
Introduction
The insect Order Hymenoptera, or 'membranously-winged insects' is represented by the wasps, bees, and ants. Roughly we can say that the order has three Suborders, viz., the Symphyta (saw-flies and allies), the Terebrantia (parasitic wasps, like ichneumon-flies), and the Aculeata (sting-bearers, such as fossorial-wasps, social wasps, bees, and ants). The most primitive Hymenoptera belong to the suborder Symphyta. As larva, most of the Symphyta feed on the foliage of plants in the same way as caterpillars (larvae of butterflies) do. But some cause galls on plants (not to be confused with the genuine gall-wasps [Cynipidae] which belong to one of the next suborders), or live (as larva) in wood (wood-wasps or horn-tails, Siricidae). It is believed that from the Symphyta the Order Terebrantia arose, and from Terebrantia (or their immediate ancestors) the Aculeata.
The next Figures show some representatives of the Hymenoptera (All Figures after SEVERA, F., in ZAHRADNIK, J., Thieme's insektengids).
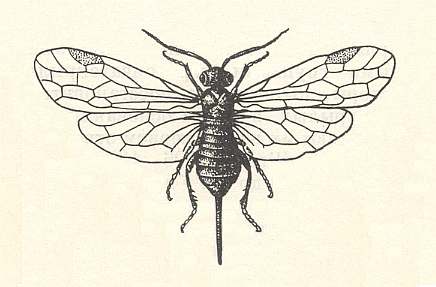
 Figure 1 : Cephaleia abietis, Family Pamphiliidae, Suborder Symphyta. 11-16 mm. |
 Figure 2 : Tenthredo campestris, Family Tenthredinidae, Saw-flies, Suborder Symphyta. 12-14 mm. |
 Figure 3 : Sirex juvencus, Family Siricidae, Wood-wasps, Suborder Symphyta. 14-30 mm. |
 Figure 4 : Hartigia nigra, Family Cephidae, Stem Saw-flies, Suborder Symphyta. 11-15 mm. |
 Figure 5 : Protichneumon pisorius, Family Ichneumonidae, Ichneumon Flies, Suborder Terebrantia. 22-28 mm. |
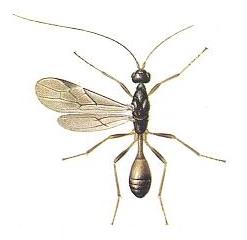 Figure 6 : Spathius exarator, Family Braconidae, Suborder Terebrantia. 4.5 - 5 mm. |
 Figure 7 : Apanteles glomeratus, Family Braconidae, Suborder Terebrantia. 4 mm. |
 Figure 8 : Scolia maculata, Family Scoliidae, Dagger Wasps, Suborder Aculeata. 20-40 mm. |
 Figure 9 : Chrysis ignita, Family Chrysididae, Gold Wasps, Suborder Aculeata. 7 - 10 mm. |
 Figure 10 : Auplopus carbonarius, Family Pompilidae, Spider Killers, Suborder Aculeata. 5.5 - 10 mm. |
 Figure 11 : Sapyga clavicornis, Family Sapygidae, Suborder Aculeata. 8 - 10 mm. |
 Figure 12 : Polistes nimpha, Family Vespidae, Social Pleat-wing Wasps, Suborder Aculeata. 12 - 12.5 mm. |
 Figure 13 : Ammophila sabulosa, Family Sphecidae, Fossorial Wasps, Suborder Aculeata. 16 - 28 mm. |
 Figure 14 : Halictus quadricinctus, Superfamily Apoidea, Bees, Suborder Aculeata. 15 - 16 mm. |
 Figure 15 : Camponotus ligniperda, Family Formicidae, Ants, Suborder Aculeata. 7 - 14 mm. |
 Figure 16 : Bombus pomorum, Superfamily Apoidea, Bees, Suborder Aculeata. The female measures 20 - 24 mm. |
 Figure 17 : Mutilla europaea. Male. Family Mutillidae, Ant-wasps, Suborder Aculeata. 10 - 15 mm. |
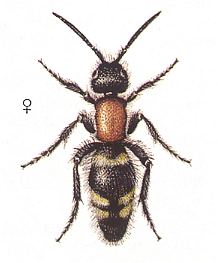 Figure 18 : Mutilla europaea. Female. Family Mutillidae, Ant-wasps, Suborder Aculeata. 10 - 15 mm. |
The Order Hymenoptera, comprising more than 100 000 recent species, distinguishes itself from most other insect orders by, among other things, the fact that they have gone through an evolution that leads from passive strategy (morphological and physiological adaptation, and simple behavior, as is present in most other insects and most animals for that matter) to active strategy. These concepts were explained and discussed at the end of the previous document. Therefore, here we can be succinct. The concept "strategy" encompasses in fact the whole organism, viz., its internal chemistry, physiology, ecology, and behavior (with emphasis on the latter). While the existential conditions of inorganic beings (such as crystals) are merely thermodynamic conditions, the existential condition of an organism is a 'strategy', a strategy to cope with the abiotic and biotic environment, and to secure energy in order to be able to remain in a far-from-thermodynamic-equilibrium state. In all lower organisms this 'strategy' is passive, being merely an adaptation to existing environmental conditions or ecological niches. In certain higher organisms, on the other hand, the strategy is active in virtue of the fact that the living conditions (the ecological niche) are largely created by the organisms -- who possess this strategy, or, who ARE this strategy -- themselves. The culmination of the development of active strategy we see in the social organization of certain species of animals. Among insects we see it in the social Hymenoptera. Remarkably, among insects it also occurs in one of the lower orders, namely in the Order Isoptera (termites). But its highest development, presumably has taken place in the Order Hymenoptera. There, and especially in ants, the height or level of social organization probably surpasses that of any other animal group, including man. In social Hymenoptera the environment is literally