
After having dealt with a number of functional wing-types as they occur in the Suborder Nematocera, we will, before continuing this survey, investigate the status of wing-venation from a biological (i.e. adaptive) as well as from a formal (i.e. derivational) viewpoint.
Introduction
Our website is chiefly meant to investigate the metaphysical structure of Reality, that is, its ontological structure or the elements making up true Being (in sofar Being). In all this, the Aristotelian concept of Substance (independent complete individual being) is central (further developed in First Part of Website (LINK Home page). Later we have worked out the division of Reality into two principal domains of being, the Explicate and Implicate Orders (Third, Fourth, and especially in Fifth Part of Website). The concept of Substance was still further worked out for inorganic beings such as crystals (in First, Second, and Fourth Part of website), and after having further elaborated on the concepts of the Explicate and Implicate Orders as different ontological domains for Substances to exist, we felt it necessary to introduce organisms -- substances par excellence -- into our new metaphysical theory. And we realized that if we wanted to test, improve, and further develop, this metaphysical theory, then we had to deal extensively with the phenomenon of organic evolution (not normally done by philosophers!), because it is here that the most baffling events have taken place, such events, that is, that might compromize our metaphysical theory (and if it does, so be it). For this I have chosen to study and incorporate into the present website the evolution of insects because, (1) they form a large (if not, huge), extremely diverse, animal group existing already for a long time (since the Carboniferous, more than 300 million years ago), and (2) I am well acquainted with them.
In the course of our website (in the latter two thirds of Fifth Part of Website) dealing with insects (especially of the Orders Diptera and Hymenoptera) much attention was given to the wing-venation. This had two reasons :
1. The venational pattern in the wings of insects is relatively easy to study, because it is largely a two-dimensional macroscopic structure that is fully exposed and therefore easy to describe.
2. The wing-venation is virtually the only feature that is well preserved in fossil insects (of the Subclass Pterygota -- originally winged insects).
We studied the venation in Diptera and in Hymenoptera, and we can now say that the nature of the venation, although complying to the same underlying fundamental scheme of longitudinal veins, is very different in the one insect Order as compared to the other. Especially in the Order Diptera we saw (and will see) that the venation is very diverse indeed, giving rise to many extravagant versions of the original prototypic scheme. And every insect Order has its own type of wing-venation and its evolution. In short, in insect evolution the wing-venation is 'on the move'. This has prompted me to ask for the significance (biological and formal) of the wing-venation, its precise role in the life and evolution of insects, and its status in the Implicate Order. Is it completely adaptive, functional, and biological, or is it, at least partly, a purely formal feature of insects (Pterygota)? Or, more generally, we ask : Why is the wing-venation so diverse and 'worked out' in insects, and what can it tell us about evolution?
Determinations of kinships of species and higher taxa in evolutionary systematics of organisms is centralized around the concept of homology of corresponding structures, for example the wings of birds and the front-legs of other vertebrates. Two given structures are homologous if they allow us to assume that they are historically connected with each other. So here -- in evolutionary systematics (taxonomy) -- it is assumed that these structures have, in the course of time, materially directly or indirectly originated, one from the other. In the context of conventional evolutionary theory this can only take place through corresponding changes of the genome (set of genes) of representatives of a given organic species. When the particular corresponding structures of the one and the other species [or higher taxon] lack demonstrable historical continuity (for instance wings of insects and those of birds) they are at most functionally similar, and not homologous. They are only analogous.
