phylogeny and was secondarily lost in elasmobranchs. The view of Stensiö has been given here merely as an example of the art of constructing ancestral trees, which may be indicated as the method of assuming a phylogenetic zig-zag course. Would one, however, apply this method in all cases where a phylogenetic connection of morphologically related forms runs into difficulties, the phylogenetic zig-zag course would be the prevailing one in the phylogeny of organisms. In fact even then not all difficulties can be resolved, and the fact of parallelly running phylogenetic rows will also in this case remain. As a result one has developed an additional way of handling things which may be called the method of backwardly projecting points of convergence.
While initially one had assumed that groups ordered according to organizational height may directly be connected with each other phylogenetically, and had as to one's astonishment on scrutinity found out that this is often impossible, one later assumed that in such cases both groups were descendants of a common ancestor. This common ancestor was then the point of convergence in which the genealogic lines, that could not directly be connected with each other, came together, after having them projected backwards in time. For example, the original assumption was that the elasmobranchs were considered to be the ancestral group of bony fish and tetrapods. But already the fact that the completely different way in which the vertebrae are formed in elasmobranchs and bony fish excludes a direct genealogic connection between these groups, had forced investigators, adhering to the monophyletic view, to assume that elasmobranchs and bony fish have a common ancestor. But a definite recent or fossil animal group has, as in so many other cases, not been found, and thus the point of convergence remains hypothetical. As a result of newly discovered ever more specialization-crossings one was forced to project the point of convergence of many morphologically closely related groups farther and farther backwards, often so into infinity. In addition one is often forced to combine this method of backward projection with the method of phylogenetic zig-zagging, in order to resolve difficulties. Thus, for instance, the arrow-worms, Chaetognatha, are difficult to connect with any other animal group. Therefore Naef held that they are descendants of very primitive archetypical Coelomates in which the archetypical coelomate would be the hypothetical ancestor of all Coelomates, i.e. of all animals with a body cavity. So in this case it is not sufficient to shift the point of convergence of Chaetognatha and the rest of Metazoa back to the hypothetical archetypic Coelomate. One has, moreover, to degrade the archetypical Coelomate considerably, i.e. let it be reduced with respect to various characters, in order to establish a connection with the Chaetognatha.
Finally, we must mention a third method which can be called that of the hypothetical mediating forms, and which in many cases also is combined with one of the other methods. This method is chiefly applied by morphologists. While it is for the more paleontlogical phylogeneticist chiefly a matter of finding phylogenetic relations between actually known fossil or recent organisms, the morphologist first of all wants to know how exactly the ancestor of some given group might have looked like. If he then has, as a result of morphological comparison of relevant fossil and recent organisms, obtained a certain picture of the archetypical form assumed by monophyletic theory, without, however, finding some definite group of organisms satisfying that picture, then he is inclined to posit hypothetical organisms for this morphologically determined archtypical form. In order to get rid of difficulties resulting from specialization-crossings and other morphological phenomena, it often is, however, not enough to establish the phylogenetic connection between several groups of organisms by means of only one single hypothetical archetypical form. Rather one is forced to assume more than one hypothetical forms,that is, mediating forms, in order to make possible a morphologically moderately satisfying derivation of the groups from one another.
The next diagram makes clear what Lichtig means here.
All mentioned methods have as their goal to accommodate facts established by morphology and paleontology with the idea of a monophyletically proceeding phylogenesis. So things show themselves such that the actually established phylogenetic rows run parallel, i.e. polyphyletically, while the hypothetical completion in the range of the not-empirically-determinable-anymore proceeds in the monophyletical sense. Monopyletic theory is considered to be in such a degree self-evident that only occasionally one doubts the legitimacy of this way of seeing things. It is true, there are, apart from investigators that do adhere to the polyphyletic theory, also adherents of monophyletic theory who hesitatingly ask the question whether the current practise of monophyletic connection of groups of organisms is right and whether it may not be better to replace it by the assumption of a polyphyletic origin of form-groups. But generally one is afraid to do so. [ . . . ]
But one may, of course, simply take the view that it is right to actually take serious the conclusions resulting from scientifically demonstrated facts, whether they do conform with current beliefs or not. We have seen that many characters in plants and animals must have been polyphyletically evolved.
Zimmermann, it is true, rightfully distinguishes between polyphyletic character development and polyphyletic genealogic development. We have to do with polyphyletic character development when a given character, for example the gymnospermian way of reproduction in plants, has originated independently in different genealogical lines. With polyphyletic genealogy, on the other hand, we have to do when the whole development of a group has taken place in independent genealogic rows, each of which goes back to its own act-of-origin-of-life. In fact, however, the difference is merely quantitative. Every organic form is composed of a number of characters, and when for some characters it is found that they have originated polyphyletically, it is not quite understandable why this would not hold for the other characters as well, resulting in similar organic forms [species] having originated independently. Indeed there are cases in which this can even be demonstrated. So, in all cases, as a result of the discovery of polyphyletic character development, the whole view of the monophyletic theory that the transformation acts in organic forms are one-off events is destroyed, and with it the philosophical foundation of this theory.