created by them. They live in self-made accommodations (nests) and raise their young. They collect and bring in food for the latter, and in some cases even have agriculture. All this is based on deep and sophisticated instincts. Every individual is not merely a member of some aggregation or colony, but is fully integrated in the social community, morphologically, physiologically and ethologically (= behaviorally). The individual is not just participating in the activities of the community merely by reason of expecting from it personal benefit, or of becoming a parasite, or of expecting protection : It is a fully social individual. In fact it is not an individual at all anymore, but an organ or function in, and of, a larger whole, the community.
Preliminary ideas concerning the evolution of Hymenoptera (from lower to higher).
Speaking, not of the origin, but of the evolution of any given group of animals, we usually have in mind the evolutionary formation of so-called 'higher' forms from 'lower' ones. However, the distinction of higher forms from lower ones is not always clear : In many animal groups, such as insect orders, there seems to be no such distinction, or, if there is any, it is only weakly expressed and hard to define. For example, in Diptera we have the Nematocera (midges, gnats, and mosquitoes). These are usually taken to be 'lower diptera', while the Brachycera-cyclorrapha (fruit-flies, parasitic flies, blow-flies, house-flies) are considered to be the 'higher diptera'. But what about the large and flourishing family Cecidomyiidae (= Itonididae) (gall-midges) which nevertheless belong to the Nematocera, i.e. to the 'lower flies'? While it is unclear in this Order, the distinction is clearly expressed in the Hymenoptera. There we see the higher Hymenoptera, such as the Aculeata (sting-bearers), and among them especially the social Hymenoptera, clearly distinguished by their highly advanced and complex instincts from the lower Hymenoptera, the Symphyta, such as the saw-flies, lacking such instincts. So in the Order Hymenoptera evolution has taken place from clearly lower forms to clearly higher forms. And the features that distinguish these forms are mainly those of the various instincts. So in Hymenoptera it is the historical development of complex sophisticated instincts from existing simple instincts that characterizes the evolution of the Order. The simple instincts are part of the passive strategy, while the complex sophisticated instincts represent the active strategy. Like already in all more or less complex adaptations (passive strategies), the evolutionary formation of active strategies cannot be explained by a directed succession of random genetic mutations. So also in this case, if not, even more so, we must assume the Implicate Order to play a role. Here we must assume that the various strategies are, in the form of noëtic patterns, already present in the Implicate Order. They are present there in their proper noëtic stability fields ('fields', that is, not in any spatial sense). And each one of them represents a biological type, which may or may not coincide with, say, a family. As we saw in the Order Diptera, these stability fields are 'successively' visited by one or more noëtic trajectories tracing out evolution, but now, that is, in Hymenoptera, definitely going from lower biological types (simple instincts) to higher biological types (complex instincts). And as in Diptera, projection (from the Implicate [= noëtic] Order to the Explicate [= physical {and biological}] Order) of [originally] noëtic patterns (such as strategies, representing biological types) only takes place when (1) the noëtic trajectory has visited such a pattern, and (2) when appropriate ecological conditions do exist in the Explicate Order. The course of the noëtic trajectory might, as in Diptera, be partly revealed by the methods of phylogenetic systematics (explained in the various previous documents in the Series on Diptera). And, as has been said, in the case of Hymenoptera the trajectory definitely goes from lower strategies to higher strategies (that is, from passive to active strategies). According to all this we can draw a preliminary general picture of the evolution of Hymenoptera :
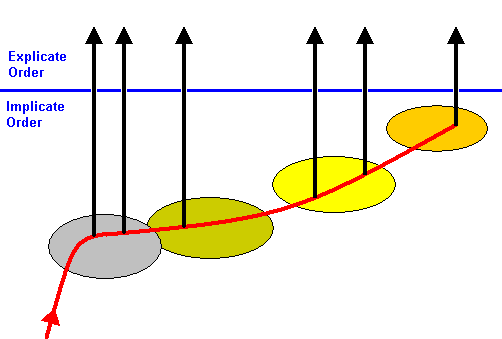
Figure 17 : Sketch of the evolution of Hymenoptera. The noëtic trajectory (red) in the Implicate Order successively visits the stability fields of various noëtic patterns representing strategies (= biological types). Its course is from lower strategies to higher. Upon the noëtic trajectory having visited the stability fields, the corresponding strategies are projected (black vertical arrows) into the Explicate Order, that is, they unfold along space and time dimensions.