Concerning the originally winged insects, that is, insects having wings or having lost them later on in evolution (Pterygota), it is assumed that all pterygote species have materially originated one from the other, and ultimately from the one ancestral form of Pterygota. All morphological (and other) sub-structures of the Pterygota have then originated from that ancestral form and thus also their wing-venation. So the venational patterns in the wings of all originally winged insects have ultimately originated from one ancestral venation (empirical data suggest us to assume this). This ancestral, prototypic wing-venation is found by Comstock and Needham through comparative investigation of many existing wing-venations of insects, especially those of the representatives of primitive Orders (and the primitive members of them). Especially the ontogenetic course (individual development) of the wing-venation in such primitive Pterygota (such as stone-flies, Plecoptera) allowed them to set up and propose their ancestral venation. This was their result :
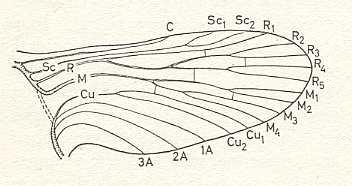
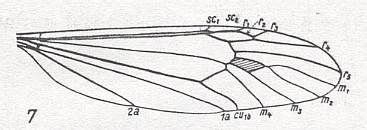
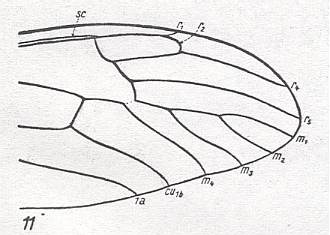
In this way it is more or less possible to determine in a given wing-venation the identity of the individual (longitudinal) veins, i.e. to find their homologous counterparts in all winged insects. These longitudinal veins are in the overall prototype the following : Costa(C), Subcosta (SC), Radius (R, or if one wants, R1), four branches of the Radial Sector (R2, R3, R4, R5), four branches of the Media (M1, M2, M3, M4), two branches of the Cubitus (Cu1, Cu2, or, if one wants, CuA, CuP), and three anal veins (1A, 2A, 3A, or, if one wants, A1, A2, A3, or 1a, 2a, 3a). By later researchers it was assumed that in the most original wing-venation of Carboniferous insects the Media possessed yet an extra anterior branch (MA). Also the position of veins in the wing-membrane -- running on a ridge ('convex veins'), running in a groove ('concave veins') -- was considered by some authors to be important for the recognition of their identity.
Empirical data let us further assume that every insect Order has its own venational prototype (which, however, in all cases is derivable from the all-insect prototype [the mentioned comstock-needham scheme of longitudinal veins] ).
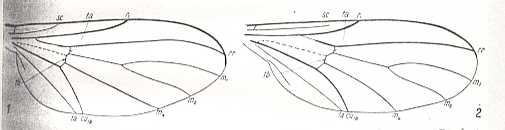
The venational prototype of the Order Diptera is still very close to the all-insect original venation. See next Figure.
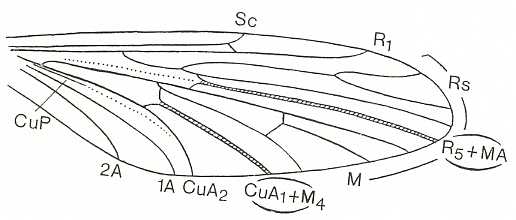
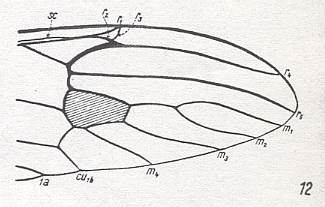
The most important deviations from it consist of the supposed coalescence of R5 and MA on the one hand, and that of M4 and CuA1 (the anterior branch of the anterior Cubitus) on the other (if we assume the veins CuA1 and MA to be present in the ancestral all-insect prototype). R5+MA is then called "R5", and M4+CuA1 is called "M4".
CuA2 (the posterior branch of the anterior Cubitus) runs freely into the wing-margin and may be called just "CuA". CuP (the posterior Cubitus) is merely a rudiment or fold, and does not reach the wing-margin. It is often not drawn in figures.
So when we now go from front to back around the wing-margin, we'll meet -- in this order-prototype, the diptera-prototype -- successively the following veins ending up at the wing-margin : SC, R1, R2, R3, R4, R5, M1, M2, M3, M4, CuA, 1A, and 2A.
If one now can -- again, for example, in Diptera -- retrieve the (evolutionary) transformations that have taken place in wing-venations in the course of geologic history, then one has data saying something not only about the transformational history of those wing-venations, but also about that of the species (and higher taxa) carrying these venations, that is, one then has extra data about the phylogeny of Diptera. Investigators such as HENNIG (1954, 1969) have worked along these lines.