What the science of Phylogeny has shown us is that those sections of the historical development of organisms that can actually be uncovered by it give us the picture of parallel rows. Departing from the monophyletic view, these polyphyletic rows were then, with the help of the above described methods, completed in the form of converging rows. Therefore the hypothetical completion deviates in a decisive point from the demonstrable part of the course of phylogeny, because in the hypothetical part convergence in genealogic lines is assumed, while in the demonstrable part no convergence at all can be detected. So it seems more natural to hypothetically complete the unknown part of phylogenesis in the same sense in which its demonstrable part goes its way. And with this one arrives at the method of recurrent rows, as is illustrated in the diagram below [ In this diagram of Lichtig (p.233) all transformations, leading to certain organizational stages in organisms, take place, of course, in what we call the Explicate Order] :
Diagram above : Scheme of recurrent rows, displaying the consecutive organizational stages A, B, C, D, E, F, G, and H. The scheme expresses, not the phylogeny of groups of organisms, but the ascend of Life to higher and higher organizational levels. Indeed, in polyphyletic theory there are no g r o u p s of species (higher taxa) that evolutionarily develop, but the species themselves, all in parallel lines. Higher taxa here are not genealogic groups but collections of species that are similar qua qualitative content in the horizontal dimension of the present diagram. So, for example, g1 is simlar to G, c3 similar to c2, etc. This similarity has not resulted from genealogic descent, but is based on qualitative content.
See for (the dimensions) "vertical row", "horizontal row", "organizational height", and "genealogic row" the considerations and figures above. [ these dimensions are added by me to Lichtig's diagram]
The parallel vertical line segments in the upper rectangles signify lines of genealogy that are actually empirically detectable. Below each rectangle one should think them to be extended by dashed lines, expressing the not empirically detectable phylogeny. Each such vertical line (continuous + dashed), running back along the time-dimension, ultimately ends up (in fact begins) at a particular origin-of-life-event. In Lichtig's polyphyletic theory these origin-of-life-events have occurred at many times in history (and still taking place today) and also at different places. The next version of this diagram expresses this.
In order to be able to depict (1) the fact that the "vertical row" must stand at right angles to the time-dimension, (2) the "horizontal row" must be in the time-dimension, and (3) the dimension of (increasing) organizational height must also be in the time-dimension, we consider the time dimension to be analyzable into two components, as can be done with other vectors such as forces. Whether this is physically legitimate I'm not sure. But that is immaterial here, because we only say that, for instance, the horizontal row signifies a series of forms standing at the same organizational height but occurring in different times. (After LICHTIG, 1938, slightly changed)Diagram above : Second version of the above diagram. The origin-of-life-events occur at different times, therefore, as seen from the present, to some line-bundles more time was available to increase organization height than was to others.
The sequence A==>B==>C==>D==>E==>F==>G==>H of the stages A, B, C, D, E, F, G, and F corresponds to the "derivational line" of species in our theory. And indeed, for us, as well as for Lichtig, (the organizational stage) H has not evolved (and is not derivable) from G, but from g1, which is similar to G and has the same organizational height. In the same way, G has not evolved from F, but from f 1 which is similar to F and has the same organization height. (see also the first version of this diagram ).In the ABOVE SCHEME the letters A, B, C, D, etc. signify consecutive organizational stages. The highest stage H may, for example, represent the angiosperm stage in the plant kingdom [it is a stage of the way of reproduction]. The continuous parallel lines signify the genealogical lines leading to the next lower organizational stage, in our example thus to the gymnosperm stage. One knows that the Angiospermae have polyphyletically developed from Gymnospermae. It is, however, impossible to point to a particular Gymnosperm group from which the Angiosperms could be derived. And of course it is still less possible to trace the phylogeny of the Angiosperms farther back into history. What Phylogeny is actually able to demonstrate is, that, and how, the angiosperm way of reproduction may have evolved from the gymnosperm way of reproduction. At stage G, which is in our example the gymnosperm stage, the continuous parallel lines again are the polyphyletic rows, leading from the gymnosperm stage to the next lower organizational stage, the pteridophytic stage (F). Also here one cannot point to a definite pteridophytic group to which one can connect the gymnosperms. If one continues this reasoning with the next lower stages, then one will obtain just parallel rows, leading from one stage to the other [and thus not from one group (taxon) to another]. The upper staircase-like part of the diagram represents the empirically determined part of the phylogeny, insofar as in such a heigh degree hypothetical issue one can speak at all of empirical determinations. The rest of the diagram represents the part in which the course of phylogeny must be hypothetically completed by us. When we are attempting to obtain a picture telling us how phylogeny of the unknown gymnosperm-like ancestors in the section (rectangle g1) from the pteridophytic stage to the gymnosperm stage [thus the phylogeny from the bottom of rectangle g1 to its top] went its way, then the analogy with the empirically determined course of the phylogeny of the known gymnosperm groups [ the continuous lines in the rectangle G ] suggest also here [i.e. in rectangle g1 ] to assume parallel lines. [ So we now have 'done' the rectangle below H, and the next lower rectangle (f2) can be done with the help of the empirical findings in the pteridophytes (F), and so on ]. Continuing this way of doing things with the remaining organizational stages (G, F. E, D, C, B, and A) we obtain a series of parallel lines, extending all the way down (in time) to their respective origin-of-life-event. And this is the picture one obtains when one completes the hypothetical part of the phylogenesis according to this method of recurrent rows, i.e. completes it in the same way as the empirical determined part in fact is.