What S. I. MALYSHEV -- in Ctanovlenje Perepontsjatokrylich i fasi ich evoljutsii (Origin of the Hymenoptera and the phases of their evolution) (in Russian), 1966 -- calls "phases of evolution", are the phases of the historical development of the strategies (in Hymenoptera).
Origin and early development of Hymenoptera :
Further down we shall follow these phases, as they are realized in groups of Hymenoptera, one by one. Their evolutionary succession in the Explicate Order (where they are observable) expresses the course of the noëtic trajectory in the Implicate Order visiting the noëtic stability fields containing the phases or strategies as noëtic patterns.
To provide the reader already in this early stage of study with an overview of the large insect Order Hymenoptera, we here present a diagram outlining -- as it is proposed by its authors -- the chief evolutionary courses in it, resulting in the evolutionary establishment of the larger hymenopterous groups :
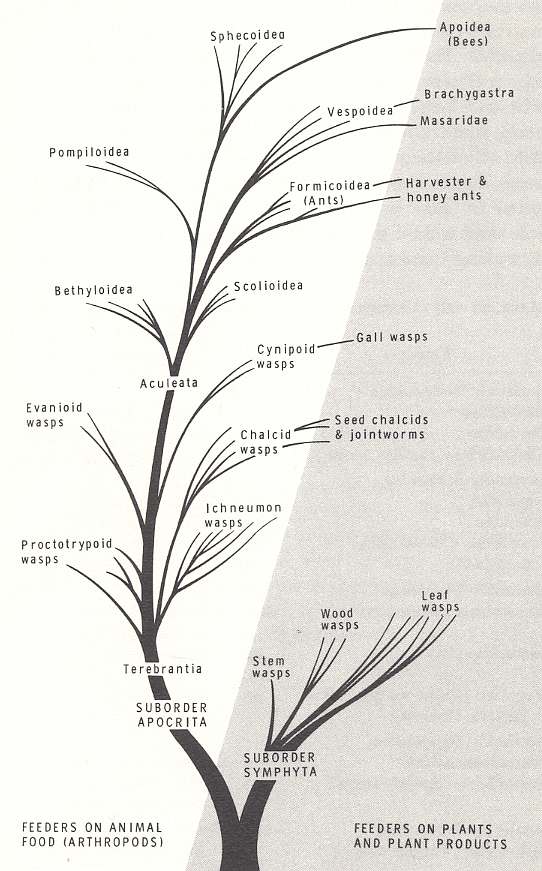
Figure above : Dendrogram showing the probable evolution of the major groups of Hymenoptera. For a magnification click (twice) on the image (and get back by the return button of the browser). (After EVANS and EBERHARD, 1973, The Wasps).
In this dendrogram the Hymenoptera are presented as consisting of two Suborders, the Symphyta and Apocrita. The latter suborder then consists of the Terebrantia and the Aculeata (In our own exposition we consider the Hymenoptera as consisting of three Suborders, the Symphyta, the Terebrantia, and the Aculeata). With respect to the feeding habit of the larvae, the Order originally consisted of phytophags (suborder Symphyta), but soon gave rise to feeders on animal food (arthropods), the terebrants and aculeates. But some representatives of the latter two groups returned to feeding on plants and their products, as is expressed in the diagram.
Strategies as noëtic patterns in the Implicate Order.
As has been said, the just mentioned phases or strategies are already present in the Implicate Order in the form of noëtic patterns. And such a pattern represents in fact a groundplan or organic type. When it is projected into the Explicate Order it exists there in the form of material individuals belonging to one or more biological species. Such an individual thus is the physical-biological expression of a given noëtic pattern. It has a definite morphology, biochemistry, physiology, and behavior, together constituting the expression of the strategy, biological type, or groundplan.
In the Implicate Order the strategy (groundplan, biological type) is completely noëtic, meaning that it exists in an enfolded state, folded back, as it were, along space and time dimensions. It is, as anything in the Implicate Order, immaterial, a kind of mathematical entity or 'thought stuff'. Therefore it was called 'noëtic' (from the Greek 'nous', which means mind).