In our theory of evolution (presented on the present website) it is asked whether or not organisms have indeed actually materially and historically originated from each other. There are many indications for it not to be so, for if this ["materially originated from each other"] were indeed the case, it had to be mediated by genes and their transformations, and such an assumption runs into difficulties :
First, the many existing complex, subtle, and 'sophisticated' adaptations, in the form of 'techical constructions' of and in the body, or in the form of subtle behavioral patterns, are such that they cannot originate and stabilize as a result of a succession of random genetic mutations followed by natural selection (as conventional evolutionary theorists believe). We have already discussed this earlier. In wing-venations the assumed transformations are, it is true, less complex, but nevertheless subtle enough to speak against such a course of things.
Secondly, specialization-crossings are wide-spread in the animal kingdom (and probably in the plant kingdom as well). And they also occur frequently in the venational patterns of the wings of insects (we saw this for instance in Part XXIX of the present Series, in Limoniidae ). The limiting effect of a specialization-crossing is based on the following :
If we accept the possibility of material transformations in organisms (through or not through the genome), then the essence of all such evolutionary transformations must be based on laws, transformation laws. And indeed one has, in paleontology and evolutionary biology, set up such laws (that are supposed to govern these transformation processes). The most important one of them is Dollo's Law of Irreversibility : Evolution cannot reverse its course, also not with respect to mere details. That is to say a given structure or relationship cannot evolutionarily reverse to precisely that condition (that original structure) from which it had been evolutionarily evolved. And now we often see the following state of affairs :
An organism B (or a wing-venation B for that matter) may have originated from an organism A (or from wing-venation A) because B possesses a feature F(1) which allows it to be seen as a product of transformation of a homologous feature F(0) in A. But A possesses yet another feature, F(4), which allows it to be seen as a product of transformation of the homologous feature F(3) in B. If we now assume that B has materially evolved from A, then we have :
F(4) ==> F(3)
in which, consequently, F(4) ==> F(3) is evolution running back, and thus a state of affairs violating Dollo's Law. And if we then -- to remedy this -- suppose that A has been materially evolved from B, then we have :
F(3) ==> F(4)
And although now the transformation F(3) ==> F(4) is legitimate, the transformation F(1) ==> F(0) is not, because that is reverse evolution. So A and B cannot have evolved from one another (A not from B, B not from A), made evident by the described specialization-crossing. One then is forced to project the common ancestral species (or structure), from which A as well as B have been evolved, farther back in time (farther back into the phylogenetic history of A and B), and all too often one has indeed done so in phylogenetic systematics, resulting in the bifurcation points lying almost at the beginning of evolution. And because specialization-crossings are abundant in organismic species (and also in wing-venations) which are compared to each other, this might be an indication that these species (and also wing-venations) have not at all been materially evolved from each other. Already in 1938 Ignaz Lichtig in his "Die Entstehung des Lebens durch stetige Schöpfung" was drawn by this (and other phenomena) to the conclusion of a polyphyletic (instead of monophyletic) development of organisms.
As regards wing-venations this might support the assumption that any given venation has not been evolved from some other venation, but only and directly from the venational order-prototype, which itself has evolved directly from the all-insect prototype (the comstock-needham scheme of longitudinal veins), because then we have not to do with specialization-crossings anymore. But also this remedy -- now concentrating on the evolution of venational patterns in the wings of insects -- runs into trouble, already discussed earlier, and here summarized systematically in the form of twelve points :
All this, that is points 1-5, not only does apply to organic species/strategies, but also to particular organ-systems or parts of them, such as the wing-venation in insects. Also in wing-venation we have to do with subtle constructions (structures) (some of them directing air flows) and with specialization-crossings. And also here we assume that there actually has taken place (in the Implicate Order) a formal derivation which we -- the investigator -- try to unravel by attempting to derive wing-venations (or their elements) (linearly) from each other or from the prototype.