It is remarkable that also the picture of phylogeny designed by monophyletic theory necessarily ever more approaches the picture just given. We have seen how one, as a result of more and more precise morphological and paleontological investigations -- especially by the implied need to exclude form-groups from an initially surmised direct phylogenetic connection as a result of discovered specialization-crossings, or when one has to do with large temporal gaps -- was forced to project hypothetical points of convergence, in which the genealogic lines, followed down into the past, are supposed to come together, farther and farther back in history. But this means that also according to monophyletic theory the genealogic lines are only weakly convergent [ See, for instance, again HERE ]. Translated phylogenetically, this means that large sections of phylogeny have proceeded independently along individual genealogic lines, and that with it also many common characters of these lines must have originated independently. If one holds, with Naef, the arrow-worms, Chaetognatha, to be the descendants of still strongly reduced [i.e. very primitive] archetypical coelomates, then it follows that, for example, the similarities between arrow-worms and lancelet must have originated independently. If one goes stil further with backward-projecting points of convergence and places them into infinity itself, then one obtains the picture that conforms with polyphyletic theory, because lines that meet in infinity are consequently parallel. The point of infinity here means the abiotic [i.e. the inorganic], that is the stage in which life originates from dead matter. Accordingly, the polyphyletic view essentially is just the logical implication of ideas to which also monophyletic theory was forced to adhere, after it has abandoned its original view that the representatives of the different organizational stages could directly be connected genealogically with one another, more and more in favor of hypothetical common ancestors.We have seen -- and in this point all authors do agree -- that in the phylogeny of living beings often the same characters appear in different series independently of one another, as for instance, the gymnosperm way of reproduction in plants, and organs of vision in different groups of animals. By making a causal analysis of these important phenomena of polyphyletic acquisition of characters it will become possible to penetrate to the chief question that is essential to the problem of monophyletic or polyphyletic descent, the question, namely, of precisely what relationship exists in organisms between affinity of essence based on similarity of morphological structure or of qualitative content, and genealogic affinity [affinity of genealogy] as a result of common descent.
[In the context of our noëtic theory we should realize that in the Implicate Order "affinity of essence" is identical to "affinity of genealogy", because here the latter is nothing more than affinity of noëtic derivation.]. But before we do so, i.e. making the just mentioned causal analysis, we first look into the phenomenon (to be analysed) itself of polyphyletic acquisition of characters a little more closely.
Polyphyletic development of characters is demonstrated in plants and animals so often, that it is impossible to give more than a small selection from the large number of examples. Usually in it one distinguishes between parallel and convergent development, that is, whether in the different genealogic rows the character is similar already from the beginning, resulting in the same end-state by similar transformations in the different rows, or that the same character has developed in the different rows from different starting-states in different ways. [Parallel development of a (complex) character in different genealogic lines involves homologous structures, so that we can speak of homologous polyphyly. In fact this then is a precise repetition of a given transformational series taking place in different genealogic limes. On the other hand, convergent development of a (complex) character in different genealogic lines involves non-homologous structures, and it might be called analogous polyphyly. In fact we accept only homologous polyphyly as representing a truly polyphyletic development of a given character. Analogous polyphyly only points to the repetition of very general traits or tendencies, such as the polyphyletic development of vivipary in plants and animals.] As an example of parallel development in the plant kingdom we can point to the transformation of the reproductive organs from an isosporous condition to a heterosporous condition, and from there to a gymnosperm condition, and finally to the angiosperm condition, while as an example from the animal kingdom may serve the transition of the suture in different groups of Ammonites from the goniatite-stage to the ceratite-stage and from there to the ammonite-stage, and, as another example, the transformation of scales in the fish-group of Actinopterygia, which in different independent rows has led from the most primitive stage, that of ganoid scales -- rhombic scales with ganoine cover -- to the next higher stage, that of cycloid scales -- scales with a hemicircular posterior margin -- and finally to the highest stage, that of ctenoid scales -- scales with a serrated posterior margin.
As an example of convergent development of characters in the plant kingdom may serve the origin of piercing organs in plants, which in different plants are formed from morphologically quite different parts of the plant-organism. Thus, for instance, in Acacia albida from transformed side-leaves, in Prosopis stephaniana from parts of the epidermis [the skin], in Halimodendron from hardened petioles, in Alhagi, Balanites, and Eversmannia from languished shoots, etc. A convergent development of a character in animals is, for instance, the formation of organs of flight in insects, reptiles, birds, and mammals.
[In Phylogenetic Systematics one does not consider convergent development (in the sense of development of a character from analogous beginnings), because their uselessness in phylogenetic systematics is obvious. Its interest solely lies in saying something about general tendencies in evolution. So in Phylogenetic Systematics only the development of a character from homologous beginnings is considered. And when one, nevertheless does speak there of convergence, then one means the homologous polyphyletic development of a given character, in contrast to a monophyletic development.].