We assume that in the Implicate Order exist many noëtic elements or basic entities. Further we assume that certain such elements form, by way of aggregation and subsequent integration, patterns, noëtic patterns. The mentioned strategies or groundplans are believed to be such patterns. Aggregation of noëtic elements, only results in a genuine noëtic pattern, that is, will only fully integrate, when the aggregation is free from contradictions, that is, doesn't contain contradictory elements or subaggregations. It must be noëtically consistent. If some alleged noëtic pattern contains a contradiction it must be t w o patterns instead of one. Further, the aggregation must carry some definite identity or nature, in fact it must be such a nature. When noëtically approaching and subsequently traversing such a pattern its identity must appear at some definite point, then continue uninterruptedly, and finally disappear at some other definite point. And not only many noëtic elements, but also many noëtic patterns can (noëtically) react with each other, resulting in noëtic reaction-products. But surely not the members of any random (chosen) pair (or triplet, etc.) of noëtic patterns are able to react with one another (in the same way as Helium does not react with Iron [or any other chemical element for that matter], or as Carbon does not react with water, to think of an analogy). Noëtic reactions, if they can take place, and the formation of the corresponding reaction-products do not take time, because they do not exist in time, neither will the reactants get exhausted partly or completely (as they do in chemical reactions). So we can expect that in the Implicate Order all possible noëtic patterns and all their possible reaction-products (which are also noëtic patterns), that is, all patterns and reaction-products, that can ultimately be derived from the existing stock of noëtic elements (which stock we assume to be unlimited), are (noëtically) present. And this means that also all organic strategies, all groundplans, and all biological types, provided they are noëtically consistent and carry a definite nature or identity, are eternally present in the Implicate Order. In the Explicate Order, on the contrary, only those noëtic patterns are materialized that have been projected from the Implicate Order into the Explicate Order. And, as has been said earlier, this only takes place when the noëtic trajectory has visited the noëtic stability fields of the respective patterns, and when, in addition, the appropriate ecological conditions actually do exist in the Explicate Order.
Preliminary notes on the instincts and their evolution in the Order Hymenoptera.
Although the more or less gradual evolutionary transformation of instincts -- from initially passive strategies to active strategies, and within the latter to high-level active (and teleological) strategies -- in (female) Hymenoptera will systematically and extensively be expounded later on, it is perhaps instructive already here to present a general and more or les abstract example of a definite part of the transformational chain, in order to let the reader know what we're taking about, and especially what precisely 'higher development', that is, development from lower to higher forms actually means. Let us, following MALYSHEV, 1966, pp.146, generally describe the successive transformations of the instincts of wasps (vespoids) by signifying the individual acts by symbols. For doing so, let us indicate where, in the chain of transformations, we here begin.
The forms that stand evolutionarily before the genuine wasps (vespoids, here solitarily living, such as fossorial wasps) are the Terebrantia, that is, the so-called parasitic wasps (ichneumon-flies and allies). In order to guarantee survival and development of their offspring many of them lay an egg in some insect larva. After that they abandon the latter, leave it where they had found it. Now, things being so, there is always a chance that the mentioned insect larva gets parasitized once again, perhaps by some other species of parasitic wasp. The orginal egg or larva might then be destroyed by the newcomer, or it might itself become parasitized by the latter (hyperparasitism). Anyway, successful development of offspring is in the parasitic wasps (Terebrantia, ichneumonoids) not fully guaranteed. And indeed, in the next phase of the development of female instincts, present in the evolutionarily next wasps, the (lower) vespoids, we see that the female wasp looks for, or prepares, some hide-away place for its infected prey (host of the wasp's larva). And in order to prevent any escape of the prey the wasp paralyzes it by one or more precisely directed stings that administer a special poison. Here we thus see the origin of the first Aculeata from some (primitive) Terebrantia. And within these first Aculeata the new instinct will develop further, which we will now describe -- but here still in a more general and abstract way by providing the various individual elements of the original activities of the female wasp of the type Pompilus viaticus ( Pompilidae, Aculeata) with different symbols :
First vespoid (pompiloid) phase (= present phase of development)
A = Hunt :
a1 = Finding a prey.
a2 = Attacking the prey and paralyzing it.
a3 = Bring it to a certain place for it to stay temporarily.
B = Constructing or adapting a nest (= definite quarters of the prey) :
b1 = Finding an appropriate place for the nest.
b2 = Burrowing of the nest, adapting it, etc., usually in connection with visiting the prey.
C = Transporting the prey to the nest.
c1 = Placing the prey before the nest.
c2 = Inspecting the nest.
c3 = Bringing the prey into the nest.
D = Laying of the egg.
E = Closure of the nest (from the outside).
e1 = Closure of the true cell.
e2 = Closure of the nest itself.