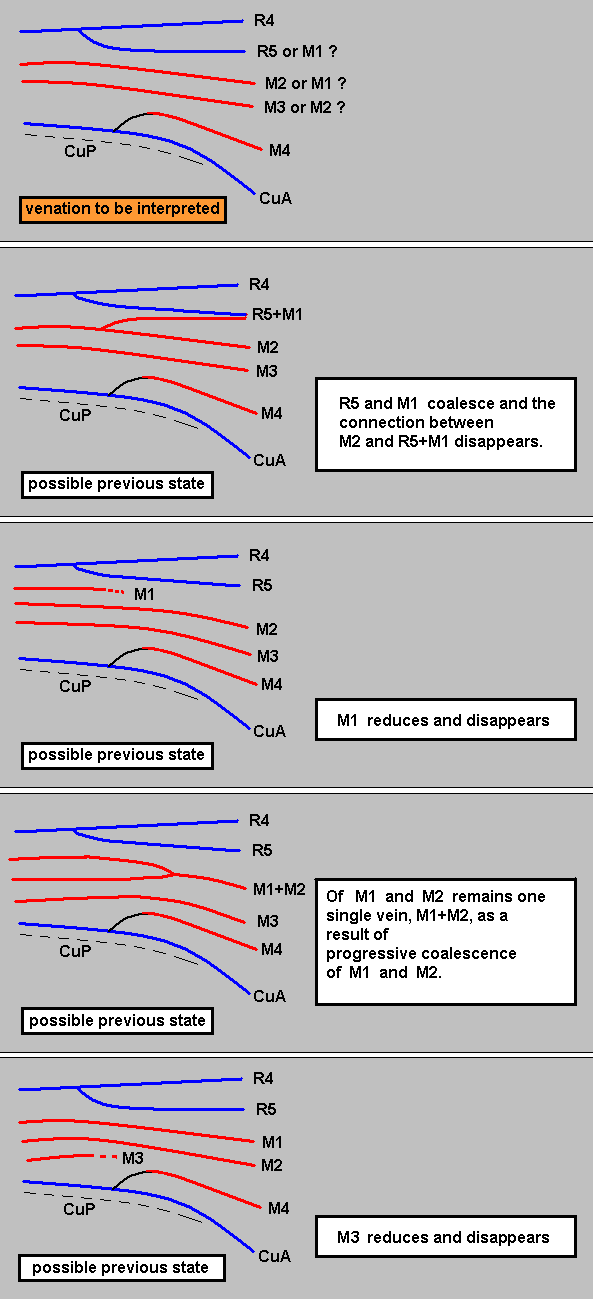
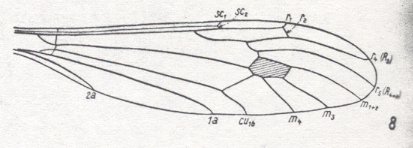
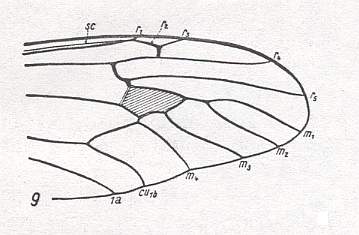
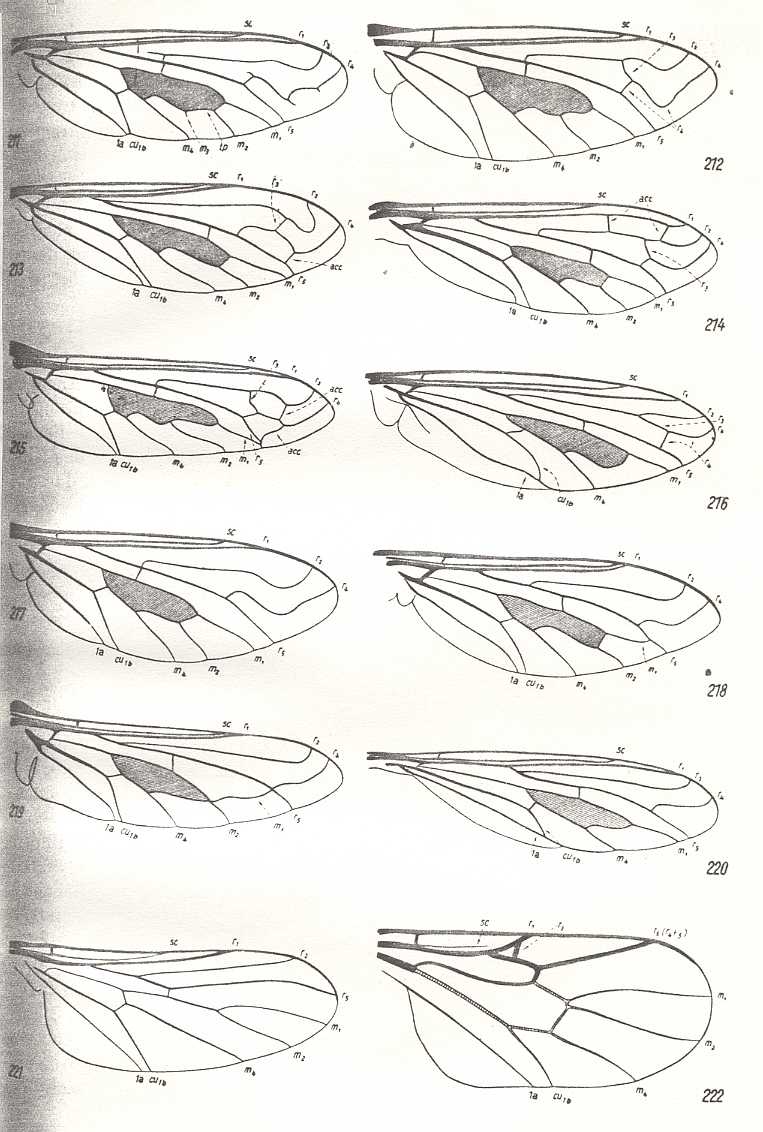
Summarizing the possible interpretions based on the transitional forms found :
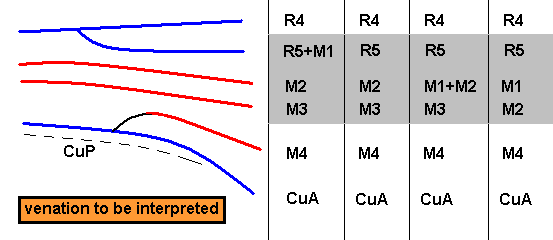
So depending on which transitional form actually exists, the veins in our example can be identified. And the equivocity is, of course only then removed when only one such transitional form exists. In fact the transitional form mediates between the condition in the Medial system in the Order-prototype (four Medial branches present) and that condition in our specimen to be venationally interpreted (three Medial branches present).
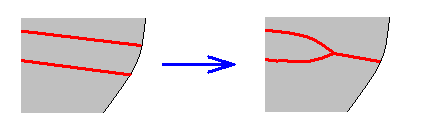
is a true derivation, and not the other way around. And, generally, we must assume the absence of a particular vein (as compared with the Order-prototype) to be a derivative state of it, and not the other way around, although there do exist cases where evidently veins have been added, as we see it for instance in the Order Neuroptera (Planipennia) if we compare their wings with those of more primitive but related forms or with the all-insect venational prototype. Probably such derivational rules differ in different insect Orders, or even in different superfamilies or families of the same insect Order : In the Tipulidae (Diptera, daddy-long-legs), for instance, we see a tendency for the main junctions between wing-veins, and for the discoidal cell, to shift towards the apical part of the wing, as compared with the venational order-prototype. And from this tendency we may deduce that "shifting towards the apex, of these structures" is part of some rule of derivation. On the other hand, in Psychodidae (moth-flies, Diptera) we observe a shift of the main junctions of the veins (the discoidal cell is absent) towards the wing-base. So here the corresponding rule of venational derivation apparently is the other way around.
Of course we could consider the two mentioned venational transformations to be functionally, not formally, determined. In that case they would then be adaptive, not governed by formal rules. However, there are a number of Tipulidea and Psychodidea (in which superfamilies the Tipulidae and Psychodidae respectively belong) in which such shiftings have not occurred or only slightly so (for instance in Trichocera [Trichoceridae], Tricyphona [Limoniidae], and Nemopalpus [Psychodidae] ), and thus as such pointing to the non-adaptive nature of these shiftings : The wings of these forms, not showing the shift, belong to the same functional type, respectively the tipuloid and psychodoid wing-types, as do those in which it has occurred.