In what follows it is not distinguished anymore between parallel and convergent characters, while this distinction is not important with respect to the problems here considered [ Indeed, here it is only important to show that nature has evolved morphologically and/or functionally similar structures independently from one another, whether they are homologous or analogous]. [ . . . ]
The forms of reproduction in plants, that show the stages of Isospory, Heterospory, Gymnosperm, and Angiosperm, have polyphyletically evolved. The consecutive order of stages, here indicated, is characterized by an increase of care of the offspring. And here we see, to which Zimmermann rightfully points, a notable parallel with vertebrate phylogeny with their end-members, the birds and the mammals [and in insects in termites, ants, higher wasps an bees (care for the young), and in Glossina (tsetse-fly) and Sarcophaga (carrion-fly) (vivipary) ]. In this context it is remarkable that also vivipary parallelly occurs in (certain) animals as well as in (certain) plants. And in both divisions it has polyphyletically originated. In the plant kingdom vivipary is found in several species, not related to one another, of the mangrove-vegetation. Among animals, as is well-known, it is to be found in mammals. But vivipary also occurs in fishes -- for example in the smooth-sharks Mustelus and Carcharias --, in amphibians -- for example in the black earth salamander Salamandra atra --, and reptiles -- for example in the bog lizzard Lacerta vivipara --, and is even found in invertebrates, in the most diverse groups. These very remarkable correspondences between the animal and plant kingdoms demonstrate quite clearly that these are not a matter of coincidence, but of law and pattern. And precisely because they are regular, they are not one-off events, but everywhere occurring when certain conditions are satisfied. [ . . . ]After having enumerated many examples of polyphyletic development, Lichtig (pp. 265) continues :
In all enumerated cases one first of all assumed that a morphologically determined group [i.e. species put together on the basis of their possession of common morphological characters -- morphological kinship] also expressed phylogenetic kinship [of its members to one another]. Only subsequent investigations demonstrated that the morphological correspondences in individual characters are opposed by essential differences in other respects, and that therefore the apparent uniform group has originated polyphyletically. Nature had, as it were, confused people by giving them the appearance of phylogenetic kinship where in reality it was merely convergence. One did, however, not conclude from this that the very method to see kinship-of-form as evidence of kinship-of-descent is false, but assumed that there are characters which have polyphyletically or convergently evolved and which therefore cannot be evidence of phylogenetic kinship, while other characters had to be seen as being inherited from a common ancestor, and thus may constitute evidence of phylogenetic kinship. From this viewpoint one may in organisms distinguish between two groups [types] of characters. On the one hand we have the characters that originate in different lines each for themselves independently of any phylogenetic kinship, and are indicated as adaptational characters, because they originate as a result of parallel adaptation to the same way of life. [ . . . ]
On the other hand we have the organizational characters, characters that are necessarily implied by the intrinsic organization of given living beings, and inherited from a common ancestor, resulting in the fact that all forms [species] possessing such a character are also genetically uniform.
To recognizing phylogenetic relationships within the conventional view the distinction between adaptational and organizational characters, or, shortly, between A- and O-characters, is paramount. A large part of disagreements on phylogenetic relationships boils down to the question whether given characters are A- or O-characters. [ . . . ]
One can, without exaggeration, say that the distinction between A- and O-characters really is the "A and O" of the whole of Phylogenetic Systematics [In HENNIG, 1954 (phylogeny of Diptera), and 1969 (phylogeny of Insecta) we see that such a distinction is still made, but that it is made dependent on the particular group of insects considered. This introduces a certain degree of arbitrariness, especially when there is little help from other characters in the group of organisms studied to non-arbitrarily decide whether a given feature has originated convergently or that it has been inherited from some common ancestor. Often, in Phylogenetic Systematics (phylogenetic taxonomy) sensu Hennig, monophyletic origin of a given character is assumed on the basis of its high degree of complexity, forbidding it to originate more than once independently. But in things like wing-venation (which is phylogenetically used especially in Hennig 1954) we don't have to do with such high-degree complexities.]. Repeatedly one was forced to made the discovery that some given character held to be an O-character turned out to be only an A-character. Every phylogeneticist believes that the characters he has used to establish phylogenetic relations are O-characters, while his opponent has erroneously taken A-characters as being O-characters. It is here like in the parabel of Boccaccio about three rings, where each son believes that he possesses the real one while his brothers possess only a fake of it. But that leads to the question what exactly the difference is between the genuine and the fake, or, in our case what exactly the biological difference is between A- and O-characters. How can we recognize a given character to be an A- or an O-character?
Because in the conventional view systematic classification should reflect phylogeny, only O-characters may be used to establish systematic categories [ in Hennig, these are monophyletic groups that can be demonstrated only with derived (apomorphous) characters]. Therefore the evolutionary taxonomist would be pleased to have at his disposal a means to recognize and to pick out from the manifold of existing characters the O-characters. Experience, however, shows us that such a general means of recognition does not exist. What in one group may legitimately be used as O-character is in another group a taxonomically useless A-character or vice versa [ In Hennig's taxonomy these 'useless characters' are the non-derived (plesiomorphous) characters, not non-derived in an absolute sense, but non-derived within a given group of organisms]. RENTSCH (1934) makes up the Golden Rule for the taxonomist looking for a universally valid criterium of legitimate taxonomic characters : There is no such rule.
To precisely the same conclusion one will arrive also by theoretical considerations. As A-characters or [equivalently] adaptational characters we take those characters that originate as a result of the continuous indirect or direct impact of the environment, that is, those characters that usually are marked as adaptational features. Rightfully, however, Dacqué notes that all features that we can detect in the concrete organism, all the way into its finest details, are adaptational characters [the totality of which we have called the strategy s.l.]. And this implies that we must consider all characters of a living being to be A-characters. No character exists of which its formation cannot in one way or another be reduced to the influence of environmental conditions, which [characters] can therefore be formed convergently in the most diverse rows whenever those conditions are satisfied.