Each one of these acts itself, of course, represents a complex action, which might be subdivided into a number of more simple ones. But usually such subdivision demands a substantial knowledge of the way of life, not only of the species concerned, but also of the forms that are related to it. For our purposes it is, for the time being, sufficient to acknowledge that the primitive maternal activety of the wasp consists of the above enumerated acts. With the help of the given symbols (indexes) we now characterize the work of the wasp that executes the original (i.e. as a starting point) order of actions as we see it in the present phase (the first vespoid [pompiloid] phase), for instance in Pompilus viaticus Latr.
Which can more compactly be written as :
Because normally the wasp takes care for several young, the complete maternal activity is of course polyserial :
or
So this is the original order of the basic instinctive actions of the maternal wasp in her providing for her offspring.
In this stage, in which we are now, following the development of the maternal instinct of the Hymenoptera, arises a great obstacle in the further evolution of the hunting as well as the building instincts. This obstacle is implied by the order of the activities at which we have now arrived. It consists in the fact that first of all the provisions (food for the larva) are taken care of, and only after that the place (nest) is being found or built to store it in. Such an order of work unavoidably brings with it a number of difficulties. First of all in such conditions the catch must have, from the very beginning, a sufficient large size in order for it to be enough food for the larva. From this it follows that a great effort of the wasp is needed, and with it a loss of time ensues for it, because it can hardly use its wings for this job. And then, for the transport as well as for the feeding of the larva the catch (the prey) must be well-paralyzed. And for this it is necesary that the catch has an appropriate organization of its nervous system and a definite relation to the poison of the paralyzer. All this strongly limits the selection of the catch. Finally, and this is perhaps the most important obstacle, the discovered, catched, and paralyzed prey must necessary be left alone without supervision until the wasp has found an appropriate location for the nest and until it has built one. In such circumstances, as is confirmed by direct observations, the catch, even when it had been brought right away to the building site of the nest, not seldom becomes the property of someone else, and the efforts of the builder become in vain. There are even (other) wasps that specialize in preying upon paralyzed catches. It is unnecessary to say that under these circumstances the building of a more sophisticated nest is impossible for all the wasps of the present phase.
And indeed, a new order of activities (that is, a new arrangement of the elementary acts) -- first building the nest, and only after this (has been done) hunting a prey -- is a great step forward on the road of development of maternal instincts in certain wasps. And so we arrive at the next phase of development of instinct :
Second vespoid (sphecoid) phase
Indeed, the well-known sphecid wasp Ammophila sabulosa L. (see Figure 13 above ) first burrows its characteristic nest, widened at its end, and only thereafter (usually the other day) brings in it a paralyzed caterpillar.
In what way did this more complex order of actions arise?
We cannot assume that in the present case the wasp simply thinks about putting the second act (B - building the nest) at the place of the first (or the first at the place of the second). We can only assume that in the ladder of the consecutive acts, geared to the provision of the offspring, the first one (A - hunting and all what is connected with it) was cancelled, while the second -- building the nest -- remained. Cancelling of individual actions from their ladder (series of consecutive actions) have undoubtedly played an important role in the historical development of maternal instincts of wasps and bees. However, we should not think that the case is simply this, or went in such a simple way.
When we assume that at the establishment of the new order (for instance in the mentioned Ammophila sabulosa) the first act was cancelled, this just means that the work of the wasp simply begins directly with the second, while the act that was originally the first act takes place later. Now the series will look like this (to recall the meaning of the symbols click HERE ) :
As has been said, the wasp will generally provide for more than one descendant, meaning that several such series, that is originally : A + B + C + D + E, A1 + B1 + C1 + D1 + E1, A2 + B2 + C2 + D2 + E2, A3 + B3 + C3 + D3 + E3, etc. are executed. Said differently, the wasp will end up with a number nests each one of which provided with a paralyzed prey for a larva to feed on.
Now back to our new series B + A + C + D + E.
As regards B (construction of the nest) there is now no doubt that it is a remains of the first series (A + B + C + D + E). But from where was A (hunt) taken? If the first act of the first series were indeed cancelled, then the insect could essentially not finish those acts connected with A, namely C (bringing the catch into the nest) and D (laying of the egg), when we take into account the presence of the instinct to lay-the-egg-on-the-prey. In such circumstances the whole original series would read (where "0" = zero) :
It is not hard to understand that the last act, E (closure of the nest), now becoming useless, also might be cancelled, and that the wasp now continues with the first act (A1) of the second series (A1 + B1 + C1 + D1 + E1), but now cancelling act B1 (constructing the second nest), which is now not necessary because a nest is already constructed (B). In this we get the following combination :
that is, precisely what we were looking for. From this it follows that the catched prey (A1) originally belonging to the second nest (built in the second series) is now placed by the wasp in the first nest. So in this way we have, not an exchange of the first acts (A and B), not a displacement in time of those acts, but just the combination of them out of two successive series ( B from the first series, while A1, C1, D1, and E1 from the second series) which is indeed expressed by our symbols. Therefore A1 of the new series is not essentially that, what A is of the first series.