So if indeed the derivational rules may sometimes change from insect group to insect group (but not from species to species), they, and thus the venations concerned, partly come from the qualitative content commonly possessed by the representatives of a number of different but related strategies forming such a group, that is, they are, partly, determined, not by the strategy sensu stricto (the latter as such representing one particular species and determining the precise way to survive in the Explicate Order), but by commonly possessed elements of the qualitative nature containing the strategy s.str. as its core. And then in groups which do not possess these elements the derivational rules are (a bit) different.
A particular functional wing-type (which might be one of the (main) types determined by Rohdendorf) is, qua qualitative content, intrinsically connected with the species' strategy (i.e. with that part of it that is commonly possessed by a number of different, but related, insect species) and is therefore, in the Implicate Order, automatically and necessarily implied by, and thus added to, the qualitative content of that strategy. But such a functional type is not more than only a generally, i.e. not completely, determined structure of the wing (and its basiala). And indeed : "not completely determined structure" can as such exist in the Implicate Order (while not in the Explicate Order). Further determination, supplementing and completing that structure -- the venational details -- comes in from without the strategy : Qualitative contents of other already completely determined venations -- existing as noëtic patterns in the Implicate Order -- noëtically influence the qualitative content of the functional (main) type : They noëtically react with it and are then incorporated into it in a correspondingly changed condition (forming a reaction-product), and so completing the determination of the venation, otherwise it cannot exist in the Explicate Order and thus cannot be projected. This changed condition of the venation or its details also is fully compatible with the qualitative content of the strategy concerned (but not necessarily implied by it) as a result of the integrating action of the latter content.
The described course of events in the Implicate Order is assumed here because of the observed existence -- in the Explicate Order -- of transitional stages between existing wing-venations, forming derivational sequences of them.
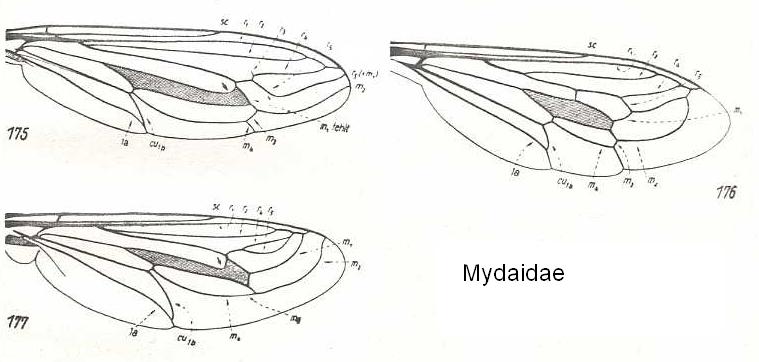
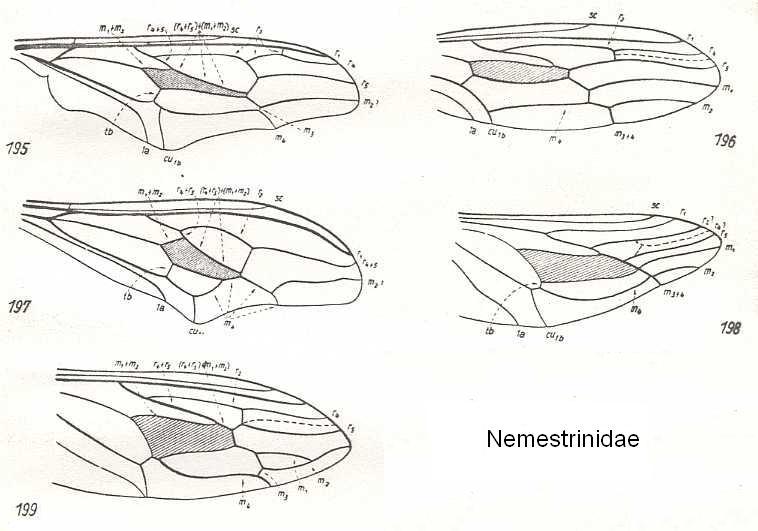
In all this, however, we should be aware of the fact that most families of, for example, Diptera do have a more or less characteristic overall pattern and distribution of veins (that is, each one of these families displays a typical-for-it wing-venation), such as in Nemestrinidae (all possessing the oblique composed 'vein', see HERE (and click on image) , or in Mydaidae (all possesing veins that, in the form of long arches, curve up towards the anterior margin of the wing-apex, see HERE , or in Syrphidae (hover-flies) (many of them possessing the 'vena spuria' and veins running parallel to the posterior wing-margin, see HERE , or in Psychodidae (moth-flies), having the main bifurcations of their veins shifted toward the wing-base see HERE (and click on image) , and in Tipulidae (daddy-long-legs), having them shifted toward the wing-apex, see HERE , HERE , HERE , HERE , and HERE .