The whole distinction between A- and O-characters, as it is typical of phylogenetic research in the conventional view, breaks down. So it is not surprising that in looking for a criterium of O-characters for such a long time no such criterium has been found. An O-character, a character that is not biologically determined by the environment, and therefore legitimately indicating genealogical kinship, does not exist [This is in need, however, of more thinking and re-thinking, especially when we consider features like family-groundplans]. Because it was common practise to conclude from the similarity of organismic forms with respect to a common possession of certain characters, insofar as these characters were considered to be O-characters, to the common descent of these organismic forms, it now becomes necessary to test our ideas concerning the relationship between qualitative kinship and genealogic kinship as to their truth.
Kinship of form [= kinship of quality] means similarity with regard to form [= similarity with regard to qualitative content], in which here the concept of form is taken in its broadest sense, and therefore including color, histological structure, [behavior], etc., in short, the whole essence of the thing having this form, meaning that one can also speak of kinship of essence [or kinship of intrinsic qualitative content]. Kinship of genealogy, on the other hand, means descent from one another or from a common ancestor. So it is clear that kinship of form and kinship of genealogy are two entirely different concepts. In the literature on evolution one usually just speaks of kinship as such, without clearly distinguishing between the concepts of kinship of form and kinship of genealogy. This way of handling things would only be legitimate when kinship of form always coincides with kinship of genealogy, meaning that there would be no genealogic kinship without qualitative kinship, and no qualitative kinship without genealogical kinship. In that case "kinship" would at the same time mean kinship of form and kinship of genealogy. In the days directly following the victory of the theory of evolution one believed that this supposition was actually true, and saw every kinship of form as a sign of kinship of genealogy. But the discovery of convergences has changed all this. Similarities in form that were the result of convergences were now spoken of as merely apparent or superficial similarities [extrinsic similarities], contrasting with kinships as a result of intrinsic similarities. But in reality in case of convergence the similarity is neither merely apparent nor extrinsic, rather we here have to do with a genuine intrinsic kinship of form. It is only the kinship of genealogy that is apparent, kinship of genealogy that is simply concluded from kinship of form. The legitimacy of so concluding, in theorizing about organic evolution, often still considered to be an evidently presupposed way of doing things, must now of course be determined. [In all this it is important to realize that indeed kinship of form is given, that is, can be directly observed empirically. On the other hand, phylogeny cannot be observed (because the times involved are much too long), but only c o n c l u d e d from something that can be observed, such as kinship of form. But in determining the presence of kinship of form one should decide between analogy and homology. And in kinship of form it is only the similarity of h o m o l o g o u s forms (structures) that counts. But homology of forms is nowadays interpreted as the forms being more or less directly historically connected, such that their bearers have a common origin and thus also the forms and structures of them. And this demand of historical continuity between forms supposed to be homologous automatically lets true kinship of form directly imply kinship of genealogy. So it seems that the two concepts -- kinship of form and kinship of genealogy -- indeed do coincide. This is plain thinking isn't it?. But wait a minute. The concept of homology was defined already before the advent of evolutionary theory in the sense of Darwin. So what then was the (original) meaning of homology? The concept was found by the rational morphologists and they evidently meant by "homology of forms" their rational derivability from one another, as is, for example, the skeleton of a bird's wing qua structure and form mentally derivable from that of the fore-limb of the reptile. And all this without supposing actual descent! (as we see it [only] of individuals of a given species). And indeed, in our noëtic theory of evolution d e r i v a b i l i t y is an existing relationship between immaterial forms in the Implicate Order, and on the basis of this property of derivability we may say that in the Implicate Order forms have been d e r i v e d from one another, without, however, letting the term "derived" to refer to some process actually taking place or having taken place in the Implicate Order.]
If one wants to enquire into the relationship between kinship of form and kinship of genealogy it is wise to divide the problem into two individual questions. The first question reads : Is all that is genealogically related also qualitatively related? The second question reads : Is all that is qualitatively related also genealogically related? In this we need not confine these questions to the realm of organisms but may also investigate them in the realm of inorganic things, resulting in a deeper understanding of the essence of the relationship between kinship of form and kinship of genealogy.
Suppose we take a given inorganic substance, for instance a lump of Calcite (CaCO3), and divide it into two pieces, then these pieces will correspond to each other in all intrinsic essential properties. They are qualitatively related, they correspond as to their intrinsic constitution, they show kinship of form. Insofar as a more complex body is involved, such as a granite [which is a kind of volcanic rock], consisting of the three minerals Feldspar, Quartz, and Biotite, one may establish differences between the two pieces. But all in all also these pieces will correspond to each other as to essential properties. Organic beings are distinguished from inorganic beings by their still higher degree of complexity, and therefore still greater differences are to be expected between the pieces resulting from division. Nevertheless in lower organisms does take place a physiologic[ally directed] process of [material] division resulting essentially in qualitatively similar pieces : cell-division in order to reproduce. As is well-known, in higher organisms reproduction takes place by means of especially differentiated cells of the body, possessing the ability to reproduce the whole organism. In line with the higher degree of complexity the individual differences between parents and offspring as well as between offspring of the same parents will be greater as as they are in lower organisms. Nevertheless also here [in higher organinisms] kinship of form does exist between the genealogic related orgamisms, because of the presence of the same essential properties in all of them. Accordingly, one may generally state that if two entities (things) are genealogically related, then they are also qualitatively related. This statement may be called the genealogic main thesis.