That things went precisely that way, that is, by combining two successive series, is convincingly demonstrated by the Ammophila's. Thus, A. sabulosa L. and A. argentata Lep., after having first burrowed a nest, and having closed it while it is still empty, hunt down a prey, and then place it in the nest, lay their egg and finish their work as usual.
The above established series of acts can be further explained : Here we in fact have from the originally first series : 0 + B + 0 + 0 + E, and in addition to it from the second series : A1 + B1 + C1 + D1 + E1, where B1 -- the building act of the second series -- is here the removal of the temporary cap (the temporary closure of the nest was act E ). If we add up these two acts (E and B1) we get zero, and thus we indeed end up with our new series B + A1 + C1 + D1 + E1. And because here the hunt (for the prey) A1 and bringing it into the nest (act C1) are closely connected and not separated anymore by the construction of the nest, we can express this as (to recall the meaning of the symbols click HERE ) :
In connection with this it is not superfluous to mention the fact that Psammophila hirsuta Kby., which still works according to the original order, that is, first catching and making ready the prey and only then building a nest, does not make a temporary cap of the nest.
It is interesting that temporary closure of the nest (cell) is sometimes also observed in other wasps providing the cell with just one prey. This is seen in, for example, Sphex albisectus Lep. and Tachytes panzeri v.d. Lind., that make ready grashoppers (of the family Pamphagidae).
Anyway, it is perfectly clear that the cancellation of the first act (the hunt) could not be indifferent to the species and that it directly will trigger a corresponding change in the whole course of the work (of the wasp), a change comprising first of all (as far as we can detect all this) the next series. In the present phase of development (second vespoid [sphecoid] phase) of the plasticity of the instincts of wasps a hard testing is shown, but the ability for self-regulation made it possible for the instinct to overcome the difficulty and have led it along a broad route : All higher wasps and all independently nest-building bees, without exception, first construct a cell, and only then take care of catching and working up a prey, that is, only then provide it with provisions.
This new work order as it appears in the present phase (second vespoid [sphecoid] phase) of development, when into the initially built cell only one prey is placed, is at present observed only in a few, partly already mentioned, fossorial wasps (Sphecoidea), and as an exception also in Pompilidae. In the rest it either almost does not at all appear or its [i.e. the new work order] original phase disappears without leaving a trace.
We will see why precisely wasps in their majority did not halt at the present phase of development but went further. We see that already in this phase a new work order has favorably expressed itself in the development of nest-building instincts in wasps. First of all, it is now for them possible to concentrate nest-building in definite places from which came to be born the habit of settling down in colonies from year to year in a favorably chosen location. The methods themselves applied in their construction work became more subtle and complex. As a result individual parts of the nest obtained a special shape, and in times also a larger size, where in one nest it became possible to built not one but several cells.
Third vespoid (crabroid) phase
The new and relatively more perfect work order of the wasps did nevertheless contain an essential insufficiency : To the place where (a little) earlier a nest had been constructed a prey has to be brought, taken in the undefinite circumstances of the hunt, and with it, having a rather large size. From this difficulty a way out was found. This took place thanks to the fact that the prepared nest, before provisions were obtained, itself formed the basis for further, and, as we can see with hindsight, fundamental change of the instinct of provision (supplying food to store in the nest). It is namely so that now there exists the possibility to bring provisions into the already built nest not in a single final portion, as it was until now, but in several such portions, i.e. to dose provisions. To begin with, in the cell was placed, instead of one large prey, two, but smaller ones. The effort of transporting the prey became lighter, especially when a prey was not directly at hand and therefore taken at some considerable distance from the nest. With all this the work order changed a bit. Expressed with our symbols (to recall the meaning of the symbols click HERE ) :
[builts a nest, catches a (small) prey, brings it into the nest, lays an egg on it, catches a second (small) prey, brings it into the nest, closes the nest (from outside)]
In order to understand this consecutive order of acts let us try to base it on the same principle with which we understood the previous series. Assuming accordingly that also in the present case certain individual acts of the serial chain were cancelled and that the remaining acts were unified without temporal conversion, we get :
First series (B + A1 + + C1 + D1 + 0) + second series (0 + A2 + C2 + 0 + E2) =
B + A1 + C1 + D1 + A2 + C2 + E2
So we have now the series given just above. But considering all individual acts that indeed are observed in nature, there are in fact more acts involved than it seems from our new formula. But we can show that nevertheless this formula is correct : Ammophila urnaria Gress. places, according to Peckham, 1898, generally two caterpillars (sometimes only one), where it temporarily closes the nest -- first after finishing the nest, and then after the egg has been laid on the first prey (on the second no egg is laid). Accordingly the whole work done to provide one larva can be represented as follows :
The opening of the nest (removal of the cap) -- second act of second series -- can be considered to be 'digging' and thus be qualitatively equated with B (burrowing of the nest), and can consequently be labelled as B1. The same goes for B2. So in words we can express the above formula as :
Burrowing a nest (B), Temporarily closing the nest (E), Catching the first prey (A1), Opening the nest (B1), bringing in the prey (C1), Laying an egg on it (D1), temporarily closing the nest (E1), Catching a second prey (A2), opening the nest (B2), bringing in the prey (C2), definitively closing the nest (E2).