These characteristic venational patterns, although not directly and completely being functionally connected with the very strategy of the insect, nevertheless reflect the purely morphological restrictions (and the corresponding noëtic restrictions in the Implicate Order) imposed by the qualitative content common to the species of such a family onto the structure of the wing-venation of them.
But also this is not rigorously maintained, as is shown in, for instance, the Bombyliidae (bee flies) where the wing-venation of a number of species strongly deviates from the 'characteristic bombyliid venation', see HERE (and click on image) , or even in Nemestrinidae, containing some species with a 'non-typically-nemestrinid' venation, see HERE .
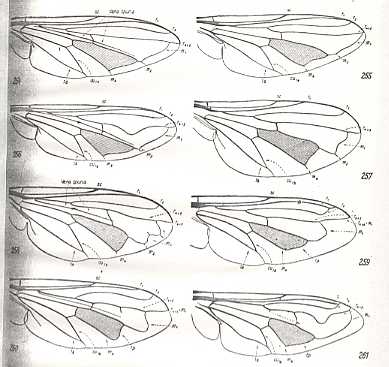
Further, if two identical or almost identical venations (or other structures for that matter) are supposed to have originated from that of two different species, not closely related to each other, then, in standard evolutionary theory, one speaks of (mere) convergence. That is, they are only identical qua venation. The species, on the other hand, possessing these identical venations, are nevertheless different, and are not supposed to be phylogenetically related to each other, and so the venation of them has not been inherited from that of some common ancestral species. See next Figure.
Convergence of wing-venation. 1. - Diadocidia spec. ( Diptera, Suborder Nematocera, family Diadocidiidae). 2. - Empidideicus spec. ( Diptera, Suborder Brachycera-orthorrapha, Family Bombyliidae). (After EDWARDS, 1926, in HENNIG, 1954)
In our theory, on the other hand, we consider these identical forms to be actually derived, not convergently from two different preceding forms not related to each other, but, at least as to their venation, directly or indirectly derived from each other, that is, the one from the other. And both belong to the same functional wing-type (in our example to the Ancient Lifting Venationally-attenuated (fungivoroid) Functional Wing-type). The derivation of one venational pattern from another (in the present case, 'convergence', involving only a slight, if any, difference between the original and the derived venation) is just a form of noëtic reaction, taking place in noëtic space (the Implicate Order).
Conclusion
With all this we have now established the status of the wing-venation in insects (and many other organ-systems of them also have a status similar to the wings and their venation). We have distinguished between (1) the functional core of wings and their venation, which core intrinsically belongs to the qualitative content of the strategy concerned, in this case to a layer of elements of it commonly possessed by a number of different but related species, and (2) elements of the venation that are extrinsic to that strategy, i.e. which are added, but which are nevertheless compatible with it and with the mentioned functional venational core, i.e. elements that complete that core, by fully determining the venation. These extrinsic venational elements can be empirically recognized on the basis of studying existing derivational sequences of venations within some given insect Order, as these sequences are imperfectly represented in the Explicate Order. Therefore, in the present series of documents on dipterous wings we always begin with the exposition of a number of functional wing-types and let it then follow by derivational sequences of relevant venations insofar as these sequences can be established from venations actually existing or having existed in the Explicate Order. (And in the previous document we already expounded a first set of funcional wing-types, and, in conformity to what has just been said, we'll investigate possible derivational sequences of the wings of these types in the next document).
Will be continued if necessary
In the next document we will try to find derivational sequences of the venations as found in the Tipulomorpha and the Bibionomorpha and relate them to the functional wing-types found in these groups of Diptera and expounded in the previous document.
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXXb.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII
Back to Evolutionary Part LXVIII
Back to Evolutionary Part LXIX
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}