When in this way kinship of Becoming determines kinship of Being, one might ask whether also the other way around every kinship of Being is the result of kinship of Becoming. In other words : given two things that are qualitatively related, must they necessarily also be genealogically related? In the realm of inorganic things we definitely can say no. Two pieces of Calcite corresponding to each other in all essential properties are not necessarily genealogically related, i.e. are not necessarily two pieces of one and the same larger piece, but may have originated totally independently in differerent places on the Earth. In the realm of organisms, on the other hand, it has often been assumed that every kinship of form must be the result of kinship of genealogy. But the presence of convergences shows that totally independently from one another and at different times the same organic forms may originate. Therefore the reversal of the genealogic main thesis, which may be called the reverse genealogic main thesis, is also in the realm of organisms not correct. Two living beings that are qualitatively related need not be genealogically related, but may have originated independently. The contrasting view, concluding from kinship of form to kinship of genealogy is false [i.e. although there may be many cases in which one can legitimately conclude from kinship of form to kinship of genealogy, the exceptions disprove the rule. And, moreover, later on it has been discovered that the same holds for the relationship between the qualitative content of a given organism and its genome (its DNA) : Although it seems that the genome (supposed to completely determine genealogic kinship) of a given organism in all cases completely determines the qualitative content of that organism, there are significant and dramatic exceptions (the angler fish and Bonellia, expounded earlier in this website). These exceptions thus demonstrate that there is no intrinsic and necessary conection between the organism's DNA and all of its qualitative content. Genealogic similarity not necessarily entails similarity of form. And so this places a question mark at the thesis that the genotype in all cases completely determines the phenotype (and it is the latter that is the point of application of natural selection!)].
Where did the error come from? It is clear that the more complex a thing is the harder it is for Nature to generate it. It is easier for her to generate iron than an earthworm and this in turn easier than a mammal. The conditions that have to be satisfied in order iron to be generated in or on some celestial body are much more simpler than those that must be present for a living being to be generated on it. But the more complex these conditions are the more rarely they will be realized. This in itself certainly correct thought has led to the assumption that inorganic things may indeed originate independently of each other, but not so organic beings. The generation of them was supposed to be a one-off event. But with all this one forgets about the vast amounts of time Nature has to her disposal making it possible for her to generate even the complex again and again. And above all one ignores the enormous equiformity which is, amidst all differences in detail, present in Nature and which, for example, entails the fact that the ontogeny of living beings takes place billion-fold on Earth again and again along the same path. And convergences clearly show that Nature has the ability also to [evolutionarily] produce organic forms again and again independently of each other. But with this the reverse genealogic main thesis has no validity anymore, and consequently the usual method to conclude to genealogic kinship from existing kinship of form becomes undermined. [ . . . ]
One assumed in all this that a particular organic form could only phylogenetically be produced by Nature a single time, and then, on the basis of this, one expected that all qualitatively related forms must also be genealogically related. An when more and more cases became known in which an allegedly genealogic kinship found in this way did not exist, one felt confused by Nature. But the blame is not to be found in Nature but in the incorrect ideas one has about her, which should be revised in the light of empirical evidence.
Knowing now that kinship of form and kinship of genealogy do not always coincide, also our views concerning the relationship between systematics [i.e. taxonomic classification] and phylogeny must change. The attempts to match systematics and phylogeny with one another, as was visioned by the prevailing evolutionary theory, met with unassailable difficulties already from the standpoint of monophyletic theory. One was forced in systematics to ignore actually existing similarities when they were not the result of genealogic kinship, and inversely ignore existing differences when genealogical unity of a group was present [such as the group consisting of all birds + certain reptiles, a group that is, although containing heterogeneous elements (birds and certain reptiles) is seen, in phylogenetic systematics, as a homogeneous phylogenetic unit because of its supposed monophyly]. In the light of polyphyletic theory the attempt to coalesce systematics and phylogeny loses every legitimacy. Systematics (taxonomic classification) and Phylogeny (classification based on genealogy) both attempt to order living beings in a particular regular way. But the principle that governs order is in both cases a different one. Systmatics in fact is the ordering and arrangement of organisms according to kinship of form, while Phylogenetics orders them according to kinship of genealogy. And because these two principles of ordering things do not go parallel, also the classificatory patterns resulting from them cannot be congruent. Therefore, systematics and phylogenetics should be independent of and separated from each other. This is especially true because the kinship of form figuring in systematics is obtainable in an empirical way, while the establisnment of phylogenetic genealogic kinship has a more or less hypothetical character. When one, for instance, speaks of slugs or of gallinaceous birds, one has to do with morphologically well-characterized true groups. And why should one suppress such empirically found facts, only because these groups are not genetically homogeneous? May difference of Becoming neutralize similarity of Being?
According to monophyletic theory the task of Phylogeny was to establish the ancestral sequences that together finally form a uniform genealogic tree. Initially one assumed that it would be easy to establish such ancestral sequences from fossil material. Practice, however, has shown that many, initially compounded as representing ancestral series are in fact not such series, because between corresponding members of them specialization-crossings occur. Such sequences that generally are based upon the evolutionary transformation of a single organ, in most cases have the character of consecutive stages [Stufenreihen] as Abel calls them, i.e. they show the way in which a given organ has been evolutionarily transformed, without the possessors of it -- the individual members of the sequence -- being connected to each other genealogically [In our noëtic theory this is expressed by the difference between "derivational line" and "morphokline"].
Things may be clarified with the example of the limb- and pelvis-reduction in whales [including teeth- and baleen-whales]. In whales, which are mammals having turned to a life at sea, have evolutionarily experienced a gradual reduction of hind-limbs and pelvis. Fossils show us this reduction following several stages. If one arranges the fossil whales according to increasing reduction of the pelvis, then one obtains a sequence of stages showing the phylogenetic transformation of the pelvis in whales all the way down to complete reduction. But this seqience is not an ancestral one, because if one would order the whales as to the specialization of their teeth, then the resulting sequence would turn out to be quite different, demonstrating the fact that neither sequence could be the true ancestral sequence, because in such a sequence the transformations of all organs should be congruent.
In addition to sequences of stages Abel also distinguishes sequences of adaptation, which show the transformation of a given organ, not within a particular closed group of organisms, but in many groups standing far apart genealogically. [ . . . ]
The concepts of sequence of stages and sequence of adaptation implicitly contain the fact that the transformation of the organs had proceeded polyphyletically, because it belongs to the very essence of these sequences that they contain fragments from the most different genealogical lines, these fragments having been ordered according to the degree of perfection of a given single organ. As Abel notes, many sequences of stages have already been discovered, but only very few genealogical sequences.
In plant phylogeny Zimmermann had shown that it is much more easy to track down the phylogeny of characters than that of phylogenetic taxa, that is, one often can follow the transformation of a single character in phylogeny -- for example the transition of isospory to heterospory and from this to gymnospermia, or the different transformations in the nature of the wood-cylinder --, without it being possible to discover the entangled pathways along which the c o m b i n a t i o n of features has been evolved, characterizing the whole plant. What one in this way obtains are, however, general regularities of phylogenetic transformation, not a particular genealogic tree, as is expected by monophyletic theory [Indeed, by comparing organisms that are, qua qualitative content, not too far apart, we find "derivational lines" of characters (such as wing-venation), which do not represent genealogical series, but give us some indication in what way certain characters might have been evolved and transformed in the course of evolution (which in our theory takes place in the Implicate Order)].
Departing from a completely different viewpoint, Naef has presented a phylogeny of the animal kingdom, completely based on morphological facts, whereby, through morphological comparison of fossil and recent living beings, he has established the morphological form, that may have been possessed by his hypothetical intermediate and ancestral forms. The in this way constructed genealogical pictures are, as expressed by Naef himself, "morphological abstractions". Phylogenetic series found in this way cannot be considered to represent true genealogic series, because in paleontologic research not any of these morphological abstractions have been detected. At best they [i. e. the mentioned phylogenetic series] have their significance in pointing to the ways along which certain phylogenetic transformations might have taken place.
H.F. Osborn and following upon him Sewertzoff state that the evolution of organisms has been compounded in the form of a work of mosaic of the transformations of individual characters. The evolution of the individual characters has proceeded separately.
This conception of the course of phylogeny is also from time to time to be found in the views of other authors, for example in the mentioned view of Zimmermann concering the phylogeny of plants.
Of the mentioned authors only Osborn adheres to polyphyletic theory. We see that also monophyletic phylogeny is forced, by the might of facts, onto such a particular path that completely deviates from that path one initially had expected to follow. One does not establish t h e genealogic tree of organisms, but rather general phylogenetic transformational processes, that often repeat, and proceed in a definite regular way. Dacqué is right when he asserts that all what Phylogeny thus far has been able to establish is adaptational phylogeny.
Within polyphyletic theory it is not hard to understand why one has failed to find t h e genealogic tree of organisms : because it simply doesn't exist. Phylogeny is, precisely as is ontogeny, a regular process, proceeding in billion-fold polyphyly. It can only be the task of the science of Phylogeny to discover the general regularities that history [partially] may reveal, and which govern the transformations of living beings in precisely the same way as they govern the changes of dead matter.
In the next document we will continue with derivational lines and morphoklines in Bibionomorpha.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXXb.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII
Back to Evolutionary Part LXVIII
Back to Evolutionary Part LXIX
Back to Evolutionary Part LXIXa
Back to Evolutionary Part LXIXb
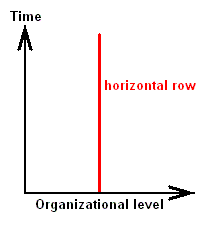
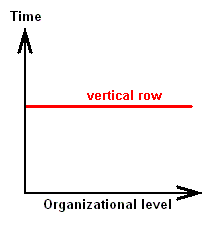
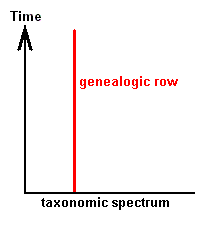
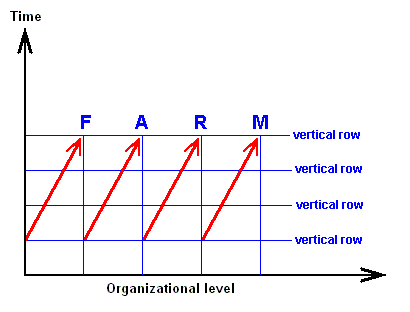
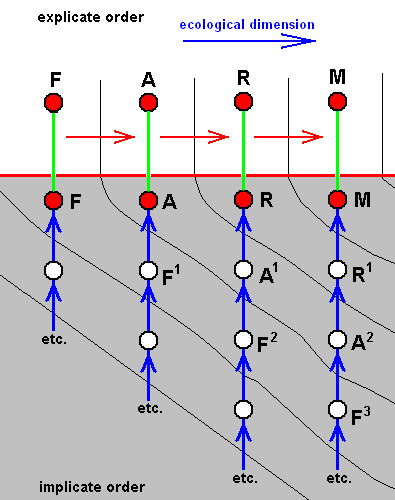
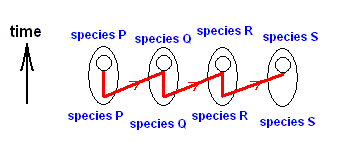
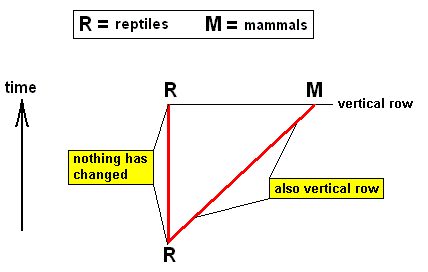
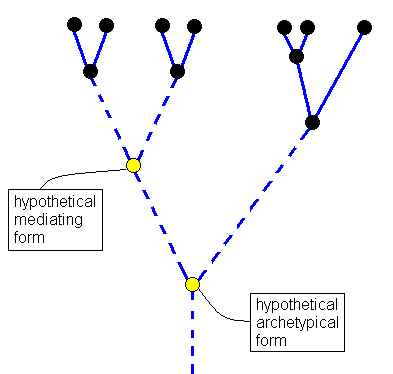
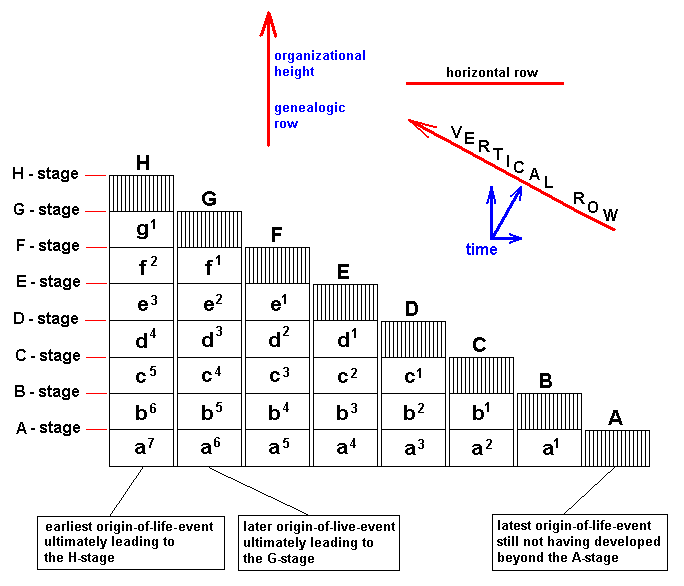
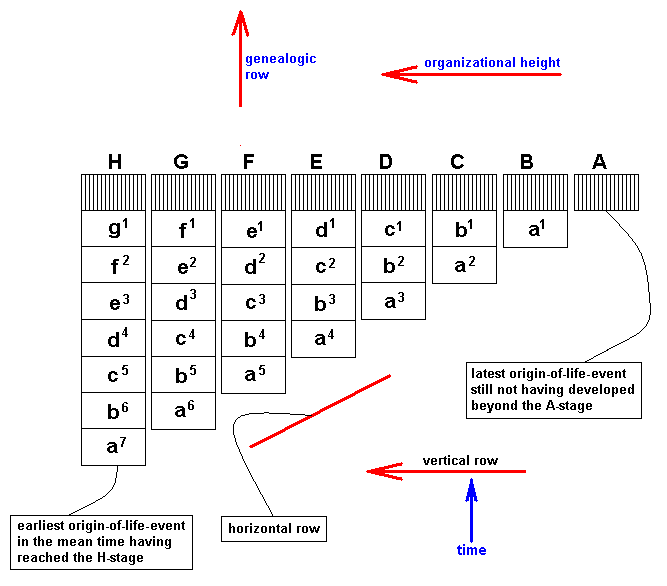
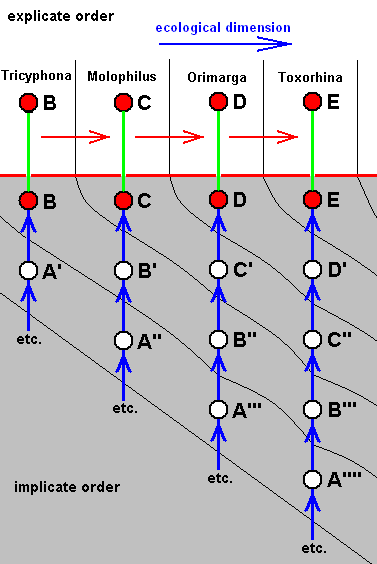{kind=link}
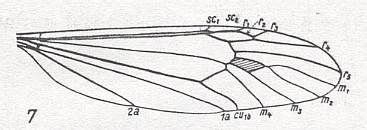{kind=link}