The temporary closure (E and E1) of the nest with a cap, and the removal of this temporary cap (B1 and B2) cancel each other, so we can take out E, E1, B1, and B2, from the above formula, resulting in :
So we have now indeed the above series.
Apparently the present way of complexification of the instincts (i.e. by way of cancelling certain acts followed by combination of the remaining acts) went further, following the increase of the number of preys being stored in a nest and their corresponding decrease of size.
Temporary closure of the nest when providing it with provisions is sometimes also done by other fossorial wasps. Anyway, the temporary closure and opening of the nest might, under certain circumstances, become superfluous, and finally disappear. So, A. holosericea L., who also brings in several (up to 5) preys, already does not temporarily close the nest. The cancellation of these superfluous acts, alleviating the work of the wasp, helps in increasing further the number of preys in the nest in accordance with a corresponding decrease of their size. And in addition, the uniform repetition of the (whole) act might create a certain inertia, which then is effective in the same direction (that is, a rhytmic sequence of acts tends to continue). It is this repetition without [causing an] essential change [of the meaning] of the act just finished before it, that undoubtedly also was of great significance in the formation of maternal instincts in wasps and bees.
Taking into account the indissoluble consecutive order in such a provioning of two acts -- hunt (A) and the bringing in of the prey (C) -- the previous series might be expressed as follows (again, to recall the meaning of the symbols click HERE ) :
where m = n+1.
And when we do not indicate anymore from what series the individual acts came from we get :
where (AC)n signifies the acts of successively hunting and bringing in n preys, after the egg has already been laid (D).
In solitary wasps, at an increased pace placing several preys in one nest, is sometimes observed yet another complexification. It consists in the fact that the egg is not laid on the first prey that has been brought in, but on some of the next, or even on the last, which we can write as :
In this case, with rapid provision, not interrupted in these wasps by some other action (temporal closure and opening of the nest, periods of awaiting), the act of laying the egg looses the connection with the first prey, and takes now place only at the completion of provisioning, so that the closure of the nest immediately follows upon the the laying of the egg. This is observed, for instance, in the American species of Sceliphron and Podium.
With this we conclude our preliminary example of the evolutionary transformation of instinct in Hymenoptera. It should serve the purpose of telling the reader what we're talking about when considering the (evolution of) strategies in Hymenoptera. In due course we will expound the complete series of evolutionary transformations of the maternal instincts (as far as known) in the whole of Hymenoptera. And as has been said, these transformations will culminate in the social Hymenoptera, especially the ants. Further we will try to establish roughly the genealogic (phylogenetic) relationships between the higher taxa of Hymenoptera. The resulting cladogram (phylogenetic tree) expresses, according to our general theory, the course of the noëtic trajectory (trajectories) in the Implicate Order. It will visit the stability fields of the noëtic patterns representing the different strategies.
The phases of development of strategies in Hymenoptera
As we have said, a strategy is the way for an organism to exist and persist. This way may consist simply in its possessing a special physiology and morphology, and a corresponding more or less simple behavior, making it possible for the organism to absorb, store, and spend energy needed to maintain its far-from-thermodynamic-eqiulibrium state (which itself makes a special, complex and stable, but still malleable, morphology of any macroscopic material entity possible at all). This is, what we call, a passive strategy. The emphasis is on material construction (morphology, physiology, and simple behavior). On the other hand, things are different in active strategies. Here, in addition to, and as further completion of, the morphology and physiology, we have the (higher) instincts that direct the behavior of the organism along lines leading, but often only indirectly so, to a specific goal, which in the very end is, like in all the simpler cases, the existence and persistence of the organism, in which the mere existence refers to that of the individual, while persistence refers to the existence of the species (the supra-individual or type).
Especially in the insect Order Hymenoptera we see (in fact deduce) an evolution from mere passive strategy to active strategy. In the latter strategy a part of the environment guaranteeing the individual development of, say, a wasp, is not simply found, but created all by itself, that is by its mother, led by special maternal instincts. The acts ensuing from such instincts are strongly teleological, that is, triggered by causes that do not yet exist. This leads us outside the domain of physics and simple biology. To this we will return later.
MALYSHEV, 1966, after several decades of studying the Hymenoptera and their behavior, has distinguished several more or less successive, evolutionary but still existing, phases of development of strategies : From the more primitive phases in the lower Hymenoptera (Symphyta [saw-flies and allies] and (partly) Terebrantia [parasitic wasps] ) to the utterly sophisticated ones in the higher Hymenoptera (mainly the Aculeata, like the [higher] wasps, all the bees and all the ants). A list of these phases was given ABOVE . To be able at all to (more or less) reconstruct the evolution of strategies in Hymenoptera, we are greatly helped by the fact that a number of representatives of earlier phases (of historical development of these strategies) are still living today. They have become relict groups usually with only a small number of species. Thanks to the acquisition of often special adaptations they have managed to persist. So it is not so that the evolutionary process simply consisted in a further development of the strategies, that is, perfecting them more and more. If that would be the case we would encounter today only the most advanced strategies. But things are different. There still exist enough remnants of primitive ancient strategies for making it possible for us to reconstruct the evolution of complex and advanced strategies (from such primitive strategies).
We will now -- following MALYSHEV, 1966 -- successively describe these phases (ancient and modern) provided with examples of representatives of them (still) living today. We will concentrate on the possibility of deriving one phase from another, allowing us to place them in an evolutionary context. [ The book of MALYSHEV, just referred to, on the evolution of instincts in Hymenoptera, Stanovljenje perepontsjatokrylich i fasi ich evoljutsii (Origin of the Hymenoptera and the phases of their evolution), written in Russian, is, as far as I know not translated into some western language. Therefore I suppose that its theories are virtually unknown in the West. Although I have no intention to literally translate the whole book, I will translate and/or paraphrase much of its content, add explanations where necessary, and supplement it with data from other sources, and also with my own views.]
It is assumed that the Hymenoptera have evolved from Mecopteriod ancestors. As we know, the insect Order Mecoptera (Scorpion-flies and allies) was in Permian times (that is the period before the advent of the Dynasaurs) much more abundant and diverse than it is today. It has given rise not only to the Diptera (see Part XIV ), but also to the Trichoptera (caddis-flies) and Lepidoptera (butterflies and moths), but also, as has been said, to the Hymenoptera. A picture of some recent representatives of the Order Mecoptera is reproduced here :

Figure :18 Recent representatives of the Order Mecoptera.
1 - Panorpa communis, Family Panorpidae. Length 20 mm, wing-span 25-30 mm.
2 - Bittacus italicus, Family Bittacidae. Wing-span 35-40 mm.
(Afer ZAHRADNIK and SEVERA, 1977, Thieme's insektengids)
It is important to know that the mecopteroid ancestors of the Hymenoptera did not possess specialized instincts. They laid their eggs in more or less indefinite or accidental places and did not show any care for them or for the young. Having emerged from the egg the larvae live a wandering life and independently provide themselves with food, in the same way as do the larvae of recent scorpion-flies, of Rhaphidioptera, or larvae of beetles such as Lampyrids and Carabids, etc. Because the Order Mecoptera is one of the most primitive orders of Holometabola (insects with complete metamorphosis) -- and might throw light on the first Hymenoptera -- it is interesting to consider them more closely (but again, see also Part XIV ).
One might suppose that the larva of Merope tuber Neuman. (Mecoptera), which is still (1966) unknown, has a similar way of life, while the larvae of the "living fossil", the Chilean Notiothauma reedi McLEDEN (Notiothaumidae), as is supposed, live in the water like the larvae of caddis-flies do.
From these data follows that saprophagy was the primary type of feeding of the primitive caterpillar-like (erucoid) larvae of Mecoptera. This is wholly in line with the general thesis according to which "in all branches of the animal world saprophagy and zoophagy represent the more primitive types of feeding. Phytophagy, on the other hand, has been worked out only in the later stages of evolution.". In correspondence with this we find among the larvae of lower Holometabola (insects with complete metamorphosis) chiefly predators and saprophags. Also with respect to the larvae of Mecoptera we may exclude phytophagy. Feeding by some of them, together with disintegrating plant remains, also living parts of moss, lichens, and fungi, is, probably, in a certain degree supplementary and not intrinsically necessary.
So then, accordingly, they were, the representatives of the archaic soil-dwelling phase, the phase in which the nearest ancestors of the Hymenoptera were in.
Having obtained the results, which are so important to us, results that is, about the life of the most recent ancestors of the Hymenoptera, we can now go to the next stage of development.
In the next document we will, accordingly, consider the Ectophytic saw-fly phase of the evolutionary development in Hymenoptera.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XXXII.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX