
Having now determined the general biological and formal status of the wing-venation in insects, and thus their theoretical context, we are now ready to determine and investigate the possible derivational lines (sequences) as they may occur in the representatives of the functional types described in Part LXIX. In fact many such derivations have already been done in earlier documents, but the emphasis was there on the light they might throw on the evolution of the corresponding insect species. In the present document (and those to come), however, the emphasis is on the wing-venation itself and how it is derivationally related to other wing-venations (inside a given insect Order).
What we have called here "derivational lines" (with respect to the transformation of the wing-venation in given groups) are in fact the "morphoklines" discussed in Part XXII of the present Series on Evolution.
In discussing morphoklines (in the mentioned Part XXII) we read them from bottom to top, i.e. the derivational direction was from bottom to top along a vertical line. Here, however, when expounding derivational lines by actually deriving one venation from another, we will read, just by reason of convenience, from top to bottom, that is, the derivational direction coincides with scrolling down the table with venational drawings.
For a good understanding of the difference between the derivability of one venation from another and the actual descent of one taxon (species, genus, family) from another (taxa of which the representatives possess the wing-venations involved in the derivation), it is instructive to reproduce here what we have said about morphoklines of wing-venation in Part XXII :
The theory of morphoklines and the polyphyletic origin of groups and subgroups of Diptera.
A m o r p h o k l i n e is a more or less continuous series of morphological stages of a chosen organ-system (in our case wings) leading from one stage, via a series of intermediate stages, to a chosen 'end stage'. We will diagrammatize such a morphokline by a vertical line directed upwards. Certain existing (or hypothetical) wing-venations (venational patterns) are represented on such a line (morphokline) by points or nodes. The wing-venation in groups of Diptera is supposed to transform and develop along a bundle of parallel lines or morphoklines. Here we give a general scheme of such a bundle :
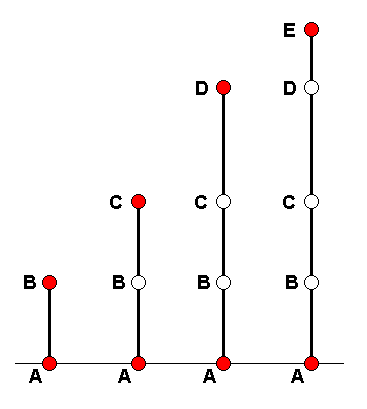
Diagram above :
A = starting venation, represented by the venation of an existing (or hypothetic) dipterous species or higher taxon.
B = second transformational stage. C = third transformational stage. D = fourth transformational stage. E = final transformational stage. The A's in all the lines represent identical or highly similar venations. The same applies to B, C, D, and E.
Vertical lines from left to right :
First line = morphokline of the wing-venation of B, where B is the venation of the wings of some existing species or higher taxon. Here the taxon represented by B has actually -- that is, historically - descended from the taxon represented by A.
Second line = morphokline of the wing-venation of C, where C is the venation of the wings of some existing species or higher taxon. The transformation has proceeded from A to C via the intermediate stage B. The taxon represented by C in this second line has not actually descended from the taxon represented by B in the first line. Only its venation (and not necessarily all other characters) has passed through the venational stage represented by B.
Third line = morphokline of the wing-venation of D, where D is the venation of the wings of some existing species or higher taxon. The transformation has proceeded from A to D via the intermediate stages B and C. The taxon represented by D in this third line has not actually descended from the taxon represented by C in the second line. Only its venation has passed through the venational stage represented by C.
Fourth line = morphokline of the wing-venation of E, where E is the venation of the wings of some existing species or higher taxon. The transformation has proceeded from A to E via the intermediate stages B, C, and D. The taxon represented by E in this fourth line has not actually descended from the taxon represented by D in the third line. Only its venation has passed through the venational stage represented by D.
As a result of the actual transformations having taken place separately and independently along the four lines, the wing-venation in the taxon E can formally be derived from that in the taxon D (third line), and this in turn from that in taxon C (second line), and this from B (first line), and B from A. (we return to this further below). However, this does not necessarily mean that the taxa themselves can be so derived, let alone that these taxa have actually, that is, historically, descended from each other. In almost all cases the representatives of such a group of taxa possess features -- other than wing-venation -- that do not allow us to assume that these taxa actually have evolved from one another (that is, see diagram above, taxon E [red point, 4th line] from taxon D [red point, 3rd line], and the latter from C [red point, 2nd line], and this one from B [red point, 1st line] ). (End of quote from Part XXII).
And -- still with respect to the diagram above -- concerning the just mentioned formal derivability of the venations ( E from D, D from C, C from B, and B from A) we must add here the following :
In the diagram above, it is assumed that in the first line (morphokline) the venation of (the representatives of the taxon) B (red point) is formally derivable (without any problems, and thus strictly so derivable) from the venation of A (red point, 1st line). On the other hand, as to the second line, not only has taxon C (red point) not actually descended from taxon B (red point) of the first line, but (of course) neither the respective wing-venations have actually descended one from the other. But the venation of C (red point) in the 2nd line may, however, formally be derived from that of B (red point) in the 1st line (these venations of C and B being, moreover, not hypothetical, not assumed, but venations of actually existing [or known to have existed] insect species), although in many such cases they will not be strictly so derivable, one from the other, as a result of specialization-crossings.
And this is where B, drawn as uncolored point, in the second line comes in : The taxon C (red point, 2nd line) has actually descended from the hypothetical taxon B designated by the uncolored point in the 2nd line -- this hypothetical taxon is supposed to be highly similar but not identical to the taxon B represented in the first line by the red point -- and therefore it is to be expected (certainly when the taxa C and B are species) that the wing-venation in taxon C (red point, 2nd line) can formally, and strictly so, be derived from that in the hypothetical taxon B (uncolored point, 2nd line). And, undoubtedly, such a taxon B (being highly similar to B in the 1st line), with such a wing-venation, has existed in the past (or still somewhere exists unchanged).
And what has been said here with respect to C and B, also goes for D and C, and for E and D.
In Part XXII all this is (also) explained in terms of a concrete example, explaining the relationship (including differences) between the derivations of one venation from another and the actual genealogical lines existing between species having these venations. From what has been expounded above, it is now clear that in fact every one single complete derivational line (derivationally connecting existing wing-venations) had to be split up into a number of different and parallel morphoklines in order to depict the actual genealogical sequences. And indeed this indicates that organisms (or at least diptera) have developed, not monophyletically, but polyphyletically.
We will now add to all this a general description of morphoklines derived (by us) from derivational lines (each of the latter giving a number of parallel morphoklines), in terms of ecology and of our theory of the Explicate and Implicate Orders (noëtic theory of evolution) as was already done in Part XXIII [there in the context of the example of the evolution of (jurassic) protorhyphoid flies all the way up into muscoid flies] [ This quote will be summarized and supplemented with extra considerations directly after it] [All what will follow is, as far as I know, nowhere published or discussed, it is completely new and therefore relatively hard to digest. But if the reader is really interested into the nooks and crannies of organic evolution he or she will happily give it a try!] :
As in all organic evolution, also in this evolution [from protorhyphids to muscoids] the successive invasions of new ecological niches (and with them of new zones) determine its course. This demands a further exposition of the potential ecological niche of a given species of organism. The representatives of that given species live in the actual ecological niche of that species. They are completely adapted to this niche by having 'internalized' certain key features of the environment. All this applies to at least all animal species.
What then is the "potential ecological niche" of a given organismic species, a species of, say, a fly?
First of all we must realize that such a species is a definite qualitative content. In the Explicate Order this content is represented by a number of individual instances, that is, here-and-now existing individual flies (adults or larvae). The existence of these individual instances means that the mentioned qualitative content is unfolded along space and time dimensions, resulting in that qualitative content to be observable in the form of existing material entities -- individuals -- each being spatially separated from one another, and each having its internal and external parts also spatially separated from one another. But these material entities can be " injected " (from the Explicate Order) into the Implicate Order (without losing them in the Explicate Order). They then are folded back along the space and time dimensions (they are enfolded or implicated). These dimensions now only appear as numerical parameters, and the result is a definite and purely qualitative content which we call a "noëtic entity" residing in the Implicate Order.
The potential ecological niche of the given organic species is also a qualitative content (in the Explicate Order represented by existing individual instances of it). It is a qualitative content because it is definite. And it is definite because it is precisely determined by something which is itself a definite qualitative content, namely the given organic species whose potential ecological niche we are now discussing. And in addition to this it is co-determined by some general features of the environment.
But how is this potential ecological niche so determined (and thus precisely delimited)?
For this we must consider the existing individual characters or features of the species, the potential ecological niche of which we are now considering. If these features change, the species generally is transformed into a new species and is adapted to a new ecological niche. Any given character (for example a feature of the wing-venation, a feature of the antennae, or some feature of the digestive system [of the larva or of the adult] ), can, evolutionarily, change only in certain definite ways or in a certain definite direction. For example, such a change, that is, such a transformation of the present state of a given feature into a new state of it, cannot be such that it is an exact evolutionary reversal to an earlier state of it, that is, a transformation back to a state-it-was-in already earlier in the history of the organism carrying this feature. Also those transformations that are incompatible with the present morphology or physiology of the organism, that is, transformations of which the proper initial state happens to be absent, cannot take place. And, of course, all transformations necessarily resulting in an unstable or unfit organism cannot take place, or, when they do so, their carriers are weeded out by natural selection. Further, the individual features of a given species are connected with each other in such a way that an evolutionary change in just a single feature, while all other features remain what they were, does not generally take place in organic evolution. So in one evolutionary step of the species actually taking place, a whole system of existing features of that species changes in such a way that the change in one given individual feature is coherent with the changes of all other features of that species. All this, consequently, determines [and restricts] how the representatives of the species will look like after one evolutionary step. It has, according to its new, but still only potential, qualitative content, become a (potentially) new species [but not yet completely so, because the mentioned first small (potential) evolutionary step consists only of the beginnings of adaptive transformations]. And this new qualitative content -- still in its state of potential, that is, not actual, qualitative content -- in turn determines (not by causing, but just by qualitatively implying) in what environment the (new) species -- if it represented this new content -- can live, or, more precisely, in what new ecological niche it can live [or better, this slightly changed content shifts the original species qualitatively and ecologically a little closer to such a new ecological niche]. The new, but still only potential, qualitative content thus connects as it were with elements of the existing overall environment, that is, it demands such elements. And if these demanded environmental elements actually turn out to be present in the existing environment (in the broad sense) we have finally found the sought-for potential ecological niche of our species. And indeed, every individual instance of this new niche represents its definite qualitative content. That content is definite because that content is precisely determined and delimited. It is so delimited by the potential form and structure of the original species (how its representatives would look like after one single [feasible] evolutionary step), which potential form and structure in turn was determined by the actual form and structure as it was present in the representatives of that species. And because it (i.e. the potential ecological niche) is, accordingly, delimited and definite, it is a genuine qualitative content (in the Explicate Order [still] existing in the form of individual instances), and can, and will, as such be injected into the Implicate Order (that is, a copy of it is so injected) where it then exists in the form of a noëtic entity, enfolded along space and time dimensions. The potential form-and-structure ( = not the actual, but potential qualitative content) of the original species -- in fact a (not yet existing) form that has acquired the appropriate beginnings of adaptations [these beginnings themselves not yet adaptive] to the potential ecological niche (which does exist) -- will not be injected into the Implicate Order because it is not (yet) an existing unfolded qualitative content (So there is nothing to inject). (Let us make this more clear by expressing it a little differently : The potential ecological niche is not some niche that does not yet exist. It is potential only with respect to the species under discussion, that is, it is not yet the ecological niche of that species. But all the environmental elements together constituting the pattern which we later call "ecological niche" are actually present (in the Explicate Order), therefore it can and will -- as a determined pattern -- be injected into the Implicate Order. On the other hand, the potential form-and-structure of the original species, that is, the new species it can become, does not yet exist (in the Explicate Order), and consequently cannot be injected into the Implicate Order.). So what is injected from the Explicate Order into the Implicate Order is : (1) the qualitative content of the original species, (2) the qualitative content of the actual ecological niche of that species, and (3) the qualitative content of the potential ecological niche of that species.
These injections result in the presence in the Implicate Order of their respective noëtic counterparts. And so we now can point to one of these noëtic entities, namely the one which in the Explicate Order exists as the potential ecological niche of our species, and this noëtic entity can noëtically react with another noëtic entity, namely the injected qualitative content of the original species. The (timeless) product of this reaction is, of course, also a noëtic entity. It is a new qualitative content having resulted from the noëtic integration of the two mentioned noëtic reactants : (1) the qualitative content of the original species and (2) the qualitative content of its potential niche. This new qualitative content, that is, the product of the noëtic reaction, is the qualitative content of the new species that has now acquired the appropriate and complete adaptations to the potential ecological niche of the original species. So far this new species still does not exist.
However, if now in the Explicate Order there exists a number of individuals of the original species that have access to at least one of the existing individual instances of the potential ecological niche of that species, the mentioned new qualitative content, that was noëtically generated in the Implicate Order, will project into the Explicate Order, that is, it will unfold itself along space and time dimensions (as they were specified in it in the form of numerical parameters), resulting in individual instances of the new species. In the Explicate Order this 'event' of unfolding (of the noëtic reaction [stages] and of the [final] noëtic product) will be 'observed' as the successful invasion by members of the original species of a new ecological niche by way of developing the appropriate adaptations (resulting in this original species to become a new species). ( End of quotation from Part XXIII ).
To further work out and clarify (but also a little modifying) these theoretical ideas we need to summarize the above considerations and add more details. Thereby four points are important from the outset :
Having mentioned these four important points, we can now work out and further clarify (and change a little) the above theoretical ideas.
We depart from a given organismic species existing in the Explicate Order. As such this species is represented in the latter Order as a collection of material individuals living in, i.e. integrated with, their species-specific ecological niche (which itself exists in the form of individual cases of it, and of related ones, within a given ecological zone).
The ecological niche of a given organismic species is a set of environmental existential conditions, and the adaptations of the individuals of the species consist in the fact that the mentioned conditions are, in a way, 'internalized' into the morphology, physiology and behavior of these individuals. The organismic species, as materially existing in the Explicate Order, is, as such, the meaning of the noëtic description of the complete qualitative content -- in the Implicate Order -- of the species. This content consists of the adaptive part of it -- the strategy -- as well as of the non-adaptive part of it. Taken together, we may call it the "strategy sensu lato" (s.l.). It is the complete qualitative content of the species. And the actual meaning of this noëtic description is generated by the projection of it from the Implicate Order into the Explicate Order.
Well, the noëtic description of the qualitative content of the species may subsequently undergo small changes of certain details of it, and we here devote special attention to those (small) changes that -- from a later perspective -- constitute the moderate beginnings of new adaptations [These changes we, as observers, describe in the Explicate Order as possible transformational steps in the morphology of certain organ-systems of representatives of the original species, or as possible non-random genetic mutations for that matter]. These possible changes cannot just be any (small) changes whatsoever, but are determined-to-be-possible by the very nature of the qualitative content of the original species. So -- and here we deviate a little from the above given quotation from Part XXIII -- the description, in the Implicate Order, of the qualitative content of the original species noëtically implies a number of other descriptions, descriptions of potential and little-changed qualitative contents, each one of them being slightly different from the original description, different that is, in certain more or less simple features, forming the beginnings of some possible new adaptational structures (morphological or behavioral). And such a potential (but as such existing -- because noëtically implied -- in the Implicate Order) little-changed qualitative content in turn noëtically implies a possible existential condition, a condition for it to exist in the Explicate Order. In fact this little-changed content moves the potentially new species closer to its proper ecological niche, so that the noëtic implication of the latter can indeed take place [This implication is driven by the supposed fact that every immaterial pattern whatsoever, residing in the Implicate Order, 'aspires' ontological completion, that is, it becoming ontologically received in (prime) matter, which is accomplished upon projection of it into the Explicate Order]. And this implied existential condition is in fact the noëtic description of a proper potential ecological niche, a niche that may receive, in the Explicate Order, upon projection, the mentioned potential little-changed qualitative content of the original species [which itself is not exhausted in all this]. In fact, however, the mentioned potential ecological niche (initially merely present as a noëtic description in the Implicate Order) cannot yet sustain this little-changed qualitative content, because such potential changes, having resulted in the latter content, do not yet have an adaptive nature. They, as has been said, constitute merely some beginnings of adaptive structures [Every such "beginning", itself far from being any particular adaptation, lies on the way that, when followed, ultimately leads to that adaptation]. So although the proper potential ecological niche -- the alternative niche -- is noëtically delimited, its 'internalization' into the qualitative content of the new species has not yet been accomplished. That is, at this stage the internalization is not necessarily implied. What we in fact say here is that an adaptation of a given entity A to another entity B cannot be accomplished by A alone, because it is an adaptation to B which originally is extrinsic to A. So in order to acquire this adaptation, A must actively involve B, that is, internalize it, and not only having it merely implied. Thus -- to return to our case -- what now, 'subsequently' takes place in the Implicate Order is a noëtic reaction between two reactants, namely between the description of such a potential little-changed qualitative content of the original species and the noëtically implied description of a possible new ecological niche. And the product of this reaction will be a noëtic description of the qualitative content of a new and complete species in which the content of the strategy-part of the old species has changed into another. And the result of this change of strategy is in fact the mentioned internalization of the potential ecological niche, it is the proper adaptation of the new species to the new ecological niche (by way of internalizing it). So now, in the Implicate Order, we have, in addition to the description of the qualitative content of the original species (which at the same time materially exists in the Explicate Order), the description of the qualitative content of a new organismic species fully adapted to a potential, but definite, ecological niche. And as soon as this niche actually and materially comes (or happens) to exist in the Explicate Order, and is ecologically adjacent to the ecological niche of the original species, the mentioned description of the qualitative content of the new species will acquire its actual meaning, that is, it will be projected from the Implicate Order into the Explicate Order.
And all this, we, as observers, 'see' taking place in the latter Order (with the help, of course, of conventional theory) as some given existing organismic species invading, with one or more of its populations, new territory, that is, invading and definitively occupying a new ecological niche and subsequently acquiring the necessary morphological/physiological/behavioral adaptations to it (as a result, according to conventional theory, of random genetic mutations and natural selection).
At last we have succeeded in describing organic evolution, not only in terms of the Implicate and Explicate Orders, but also in terms of non-random changes, and at the same time having succeeded in having shown how -- also in our theory -- species may evolve from other species, and most importantly, that organic evolution is driven by ecological factors, but that the existing adaptational features of organisms are not 'made' in (and by the forces of) the Explicate Order, that is, are not physically constructed, but come from noëtic 'activities' in the Implicate Order, guaranteeing the 'rational' aspects present in most adaptations in organisms.
It is precisely in this very context of : (1) ecology, (2) qualitative nature or content of a given organic species, (3) actual ecological niche of a given existing species, (4) its potential ecological niche, (5) actual and potential ecological zones, (6) injection into, and projection from, the Implicate Order, (7) noëtic reactions, etc., etc., that we must see and evaluate all the derivational lines and the morphoklines derived (by us) from them, along which wing-venations or other structures are derived from others.
In order to adequately apply the theory of morphoklines to the historical development of winged insects, especially of the Order Diptera, we must add still more to what has been stated above.
Now we must clarify the precise relationship between morphoklines, derivation, and the Implicate Order.
For this, let us depart from the above diagram of morphoklines, with some slight changes :
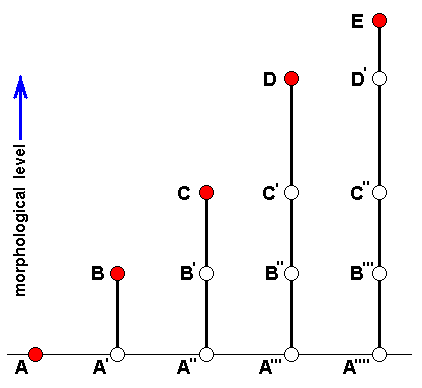
Diagram of Morphoklines : Each vertical line is a morphokline (connecting nodes), that is, a derivational sequence of species, and therefore of all species-constant characters [In practise, however, a morphokline (and also a derivational line) deals with only a single character and its transformations (and then generalized for all species-constant characters)]. Such a morphokline must be read from bottom to top. Each node on a morphokline is a species (including all its species-specific characters). Of these species the ones symbolized with red nodes actually exist or have so existed in the Explicate Order. The species symbolized with uncolored nodes are hypothetical (including their species-specific characters).
A', A'', A''', and A'''', are similar to A, but slightly and increasingly differing from it.
B', B'', and B''', are similar to B, but slightly and increasingly differing from it.
C', and C'', are similar to C, but slightly and increasingly differing from it.
D' is similar to D, but slightly differing from it.
B, although approximately derivable from A, is not strictly derivable from it. Therefore we must derive it from A'.
C, although approximately derivable from B, is not strictly derivable from it. Therefore we must derive it from B'.
D, although approximately derivable from C, is not strictly derivable from it. Therefore we must derive it from C'.
E, although approximately derivable from D, is not strictly derivable from it. Therefore we must derive it from D'.
So in the context of a diagram like this (which context is more or less epistemological) the uncolored nodes are hypothetical species (and their species-specific characters).
The series A==>B==>C==>D==>E we call a derivation line, established by the empirically determined fact that the character with which this particular line deals is such that, with respect to this character, E can approximately be derived from D, D can approximately be derived from C, C can approximately be derived from B, and B can approximately be derived from A. The species A, B, C, D, and E are species actually existing or having existed in the Explicate Order. Their approximate derivability from one another, with respect to the character mentioned, gives us an indication (but no more than that) of the nature of the transformations of this character in the ancestral lines of the species A, B, C, D, and E. And so, on the basis of this, we can set up morphoklines, i.e. a morphokline of each of these species. And the latter displays the polyphyletic evolution of the group (A, B, C, D, E).
Now we present these morphoklines as they are related to the Implicate Order. And this means that the diagrams depicting this are not epistemologically meant anymore (i.e. not about insofar we might know things), but ontologically meant (i.e. about insofar things really are, independent of being known) : There we have to do, not with how we might derive one structure (or species) from the other, but how they are actually formed from each other (involving noëtic descriptions of potential ecological niches, as described above). So what we are now going to express is (1) the observable history in the Explicate Order, that is, the successive appearance of organismic species in the Explicate Order, and (2) the noëtic or implicate history of them ( Here "history" in the sense of consecutive order of noëtic derivations [involving noëtic implications and reactions], and thus not involving Time) in the Implicate Order. As has been said in earlier documents the latter history can partly be uncovered with the methods of Phylogenetic Systematics in the sense of HENNIG (1954, 1966, 1969). But as we know, these methods are seriously hampered by the existence of the so many specialization-crossings between species or taxa to be derived from each other.
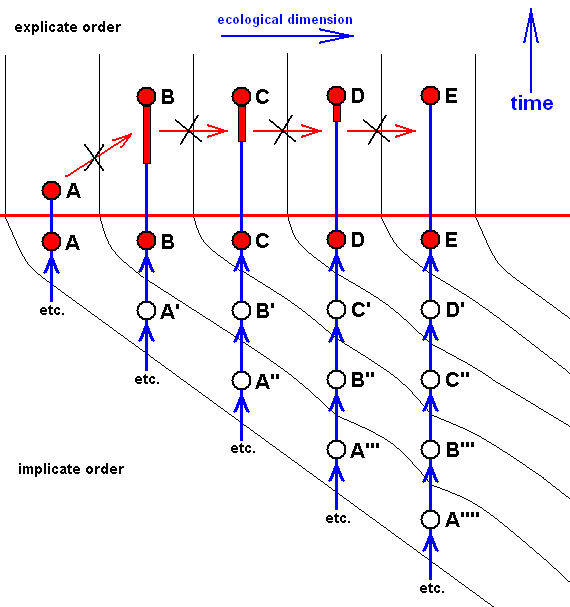
Diagram of the above morphoklines as they are in fact situated in the Implicate Order :
Here the case of only approximate derivability, and, consequently, not strict derivability -- in the Explicate Order -- of E from D, D from C, C from B, and B from A. This is indicated by the crossing-out of the red arrows drawn between them.
The red horizontal line separates the Explicate and Implicate Orders. Above this line the Explicate Order, below it the Implicate Order. As we see, the above four morphoklines (previous diagram) completely reside in the Implicate Order. The noëtic descriptions (of strategies to exist in the Explicate Order) formed in the Implicate Order (as expounded above) are projected (blue vertical lines) into the Explicate Order, and maintain from then onwards a continuing alternation between injection and re-projection. The height of the bottom-end of the downward red extention of the red nodes, B, C, and D (in A and E it is zero) and the elevation of nodes not having such extensions (A and E) indicate the time of first projection of these strategies (which corresponds to the time of first appearance of the species in the Explicate Order), while the (height of) their upper end signifies their last projection, that is their extinction in the Explicate Order. This applies to the species A, while the species B, C, D, and E are supposed to be still living today. So in the present diagram species A first appeared in the Explicate Order, but got extinct soon after that (i.e. cessation took place of its injection-reprojection-reinjection-reprojection etc.). Then species B appeared and remains all the way up to the present day, then species C, also persisting up to the present day, then species D and species E, both also persisting up to the present day.
In the Explicate Order we have, in addition to the space dimensions, also the time and ecological dimensions. The upper part of the diagram spans between the latter two dimensions. The strategies A, B, C, D, and E are projected into their (niche and) corresponding material ecological zones in the Explicate Order, separated in the diagram by thin vertical lines. The corresponding consecutive oblique zones in the Implicate Order mark the noëtic descriptions of actual and potential ecological niches (not individually delimited) (potential existential conditions) of the respective species (noëtic descriptions of strategies), potential ecological niches, together forming zones. The noëtic descriptions of the strategies A, A', A'', A''', A'''', for example, each of them include the description of their full integration with the proper existential conditions (upon projection, the actual material ecological niche). These noëtic descriptions of the existential conditions of the mentioned five strategies together form a zone in the Implicate Order as drawn. And, as has been expounded above, each strategy determines its potential changes forming a first evolutionary step, and in turn implying the noëtic description of the potential ecological niche of that strategy. And all the descriptions of these potential ecological niches together form the next zone. Noëtic reaction between the description of a strategy and the description of its potential ecological niche results in the description of a new strategy completely integrated with the description of the potential ecological niche which description now has become that of its actual ecological niche ( The [description of the] potential ecological niche of the original species has become the [description of the] actual ecological niche of the new species). So the strategies A', A'', A''', and A'''' noëtically transform (without exhausting themselves) into, respectively, the strategies B, B', B'', and B''' (while strategy A is directly being projected into the Explicate Order). And these in turn will determine their potential ecological niches and transform into the strategies C, C', C'' (while strategy B is directly being projected into the Explicate Order), and these in turn are transformed into the strategies D and D' (while C is directly being projected), and the latter (D'), finally, transforms into the strategy E ( D and E both being projected).
In terms of observable history of the organism-group (A, B, C, D, E) we see, as has been said, species A appearing first, and soon becoming extinct, then, one after the other, the appearance and persistence of the species B, C, D, and E. What we do not see (observe or deduce from theory), on the other hand, is the species A, B, C, D, and E actually descending, one from the other, that is, we do not see an ancestral tree (simply because there isn't such a tree). We only see -- in the Explicate Order -- the approximate (and thus not strict) derivability, one from the other.
The implicate history (= noëtic history) of A (looking backwards) is : A, ---, ---, ---, (etc.) (where ", ---, ---, ---," means that the implicate history, of course, can be traced further back, but that we do not do that here.) (see diagram ).
That of B is : B, A', ---, ---, ---, (see diagram ).
That of C is : C, B', A'', ---, ---, ---, (see diagram ).
That of D is : D, C', B'', A''', ---, ---, ---, (see diagram ).
That of E is : E, D', C'', B''', A'''', ---, ---, ---, (see diagram ).
So in the present case the group (A, B, C, D, E) has -- in the Implicate Order -- evolved, not mono-, but polyphyletically (as far as having it followed back into noëtic history), although the original species A, A', A'', A''', and A'''' are here supposed to be very similar to each other (expressed by the fact that they are connected with the same ecological zone (as represented in the Implicate Order).
If some given species of winged insects are, as to their morphological/physiological/etc. constitution, actually derived (in the Implicate Order), one from the other, then, by implication (in fact by definition) all their species-constant features, and thus also their wing-venation, are also so derived, one from the other. But of course, if we can -- now epistemologically thinking -- derive one given wing-venation from another, that does not prove that the respective insect species have actually derived from each other, even species of the same group such as a family. So, when setting up derivational lines of wing-venation present in such a group, that is, ordering the existing species of this group according to the consecutive derivability of wing-venation, we subsequently derive the corresponding morphoklines of wing-venation, morphoklines, that is, of each of these existing species (and thus such a morphokline containing several venations). And such a morphokline may, or may not, be, at the same time, the morphokline of the final species situated at its top (i.e. its end), reflecting the noëtic descent of that species successively from the other species of the morphokline.
Continuing with the above diagram and further developing things :
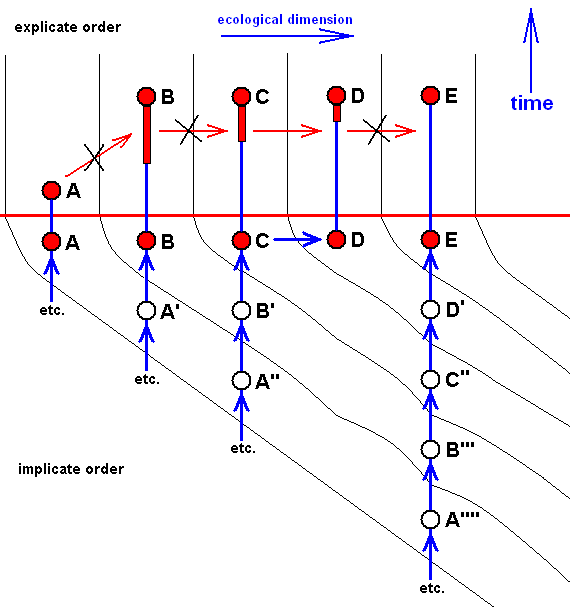
Diagram of morphoklines as they are in fact situated in the Implicate Order :
Case of not strict derivability -- in the Explicate Order -- of E from D, C from B, and B from A, but definitely of strict derivability of D from C (indicated by the not crossed-out red arrow).
This derivability of D from C is (as can be concluded) based on the actual derivation -- in the Implicate Order -- of the noëtic description of strategy D from that of C (indicated by the blue arrow), accomplished by the now-already-familiar-to-us noëtic process : The noëtic description of strategy C determines (while itself -- without exhaustion -- at the same time being projected into the Explicate Order) the noëtic description (also without exhausting the original description) of that strategy how it would be one evolutionary step further (beginnings of a possible adaptation to some new ecological niche), which description in turn determines the noëtic description of the potential (new) ecological niche. And this latter description noëtically reacts with the description of the little-changed original strategy, resulting in the description of a new strategy, D, this description now including the complete set of features adapting it to the new ecological niche.
In terms of observable history of the organism-group (A, B, C, D, E) we see species A appearing first, and soon becoming extinct. Then, one after the other, we see the appearance and persistence of the species B, and C. Then we see species D evolving from species C (without exhausting the latter), and finally we see species E appearing. In fact, we do not actually see species D originating from species C (because that takes too much time), but we conclude to it basing ourselves on empirical circumstantial evidence. What we do not see (observe or deduce from conventional theory), on the other hand, is the actual formation of the species A, B, C, and E.
Here the implicate history of A (looking backwards) is : A, ---, ---, ---, (see diagram )
That of B is : B, A', ---, ---, ---, (see diagram ).
That of C is : C, B', A'', ---, ---, ---, (see diagram ).
That of D is : D, C, B', A'', --- (see diagram ).
That of E is : E, D', C'', B''', A'''', ---, ---, ---, (see diagram ).
So also in the present case the group (A, B, C, D, E) has -- in the Implicate Order -- evolved, not mono-, but polyphyletically (as far as having it followed back into noëtic history), but the number of initial species is now smaller (four instead of five) than in the previous case : it now consists only of A, A', A'', and A''''. On the other hand, the group (C,D) is monophyletic. Its most direct common noëtic ancestor is B' (and its more remote one A''). And it is clear that species C and D are members of one and the same morphokline : A'', B', C, D, which can, alternatively, be drawn as a straight sequence of nodes. The difference with normal morphoklines consists in the fact that its last two members have both been projected into the Explicate Order.
So thanks to the general possiblity of monophyletic groups of organisms (also in our theory) we do not need, to assume, for instance the descent of the 80000 species of Diptera from 80000 earlier species of Mecoptera (Scorpionflies). A wealth of Mecoptera (of species as well as of individuals) that was the same as in the later Diptera, and which is also maintained when we descent further back into history, is difficult to imagine. But if monophyly nevertheless turns out to be a rare phenomenon, or perhaps not existing at all, then we indeed would have at least 80000 mecoptera species having given rise to diptera (and, in addition, still other mecoptera species having given rise to caddis-flies (Trichoptera)). It must, however, be realized that speaking about the number of species evolving into other species is speaking about events taking place, not in the Explicate Order, but in the Implicate Order, where from certain noëtic descriptions of strategy others are derived. And in noëtic space there is no crowding of species and individuals. And in the Explicate Order there will also be no such crowding because it is to be expected that only a fraction of the -- in the Implicate Order -- noëtically forged descriptions of strategies will actually be projected into the Explicate Order (and appear there as organismic species), because projection depends on the actual existence -- in the Explicate Order -- of proper environmental conditions, on the actual existence, that is, of the precise ecological niches to which these strategies are precisely geared.
Let us continue our exploration of derivabilty of existing forms from other such forms and the interpretation of it in terms of the Implicate Order.
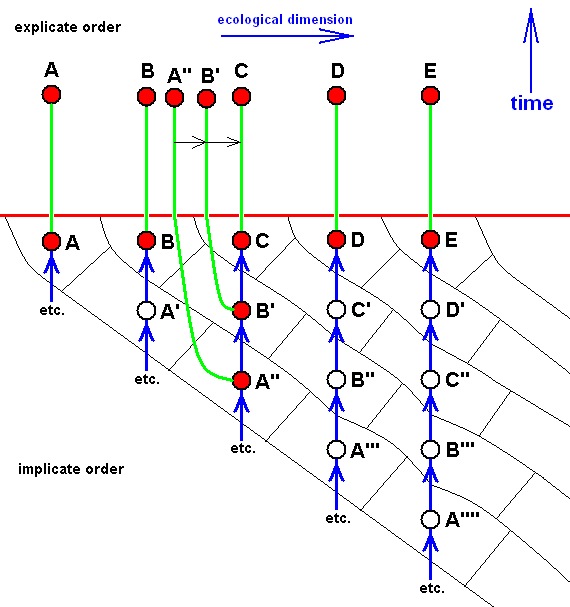
And if, in addition to the strict derivability -- in the Explicate Order -- of D from C, also C would strictly be derivable from B, then we have :
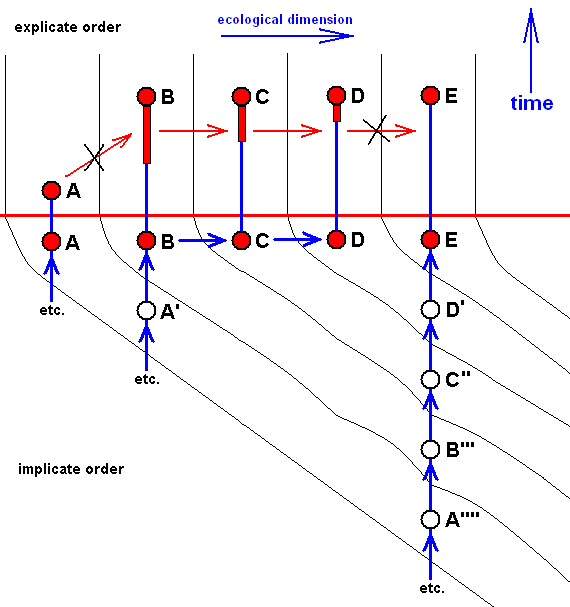
Diagram of morphoklines as they are in fact situated in the Implicate Order :
Case of not strict derivability -- in the Explicate Order -- of E from D, and of B from A, but definitely a strict derivability, not only of D from C, but also of C from B.
Here it can be assumed that the strict derivability -- in the Explicate Order -- of the species D from C, and of C from B, is based on the actual derivation -- in the Implicate Order -- of the strategy D from C, and of C from B, a derivation accomplished by the above expounded noëtic process.
In terms of observable history of the organism-group (A, B, C, D, E) we see species A appearing first, and soon becoming extinct. Then we see the appearance and persistence of the species B. And from species B we see evolve species C, and from C we see evolve species D, and finally we see species E appearing. In fact, we do not actually see species C originating from species B, and species D originating from species C (because that takes too much time), but we conclude to it basing ourselves on empirical circumstantial evidence. What we do not see (observe or deduce from conventional theory), on the other hand, is the actual formation of the species A, B, and E.
Here the implicate history of A (looking backwards) is : A, ---, ---, ---, (see diagram ).
That of B is : B, A', ---, ---, ---, (see diagram ).
That of C is : C, B, A', ---, ---, ---, (see diagram ).
That of D is : D, C, B, A', ---, ---, ---, (see diagram ).
That of E is : E, D', C'', B''', A'''', --- (see diagram ).
So also in the present case the group (A, B, C, D, E) has -- in the Implicate Order -- evolved, not mono-, but polyphyletically (as far as having it followed back into noëtic history), and the number of initial species has now decreased still more : it now consists only of A, A', and A''''. On the other hand, the group (B,C,D) is monophyletic. Its most direct common noëtic ancestor is A'. And also here the species B, C, and D belong to one and the same morphokline : A', B, C, D, which can be drawn as a straight sequence of nodes. But here the last three members of this morphokline have been projected.
The above diagrams depicting morphoklines give the most simple cases of the system of noëtic reactions, derivations and successive projections : the 'final' strategies are -- in the Implicate Order -- each produced through a step by step process, and then projected into the Explicate Order. But of course these 'final products' (the end-points of the morphoklines) are not "final products" at all : First of all it is to be expected that the production and projection of new strategies will continue beyond the 'final products'. And moreover, every noëtic reaction, whatsoever, taking place between a noëtic description of a given strategy and the noëtic description of its potential ecological niche results in the noëtic description of a new, and especially complete, strategy, a noëtic description that is, (ultimately) of a fully adapted new species. And these will in turn give new species themselves, etc. So precisely which ones of these new strategies will be projected does not depend on the position of such a strategy on the morphokline, but only upon whether the proper ecological niche is actually present in the Explicate Order. And this means that our above diagrams have far from exhausted all possibilities.
Let us see how things look when in one of the morphoklines of our diagrams all of its members are projected into the Explicate Order (also here the order in which they are projected depends on existing ecological settings in the Explicate Order) :
In this Diagram we do not -- in the explicate-order-part -- explicitly distinguish between existing ecological settings anymore, but of course know that every species occupies its own ecological niche. Also we do not so distinguish between the points in time of first and last projection (that is when, in geological history, the species appeared and when it became extinct).
In the implicate-order-part of the present diagram, we explicitly distinguish between areas in noëtic space representing different (noëtic descriptions of) ecological n i c h e s, but do not distinguish between ecological z o n e s anymore [ That is, we do not let the long oblique lines delimit (noëtic descriptions of) different ecological zones anymore {Later, when we depict existing derivational lines in terms of morphoklines, we will do it the other way around again : we depict areas -- in the implicate-order-part -- standing for noëtic descriptions of ecological zones, and do not explicitly delimit the individual niches anymore}]. [Compare with the first of our above diagrams ]. Finally, all projections (with their subsequent injections and re-projections) are expressed by green lines.
In the case here depicted, of the morphokline A''- B' - C not only the strategy C is projected into the Explicate Order, but also all other members of it. This results in the fact that in the Explicate Order there are now (in addition to others) three species present (existing or having existed) that can, if known, be perfectly derived, one from the other, that is, C from B', and B' from A''. So in this case the noëtic history of the species C -- a species present in the Explicate Order -- is reflected in the species B' and A'' also existing in the Explicate Order. Of course, when the species C, B', and A'' are all recent species, they cannot, insofar as simultaneously existing, have been evolved from each other. As always, their history is a noëtic one, and of species C this history is, when descending, the noëtic description of the strategy C, the noëtic description of the strategy B', the noëtic description of the strategy A'', etc.
It can also be the case that the so-called last member of a morphokline is not projected into the Explicate Order (because of the absence there of the proper ecological niche) :
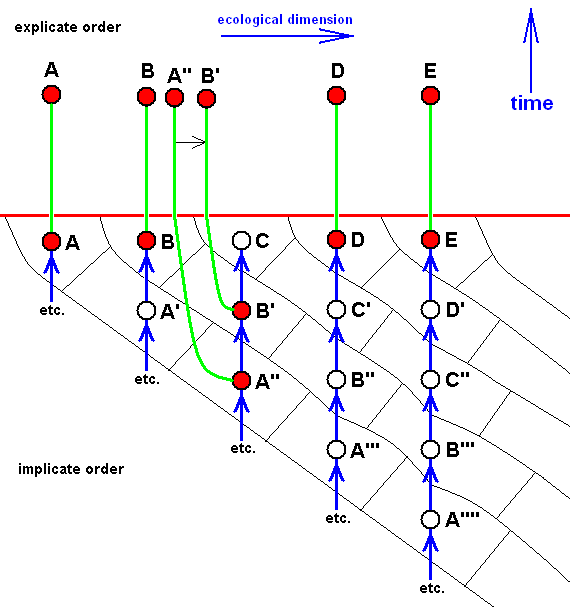
Here, of the morphokline C - B' - A'' only the strategies B' and A'' have been projected into the Explicate Order. So only the species B' and A'' have appeared in the Explicate Order.
Or, no members at all of that morphokline have been projected :
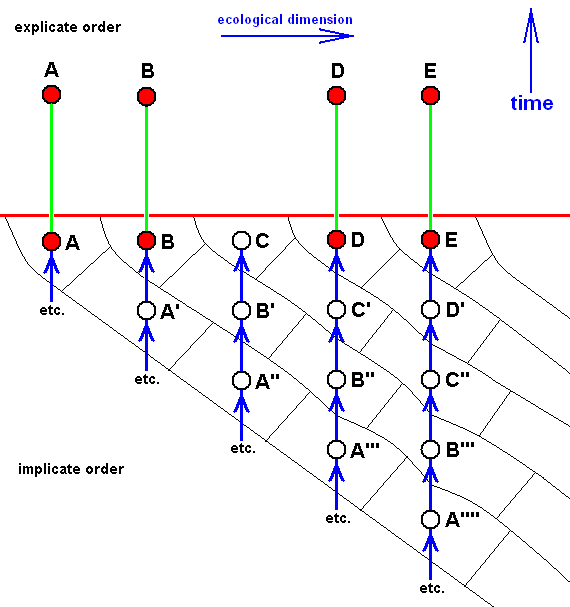
Here, no species of the (depicted part of the) morphokline C - B' - A'' have (until now) appeared in the Explicate Order.
Or, just, say, only two members of the whole group (A, B, C, D, E) have been projected :
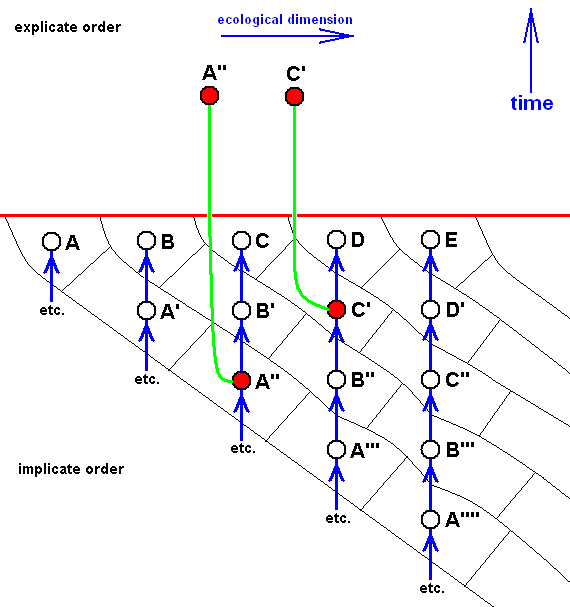
Here, only two species of the group (A, B, C, D, E), namely A'' and C', have appeared in the Explicate Order. This describes the often encountered fact of the presence -- in the Explicate Order -- of one or two species that are very isolated from any other group or species.
The noëtic (= implicate) history of C' is (descending) C', B'', A''', ---, ---, ---
With these general theoretical 'tools' we are now able to place the coming derivational lines of wing-venation in Diptera in their proper context. And remember, when finding a morphokline of wings (in most cases a -- mental -- reconstruction of it, based on approximate derivabilities of venations found in the Explicate Order), we have at least an indication that we have here at the same time to do with the morphokline of the species, that is, of the species at its end (top) (this morphokline being the sequence of derived -- in the Implicate Order -- strategies sensu lato (= the descriptions of the complete qualitative content of species), that is, the complete bundle of morphoklines of all species-constant characters). And in the mentioned derivational lines of wing-venation in Diptera we read (for reasons of convenience) the derivations from top to bottom, that is, from the original to the derived.
Let us, still in a general way, look into cases that can be expected when analyzing derivational lines of wing-venation. For this we will use selected sections from the above diagrams ( Explicate Order : above the red line. Implicate Order : below it. Green line = projection. Uncolored node = content not projected. Blue arrow = noëtic derivation). In all this, a projected strategy-structure (= a strategy-structure now existing as a material organismic species in the Explicate Order) is, as to its qualitative content, identical to the corresponding noëtic description -- residing in the Implicate Order -- that has been projected. The only difference consists in the fact that a projected strategy-structure now exists in the form of material individuals in the Explicate Order, instead of merely in the form of a qualitative content (immaterial form, noëtic description) in the Implicate Order. Derivability relations that might occur between such immaterial contents do not change upon projection of them into the Explicate Order. It must further be realized that we, as observers, can see the absence or presence of derivational relations only in entities having been projected into the Explicate Order, and from these observed relations we may conclude how they come forth from noëtic derivations in the Implicate Order, where these derivations are in fact noëtic generations (originations) of one noëtic description from another, constituting the noëtic history of the relevant species :
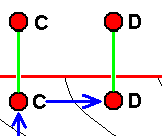
|
If wing-venation D is -- in the Explicate Order -- s t r i c t l y derivable from wing-venation C, then we hold that the fact of this derivability is the result of the actual origination of D from C in the Implicate Order. And by the same reason we may hold that C and D are at the same time species -- where species D has, also in the Implicate Order [and only there], evolved from species C, although this is not certain. It does become fairly certain when both wings, in addition to their strict derivability one from the other, do clearly stand out from other wings of representatives of the same group such as a family. |
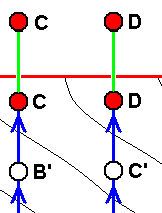
|
If wing-venation D is -- in the Explicate Order -- only a p p r o x i m a t e l y derivable from wing-venation C, then we hold that the fact of this incomplete derivability is the result of the actual origination -- in the Implicate Order -- of D not from C, but from C', where C' is not identical to, but nevertheless similar, to C. That D and C at the same time represent species might be the case but is less certain. C' is a hypothetical species (or venation), in fact a noëtic description of a strategy (s.l.) assumed to exist in the Implicate Order and not having been projected into the Explicate Order. This only approximate derivability (in the Explicate Order) of D from C forces us to assume a separate morphokline for D, separate that is, from that of C, meaning that D and C have separate noëtic histories at least for so far as having them traced back. There may, however, exist cases where we do not have to assume separate morphoklines, see image further below . |
And if (as in the next image) the just mentioned 'hypothetical species' (or venation) is subsequently discovered in the Explicate Order (either as a fossil or as a recent species), then we are certain that the strategy (s.l.) C' really exists in the Implicate Order, rendering it not hypothetical anymore [A given strategy (s.l.) truly existing in the Implicate Order, but not having been projected into the Explicate Order, is, as such, not a mere hypothetical strategy. It is real, in the sense of really being present in the Implicate Order (i.e. at least in this Order). But precisely that we will only know for certain when this strategy has been projected into the Explicate Order as a species (and subsequently having been found there)]. See next image.
And if C'' is actually found to be present in the Explicate Order, then it confirms the assumption :
Further, we might encounter the following case :
Generally we can say that wing venations-to-be derived, and even more so species-to-be-derived, often involve specialization-crossings, forcing us to push back the supposed common ancestor further and further down into the depths of noëtic history, increasing the probability that they have evolved independently of each other after all, that is, that they have evolved polyphyletically instead of monophyletically. Further, in wings and wing-venations, even within the confines of a single family, we have cases in which the wing-venation of a given species can perfectly be derived from that of another species, but that the overal shape and 'face' of the wings of this latter species are very different from those of the wings from which their wing-venation could be derived. In such cases, where we assume that such wings have in one way or another evolved from each other we do not know the direction of transformation of the shape and 'face' involved. The difference in the latter features might well point to the corresponding wings not having been evolved, one from the other. As a result of all this, it is perhaps safest to assume (at least initially) a polyphyletic (noëtic) descent of insects, or all organisms for that matter. And indeed, a noëtic descent it is, because we do not hold that species (and their features) evolve from other species in the Explicate Order. They noëtically are constituted, one from the other, in the Implicate Order. But although it is wise to depart from possible polyphyly (i.e. initially assuming polyphyly) in a given group, we know that monophyly does occur. So we are sure that the organismic world is not completely polyphyletic, but large segments of it may be.
Before we are going to discuss concrete derivational lines of existing (in the Explicate Order) wing-venations in Diptera, and to show the fact that each one of them involves a series of morphoklines that extend into the depths of the Implicate Order, it is wise to repeat here what we have said (much) earlier about the difference between these two Orders, especially with respect to the evolution of organisms seen as successive projections of (noëtic descriptions of) strategies from the Implicate into the Explicate Order.
In the Implicate Order there do not exist "individuals" and not "individual cases", neither do there exist particular "places" (in a strictly spatial sense), nor particular points in time. There is no "here" and "there", no "before" and no "after" (in a strictly temporal sense). The Implicate Order is just noëtic space where the dimensions are not spatial (neither temporal), but purely qualitative (they measure qualitative 'space'). The "ens extensum" (extended Being) is made up here by qualitative content which is in fact natural numbers and their relations to one another. They (these numbers and there relations) encript descriptions -- noëtic descriptions. The entities of the Implicate Order are, therefore, not material, not physical, but noëtic.
In the Explicate Order, on the other hand, everything exists in Space and Time. Therefore any given qualitative content there exists in the form of individuals or individual cases. This implies that all entities in the Explicate Order are physical and (thus) material. For something to be 'immaterial' there is only the result of deliberate abstraction by humans : " If it -- pointing to the alleged immaterial thing -- were not based in, or being a part of, matter". So in the Explicate Order certain entities, i.e. certain things, are "here", i.e. in a certain region, and "now", i.e. at the present point in time. Or they are, not here, but "there", and, not now, but "then". Seen from the Explicate Order a projection (from the Implicate Order) of something takes place at certain places (and not at others), and at certain points in time (and not at others). But seen from the Implicate Order it is just projected. If we say that a strategy is only projected when the proper existential conditions, to which that strategy is intrinsically geared -- which conditions are 'internalized' into it -- actually exist (in the Explicate Order), we mean this as seen from the Explicate Order, not from the Implicate Order.
Seen from the Implicate Order a given strategy (s.l.) is projected into the Explicate Order, that is, into its [i.e. the strategy's] existential conditions. Seen from the Explicate Order a new species appears in its proper environment. As such it appears here, in this region (i.e. in this particular 'individual' of the proper environment of that species) because its direct noëtic ancestor already existed nearby, or some population of it had migrated up to it (which is all part of the new species' "proper environment"). Projection of the new (but related) organic strategy (= appearance of a new [but related] species) is in the Explicate Order seen in the form of a population of the original species having invaded the mentioned environment and having, subsequently, adapted to it.
Derivational lines (morphoklines) of wing-venation.
For the Order Diptera we'll start with the tipuloid wings of the families Limoniidae, Tipulidae, and Cylindrotomidae.
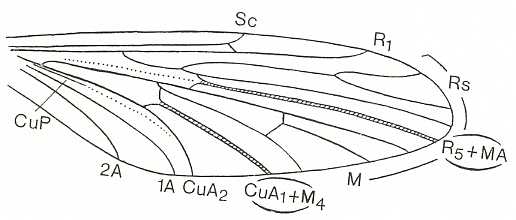
At the beginning of all derivation we depict here the venational dipterous Order-prototype from which all dipterous venations can be, directly or indirectly, derived. If we go around the wing margin from left to right and front to back we meet, in this prototype, the following veins : Subcosta (SC), Radius (R1), four branches of the Radial Sector : R2, R3, R4, R5 (the vein R4 is supposed to have merged with MA and is called "R5"), four branches of the Media : M1, M2, M3, M4 (the vein M4 is supposed to have merged with CuA1 and is called "M4"), CuA2 (also called just CuA), two anal veins : 1A, 2A. The cell in the center of the wing and enclosed by medial branches and the intermedial cross-vein (tp) is the so-called discoidal cell. The connecting vein between the Radial and Medial systems (here between R5 and M1+2) is the cross-vein ta, also called r-m. The connecting vein between the Medial and Cubital systems (here between M4 and CuA2) is the cross-vein tb, also called m-cu.
The first six derivational lines concern the wings of representatives of the family Limoniidae, studied in Part XXIX of the present Series. ( For the authors of the drawings, and also the authors of the species names, see the just mentioned Part). Without indication of geological age of the form pictured means that the species is recent. When a species does not belong to the recent fauna -- when it is fossil -- geological age and region where it was found are given.
In the "derivational lines" to come, we will, reading such a line from top to bottom, formally derive one venation from the previous one. Because most of the wing-venations depicted in these lines are from recent insect-species, these species, and with them the wing venation of their representatives, cannot of course, have historically descended, one from the other. So it would seem that there is no point in (formally) deriving one recent venation from another, because formal derivation to be possible depends on actual descent. Fortunately, however, organisms have evolved (or just changed) at different speeds. That's the reason why the recent fauna still contains many primitive forms. And these primitive but still living forms, when compared with related but more advanced living forms, give us an indication what kind of evolutionary transformations actually have taken place in the course of history. That's why indeed we may compare recent forms with other recent forms. And when we say that a recent species B has originated from a recent species A, we mean that the recent species B has originated from an immediate ancestor of species A, which ancestor is supposed to have but little changed or not changed at all. In the beginning of the present document all this is clearly explained with the concept of morphoklines, see the diagram (and its subscript), above .
In the venational morphoklines-to-come we have made use of drawings of wing-venation as they have been presented in the literature by specialists. And because morphoklines are supposed to express the derivational structure that objectively obtains in some given, naturally existing, whole set of wing-venations (as we see it, for instance, in the Order Diptera), it might be expected that the morphoklines we ourselve determine (in this website) on the basis of available material, suffer, not only from incompleteness, but also from often giving us a contorted picture of this derivational structure. Some species and their wing-venation not yet discovered, or the various venational drawings not in my possession, can be expected to change the derivational picture dramatically as soon as they have been discovered or are in my possession, and are studied in this respect. They may turn out to be "missing links" in derivational series, or they may refute one or two worked-out derivational lines. But in fact all this is not so bad as it looks. Confining ourselves to the Order Diptera, we can say that we possess the drawings of most, if not all, venations that may be important from a derivational viewpoint. Further, if we have missed a "missing link" or two, this does not matter so much, because the derivational relationships have been detected anyway, also without this link being available. The "discovered link" only confirms the existence of a derivational relation already found earlier between two compared forms. The only thing that might change as a result of the availability of additional drawings of wing-venations is that some given group of them, initially considered to be isolated because of a lack of derivational relations it shows with members of other more or less adjacent groups, now, on the basis of additional drawings having become available, turns out to be not so isolated afterall, but derivationally connected with one or more other groups. However, such "missing links" can, without having found them, be expected. They are then "hypothetical" venations (or species). And in other cases of presumed isolated groups it is rather easy to see that they are really isolated, and that discovery of missing links is not to be expected. So although small deviations from the truth may occur in our derivational lines it is not a serious matter. But that doesn't mean that we are not interested in having more venational drawings (especially of fossils) at our disposal.
Each derivational line, consisting of a number of wing-venations more or less formally derivable, one from the other, is followed by the corresponding diagram depicting the morphoklines. In these diagrams we enter the respective species with their genus name only (except where, in such a diagram, two or more species of the same genus occur). And in addition to the genus name we there represent the species also by a letter. Further, when two or more venations, depicted in a given derivational line, are perfectly, and that means strictly, derivable, one from the other, but belong to different insect genera (or higher taxa), we consider them to belong to different morphoklines. Only venations that all belong to the same insect-genus and are strictly derivable one from the other on all accounts, are considered here to belong to one and the same morphokline. Finally, in the explicate order part of the diagram we, as has been said, place the species of the derivational line. The wing-venations of the representatives of these species are more or less derivable, one from the other. But even when they are strictly so derivable, we may assume that this derivability does not hold for the species, each as a whole (i.e. with respect to all their morphological, etc. characters) or even with respect to the wing as a whole. Earlier we depicted this by a crossed-out red arrow between them, but in the diagrams that will follow we just place a not-crossed-out red arrow between two such species, and now this arrow signifies that the wing-venations are formally derivable, but that this not necessarily holds for the wing as a whole, let alone for the species as a whole. And although -- in the implicate order part -- we interpret the long oblique zones as each representing the noëtic description of a particular ecological zone, we do not explicitly delimit the individual niches anymore, but know that they do constitute such a zone.
Let us now, finally, start with the actual derivations.
Limoniid wings without discoidal cell.
LINE 1 :
These four wings are, it is true, formally derivable, one from the other. But the wings themselves, each one taken as a whole wing, are different from each other. Of course we may see these different overall 'looks' of the wings as part of their derivation, one from the other, but the suspicion arises that they form four separate and independent morphoklines, and that the presented wing-venations suggest how these morphoklines might look like [We must realize that in our theory all actual processes of derivation -- all true origination -- in all cases take place in the Implicate Order only] :
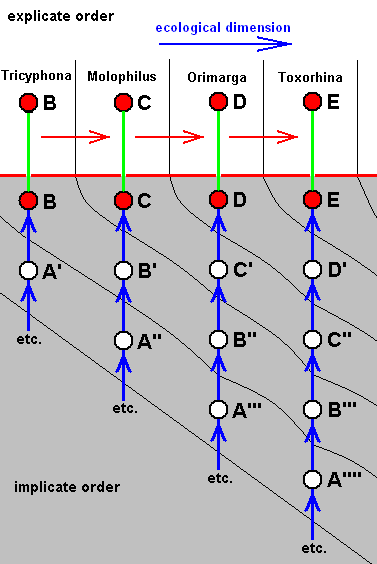
The four morphoklines of the venation of the limoniids Tricyphona, Molophilus, Orimarga, and Toxorhina (coded, respectively as B, C, D, and E ). Blue arrows indicate noëtic derivations, and here the red arrows indicate formal derivability of the venation in these genera. Green lines indicate projection from the Implicate into the Explicate Order.
Actual formation of these four existing venations, one from the other in the Implicate Order [That is one having been originated from its immediate noëtic ancestor] resulting in one morphokline only, is, however, not likely.
Thus what is not likely is the following :
So we must assume the existence of, not one, but four morphoklines. Thus, for example, in the Implicate Order the noëtic description C (corresponding, upon projection, with members of the genus Molophilus) cannot noëtically be derived from B, so it has actually originated (and thus noëtically derived) from B', which is similar (but not identical) to B. For the same reason B' cannot noëtically be derived from A'. It therefore noëtically originates from A''. Similar things hold for the other genera in this diagram.
Let us explain precisely what a diagram like the one just given (and generally the ones that will come) tells us, and what it does not.
In the above diagram, B, C, D, and E are existing organismic species, i.e. species existing (or having existed) in the Explicate Order. In the Implicate Order they exist as noëtic descriptions of strategies (indicated by the same letters) [For reasons of convenience we keep calling them "species" -- and the same goes for those that are not projected -- while in fact they are supposed to be noëtic descriptions of corresponding strategies [s.l.] that will, only upon projection, appear as true organismic species]. In both the explicate-order-part and the implicate-order-part of the diagram these existing (i.e. projected) species are represented by red discs. In the diagram these species are ordered as to the derivability of the wing-venation possessed by their representatives.
The uncolored discs, on the other hand, represent noëtic descriptions that are not projected into the Explicate Order, and do, consequently, not exist there as organismic species. In fact, in diagrams like the present one -- diagrams of morphoklines -- the uncolored discs represent hypothetical species allegedly making up the noëtic history of the existing species, that is to say, the vertical column of uncolored discs drawn beneath some given red disk makes up the series of alleged noëtic precursors of the latter. However, although the discs represent noëtic descriptions of (complete) strategies/species, the symbols such as B', B'', C '''', etc. placed at these uncolored discs do not refer to corresponding strategy/species, but to similar venations, that is, to a certain characteristic of such a species. So in the diagram above, the sign D' refers to the fact that the hypothetical species (represented by the respective uncolored disc) has a wing-venation similar to that of the true species D, while, to take another example, the sign C'' indicates the fact that the hypothetical species, represented by the uncolored disc accompanied by that sign, has a wing-venation similar to that of the true species C, but less similar than is that of D' to D.
The (true) species (represented in the explicate-order-part of the diagram by their genus name) Tricyphona, Molophilus, Orimarga, and Toxorhina, are in the diagram arranged as to the derivational order of their wing-venations. They constitute a derivational line with respect to wing-venation. And although the derivations are merely formal and at the same time being not strict derivations of the wing-venations of these species, they give us some clue as through what venational stages the species had passed during their noëtic history :
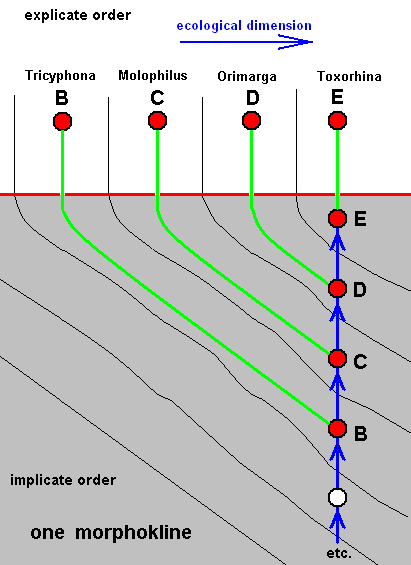
The four morphoklines of the (imaginary) chaetotaxy of the limoniids Molophilus, Orimarga, Toxorhina and Tricyphona (coded, respectively as C, D, E, and B ).
Here we have -- to take just a few examples -- the fact that species E noëtically directly stems from a hypothetical species -- D' -- of which we know (in fact suppose) that it has a chaetotaxy similar to the existing species D. And this is precisely what is expressed by the sign D', the similarity of the chaetotaxy with that of species D. In the same way the species B directly stems from the hypothetical species E' which is supposed to have a chaetotaxy similar to that of the existing species E. And also here the sign E' does not refer to the hypothetical species but to its chaetotaxy being similar to that of E.
So while the venational morphokline of species E was : A'''', B''', C '', D', E, the morphokline of chaetotaxy is : A''', C '', D', E.
And while the venational morphokline of species D was : A''', B'', C ', D, the morphokline of chaetotaxy is : A'', C ', D.
And, further, while the venational morphokline of species C was : A'', B', C, the morphokline of chaetotaxy is : A', C.
And, finally, while the venational morphokline of species B was : A', B, the morphokline of chaetotaxy is : A'''', C ''', D'', E', B.
So the chaetotaxic morphoklines (based on a chaetotaxic derivational line) are different from the venational morphoklines (based on a venational derivational line). The former tell us about the transformations of chaetotaxy in the line of noëtic precursors of each of the mentioned species, while the latter tell us about the transformations of the wing-venation in the line of noëtic precursors of each of these same species.
Accordingly, we may describe the hypothetical noëtic precursors of the species C, D, E, and B, either in terms of their chaetotaxy or in terms of their wing-venation (or in terms of some other organ or feature of these species). And these must be included when we describe these precursors in terms of species :
The in the Explicate Order existing species E noëtically directly stems from a certain hypothetical species that is supposed to have a wing-venation similar to that of existing species D (see venational diagram above) and a chaetotaxy also similar to that of existing species D (see chaetotaxy diagram).
The in the Explicate Order existing species B noëtically directly stems from a certain hypothetical species that has a wing-venation similar to that of existing species A (not explicitly given in the diagram) (see venational diagram above) and a chaetotaxy similar to existing species E (see chaetotaxy diagram). The signs A' (in the venational morphokline) and E' (in the chaetotaxy morphokline) ultimately refer to one and the same (hypothetical) species, where A' especially refers to its wing-venation and E' to its chaetotaxy. That is to say that each existing species has a linear noëtic ancestral line of (hypothetical) species (noëtic descriptions of strategies), extending into the Implicate Order. And each such a hypothetical ancestral species can be characterized by the state of certain of its organs or of other of its features. Here we have mentioned its chaetotaxy and its wing-venation. And of course such a hypothetical species is only completely characterized and defined when all its features are included in this way. So while chaetotaxy is used here as just a (largely imaginary) example of a feature other than wing-venation, all our diagrams depict morphoklines of wing-venation. And in them the uncolored discs, connected by blue arrows indicating their noëtic origin from one another, represent hypothetical species, while the signs associated with these discs refer only to their wing-venation.
The hypothetical species in our diagrams of morphoklines (morphoklines of venation, but the same holds for any other morphoklines) suggest a polyphyletic evolutionary development of organisms (in which occasionally small monophyletic groups may occur). But our view differs from conventional views in that this evolutionary development -- polyphyletic or otherwise -- takes place, not in the space-time world of the Explicate Order, but entirely in the Implicate Order. And there, "evolutionary development" is not directly a physical biological process, but a noëtic process of developing strategies for immaterial patterns to exist (as material patterns) in the Explicate Order, a process driven by the 'aspiration' of all immaterial patterns to become ontologically complete, that is, become material. Of course, here in the Implicate Order the mentioned "process of developing strategies" is not a process taking place in time and space, but a noëtic process, having the nature of formal derivations (often resulting from noëtic 'reactions'). And the mentioned immaterial patterns are in fact noëtic descriptions, and a "strategy" is a noëtic description prescribing how this strategy itself -- as an immaterial pattern -- becomes able to exist in the Explicate Order, where it then -- upon projection -- appears as a (new) organismic and material species.
So when we speak about the polyphyletic evolutionary development of organisms we mean their phylogeny in the Implicate Order. And, as has been said, cases of monophyly may exist, and the science of Phylogenetic Systematics (originally meant to reveal phylogenetic relationships between organisms in the Explicate Order) may be used by us to reveal monophyletic groups in the Implicate Order, i.e. reveal the content of such groups. However, conventional phylogenetic systematics considers polyphyly of species to be non-existent, and acknowledges only polyphyly of certain characters. So in phylogenetic systematics monophyly is the rule. But it must acknowledge the difficulties that exist in proving especially larger groups to be monophyletic. There are four causes of these difficulties : First, it is often hard to find in such a group reliably derived characters (apomorphic character states), derived that is, with respect to those of species not belonging to the group studied in this respect. Secondly, it is often hard to decide which state of a given character is derived and which state non-derived (plesiomorphic). Thirdly, a derived character must, in order to demonstrate the monophyly of the group, the species of which possess that character, be such that it can legitimately be asssumed that it has, in the larger group containing the (smaller) group studied, originated only once in some single ancestral species and was then inherited by all that species' descendants, descendants that is, who then form the monophyletic group (and the more such characters are found, the more certain is the monophyly of that group of organisms). It is, however, difficult to prove that the derived character (state) has this nature (of having it originated only once). That is to say, such a new character, or new character-state, must be some elaborate and uncommon construction or re-construction of which repeated and independent originations are hard to imagine. Nevertheless many such characters are known to have independently originated repeatedly in different organisms. And finally, the most troublesome difficulty for conventional phylogenetic systematics is the abundance of specialization-crossings in pairs of related species (or higher taxa), which forces phylogenetic systematics to push back further and further in time the evolutionary bifurcation-point where the species have allegedly separated from each other, that is, when one ancestral species splits up into two daughter-species. Indeed, already in wing-venations of related species of insects specialization-crossings abound.
And apart from the points mentioned, phylogenetic systematics relies on the neo-darwinian view of evolution with its problematic assumption that only random genetic mutations and natural selection form the driving force of organic evolution. According to this view every derived character (of an organism), or at least many of them, was the result of some random genetic mutation or series of them, subsequently preserved by natural selection.
Of the next derivational sequence (line) the first two members are the same as in the previous one :
LINE 2 :
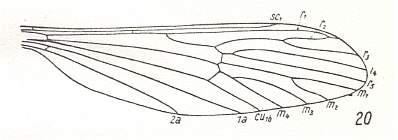
|
Tricyphona protea, Limoniidae. R2, R3, R4, and R5 present. M1, M2, M3, and M4 present. No intermedial cross-vein, and therefore no discoidal cell. The cross-veins r-m and m-cu present. CuA, 1A, and 2A present. |
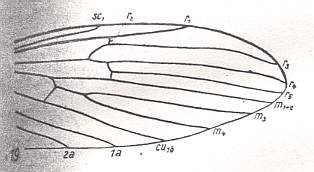
|
Molophilus flavoscutellatus, Limoniidae. R2 has shifted basad. It has become crossvein-like. M1 and M2 have coalesced. |
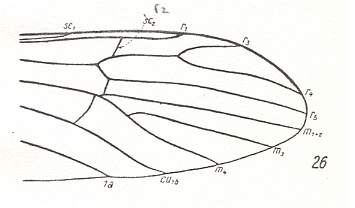
|
Empeda nubila, Limoniidae. The veinlet pointed to is R2 (not Sc2). R4 has 'jumped' onto R3. |
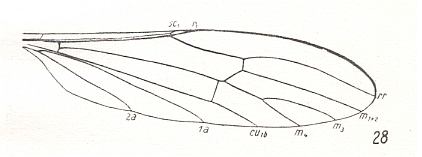
|
Toxorhina muliebris, Limoniidae. RS has become unbranched. R2, R3, and R4 have vanished. |
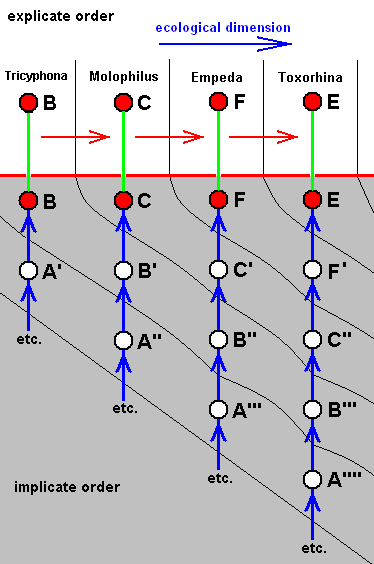
The four morphoklines of the venation of the limoniids Tricyphona, Molophilus, Empeda, and Toxorhina (coded, respectively as B, C, F, and E ). Blue arrows indicate noëtic derivations, and the red arrows indicate formal derivability of the venation in these genera. The fact that a given and legitimate derivational line (previous case) allows the replacement of one of its members to result in yet another legitimate derivational line, does not constitute a problem in our noëtic theory, because our diagrams simply express the fact that there is a morphokline of the venation in Orimarga as well as one of the venation in Empeda.
The next derivational line has again the first tree member being the same as in the previous lines :
LINE 3 :
|
|
Tricyphona protea, Limoniidae. R2, R3, R4, and R5 present. M1, M2, M3, and M4 present. No intermedial cross-vein, and therefore no discoidal cell. The cross-veins r-m and m-cu present. CuA, 1A, and 2A present. |
|
|
Molophilus flavoscutellatus, Limoniidae. R2 has shifted basad. It has become crossvein-like. M1 and M2 have coalesced. |
|
|
Empeda nubila, Limoniidae. The veinlet pointed to is R2 (not Sc2). R4 has 'jumped' onto R3. |
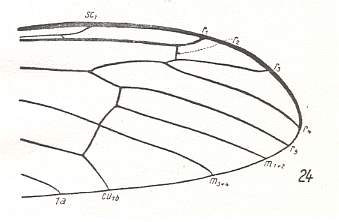
|
Hexatoma bicolor, Limoniidae. R2 has shifted onto the apical section of R3. M3 and M4 have coalized. |
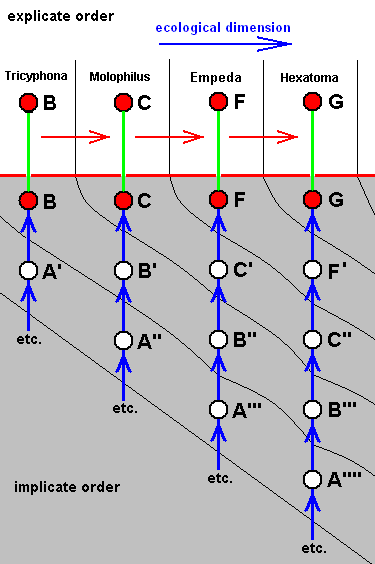
The four morphoklines of the venation of the limoniids Tricyphona, Molophilus, Empeda, and Hexatoma (coded, respectively as B, C, F, and G ). Blue arrows indicate noëtic derivations, and the red arrows indicate formal derivability of the venation in these genera. Also here the fact that a given and legitimate derivational line (previous case) allows the replacement of one of its members to result in yet another legitimate derivational line, does not constitute a problem in our noëtic theory, because our diagrams simply express the fact that there is a morphokline of the venation in Toxorhina as well as one of the venation in Hexatoma.
Limoniidae with discoidal cell.
LINE 4 :
The four morphoklines of the venation of the limoniids Limnophila, Dicranoptycha, Antocha, and Orimarga (coded, respectively as I, J, H, and D ). Blue arrows indicate noëtic derivations, and the red arrows indicate formal derivability of the venation in these genera. Here we see that the venation of Orimarga may figure in two different morphoklines (for the other one, see the above diagram ) :
One of them is the present one : ... A'''', I''', J'', H', D and the other is ... A''', B'', C', D. And it might well be that, at least qua venation, C' = H', B'' = J'', and A''' = I''' (After all, C', H', B'', J'', A''' and I''' are just hypothetical venational links).
LINE 5 :
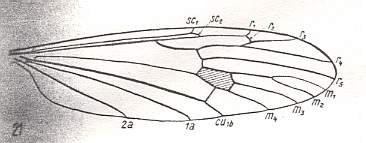 Of the limoniid wings with a discoidal cell this wing is the most primitive. |
Limnophila ferruginea, Limoniidae. The shaded region (that is, the region drawn shadedly) is the discoidal cell. Fork of R2+3 shifted onto R4. R2 ending up in R1. M1, M2, M3, and M4 all present. CuA, 1A, and 2A present. |
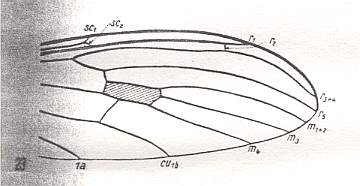
|
Dicranoptycha livescens, Limoniidae. R3 and R4 coalesced. M1 and M2 coalesced. |
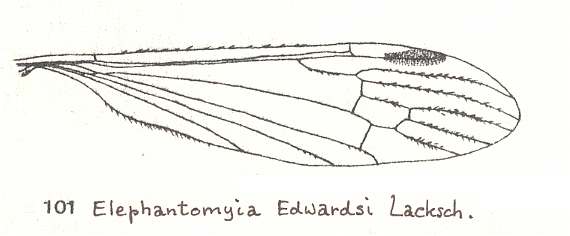
|
Elephantomyia edwardsi, Limoniidae. R2 absent. Main junctions of the veins shifted towards the wing-apex. |
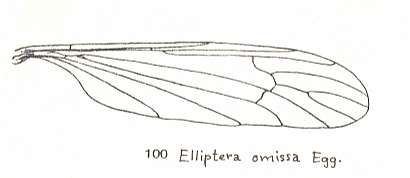
|
Elliptera omissa, Limoniidae. The cross-vein tp (intermedial cross vein between M2 and M3) vanished. Therefore, discoidal cell absent. |
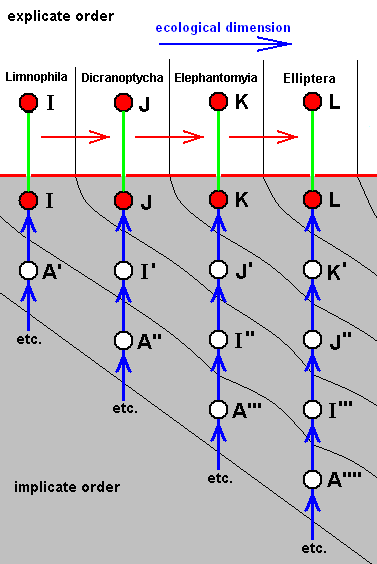
The four morphoklines of the venation of the limoniids Limnophila, Dicranoptycha, Elephantomyia, and Elliptera (coded, respectively as I, J, K, and L ). Blue arrows indicate noëtic derivations, and the red arrows indicate formal derivability of the venation in these genera.
LINE 6 :
|
Of the limoniid wings with a discoidal cell this wing is the most primitive. |
Limnophila ferruginea, Limoniidae. The shaded region (that is, the region drawn shadedly) is the discoidal cell. Fork of R2+3 shifted onto R4. R2 ending up in R1. M1, M2, M3, and M4 all present. CuA, 1A, and 2A present. |
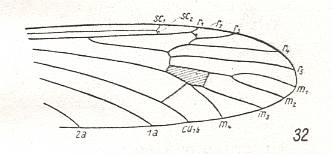
|
Architipula clara, Limoniidae. Upper Liassic (= upper lower Jurassic) of Mecklenburg (Germany). Virtually the same venation as in (the recent) Limnophila, despite its age. |
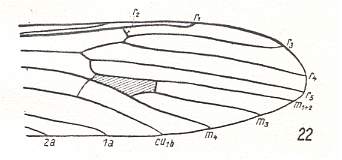
|
Erioptera trivialis, Limoniidae. Fork formed by R2 and R3 seemingly shifted basad. This is not a straightforward derivation of this fork from that of the previous forms (Architipula, Limnophila), but merely a somewhat different configuration and position of its elements. M1 and M2 coalesced. |
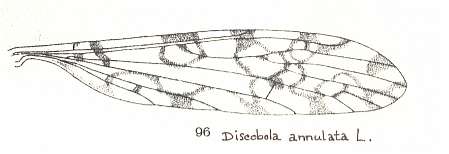
|
Discobola annulata, Limoniidae. R3 absent. |
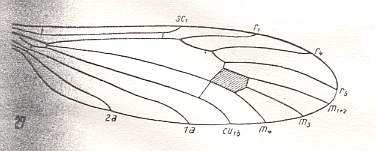
|
Atarba picticornis, Limoniidae. R2 (and also R3) absent, leaving the Radial Sector 2-branched. |
|
|
Elliptera omissa, Limoniidae. The cross-vein tp (intermedial cross vein between M2 and M3) vanished. Therefore, discoidal cell absent. Main junctions of the veins shifted still further towards the wing-apex (except the base of RS). |
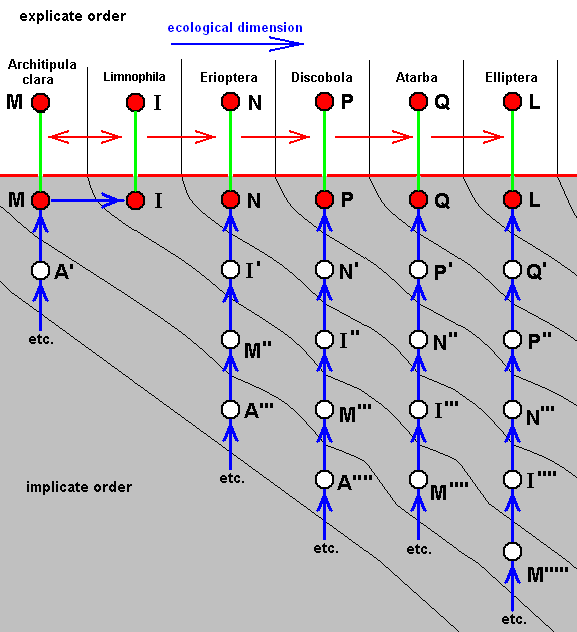
The six morphoklines of the venation of the limoniids Architipula clara, Limnophila, Erioptera, Discobola, Atarba and Elliptera (coded, respectively as M, I, N, P, Q and L ). Blue arrows indicate noëtic derivations, and the red arrows indicate formal derivability of the venation in these genera. The morphokline (= noëtic history) of Architipula clara is : M, A', ..., while that of Limnophila is : I, M, A', ....
One might think that the positions of A. clara and noëtic ancestors might be drawn in a more simple and straightforward way, such as in the next diagram :
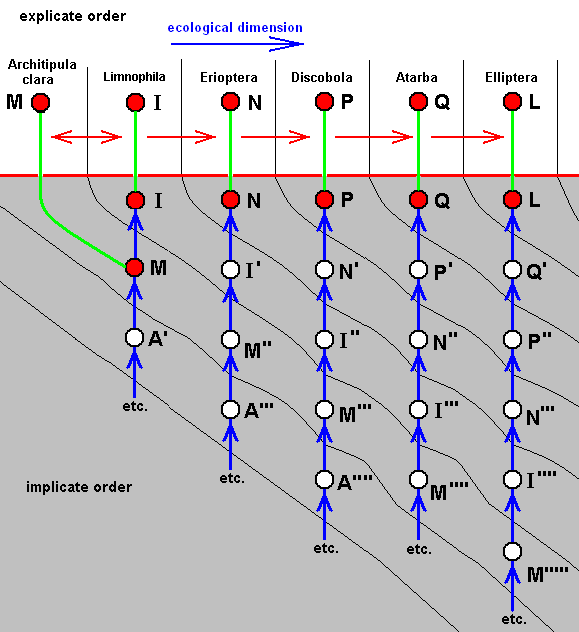
Same as previous diagram, but with A. clara, and noëtic ancestors -- in the implicate order part -- displaced, resulting in the morphokline I, M, A', ... to be drawn as a straight line like all the other morphoklines. And so we might think that this version of the original diagram is fully equivalent to the latter (because the noëtic histories are the same in each). But this isn't so : As soon as we have decided on precisely where in the (implicate order part of the) diagram to place M, I, A' (and all others), that is, have decided on precisely in what noëtic neighbourhood -- in the Implicate Order -- these strategies should reside, we cannot change that anymore. So each one of the two versions is in itself correct, and as such equivalent, but they cannot be used together at the same time, because the one does not imply the other. But as long as we can still choose, we prefer the second version.
Strictly, here in this diagram, M (in noëtic space, i.e. in the implicate order part of the diagram) should in fact be M', and A' should be A'' according to their precise position in noëtic space, and Architipula clara would then be denoted by M'.
However, despite the fact of their wing-venations being almost identical, Limnophila and Architipula clara could nevertheless lie on different morphoklines. In that case the identical wing-venation is an example of convergence (same product, different origins) :
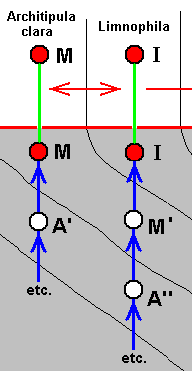
Generally, in the explicate order part of diagrams like the above three ones -- in which fossil species occur -- it would be natural when we placed the species in geologic order (that is the order of their appearance in the Explicate Order). But in fact the type of diagram we're now considering is not able to express order of appearance of species, i.e. expressing it such that it is fully integrated with the rest of the diagram. Therefore in our diagrams the consecutivity of the species entered in the diagram does not reflect the order of appearance of these species. And in this part of the diagram, i.e. in the explicate order part, we will indicate the derivational relationships of the venation of these species by red arrows. And only when these venations can strictly be derived from one another and in addition belong to the same genus we may assume that not only the venations, but also the corresponding species are -- in the Implicate Order -- derived from one another. Like all such true derivations we indicate them -- in the implicate order part of the diagram -- with blue arrows. In all cases where the species are not strictly derivable from one another, they, together with their (hypothetical) noëtic precursors, will be placed in separate morphoklines. And also here blue arrows indicate true derivations, but now among hypothetical precursors. In fact, the formal derivational relations that are found to exist between the wing-venations encountered in a given group of insects, and indicated by red arrows in the diagrams, but also the derivational relations existing between structures other than wing-venation (including, for instance, the structure of the thorax of the adult insect, its mouth-parts, its antennae, bristle patterns, etc, and also the morphological structures of the larvae -- mouth-parts, bristles, integument, etc.), give us a clue as to which morphological stages each species of the group went through during its phylogenetic development. These stages represent the line of ancestral species of each species of the group. But these existing derivational relations just provide us with a clue as to how these stages might have looked like, and nothing more. So each morphokline (in our case based on wing-venation, but representing species) of an existing species of a given group of insects consists of a series of ancestral species that more or less reflect the uncovered derivational relations as found in the recent (or fossil) representatives of this group.
As can be seen from the above 6 derivational lines, the derivational relationships between limoniid wing-venation display a criss-cross nature, but in all this variation the wing-venation remains within the boundaries of the typical overall structure.
The next derivational line also starts with wings of Limoniidae, but mainly includes wings of the families Tipulidae and Cylindrotomidae.
Line 7 :
In derivational line 7 we see a gradual transformation of the course and configuration of the veins R1, R2, R3, and R4 (collectively calling it the system "(R1 - R2 - R3 - R4)". The line begins with this system as seen in Limoniidae, passes over into that seen in Tipulidae, and concluding with it as it is seen in Cylindrotomidae.
|
Of the limoniid wings without a discoidal cell this wing is the most primitive. |
Tricyphona protea, Limoniidae. Fork of R2+3 independent of the next vein, i.e. not yet shifted onto R4. R2, although long (i.e. of normal proportions) and not recurrent, ends up in R1. M1, M2, M3, and M4 all present. The cross-vein tp (= intermedial cross-vein) absent and therefore there is no discoidal cell. CuA, 1A, and 2A present. With respect to the venational transformation of the system (R1 - R2 - R3 - R4) the present form (Tricyphona) is the starting condition. |
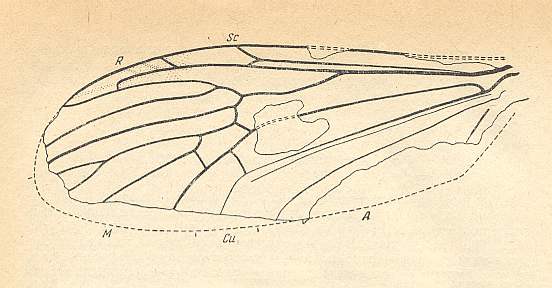 |
Architipula radiata, Limoniidae. Lower Jurassic of Issic Kul, central Asia. Length of wing 5.9 mm. Fork of R2+3 shifted onto R4. It has a long stalk which branches off from the basal section of R4. Especially R2 is long, and recurrent, ending up in R1. R3 ends up fairly close to the wing-apex. M1, M2, M3, and M4 all present. Discoidal cell present. CuA (and also CuP), 1A, and 2A present. The structure of the complex (R1 - R2 - R3 - R4) is similar to that in the next wings of this line, but not necessarily structurally preceding them in this respect. |
|
Of the limoniid wings with a discoidal cell this wing (and the next) is the most primitive. |
Architipula clara, Limoniidae. Upper Liassic [= upper lower Jurrassic] of Mecklenburg (Germany). The shaded region (that is, the region drawn shadedly) is the discoidal cell. Fork of R2+3 shifted onto R4. R2, being short, crossvein-like, ending up in R1. M1, M2, M3, and M4 all present. CuA, 1A, and 2A present. |
|
Of the limoniid wings with a discoidal cell this wing is the most primitive. |
Limnophila ferruginea, Limoniidae. Fork of R2+3 shifted onto R4. R2 ending up in R1. M1, M2, M3, and M4 all present. CuA, 1A, and 2A present. |
|
|
Pales dorsalis, Tipulidae. R2 stronger recurrent. End of R3 approaches that of R1. |
|
|
Phalacrocera replicata, Cylindrotomidae. R2 still stronger recurrent. R3 having the same position as in Pales. M3 absent. M4 has taken its place. The previous and next wing each have all the four medial branches, therefore the present form cannot derivationally mediate between them. But it can do so with respect to the (R1 - R2 - R3 - R4) system. |
|
|
Tanypremna spec., Tipulidae. R2 and R3 (together still branching off from R4) still further transformed : R2 short and recurrent, and R3 very short and ending up almost at the endpoint of R1. R5 still present. M1, M2, M3, and M4 present. |
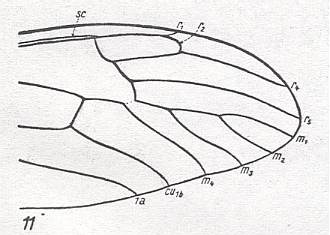
|
Dolichopeza albipes, Tipulidae. R2 recurent, ending up in R1. R3 vanished. R5 (and, of course R4) still present. M1, M2, M3, and M4 present, but the intermedial cross-vein tp missing. Therefore this form has no discoidal cell. |
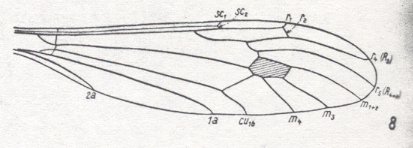
|
Idiotipula confluens, Tipulidae. This is also a form with R2 (moderately) recurrent and ending up in R1, and R3 absent, while R4 and R5 being still present, - but with discoidal cell. M1 and M2 coalesced. |
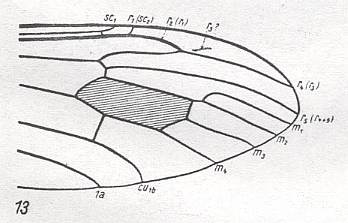
|
Cylindrotoma distinctissima, Cylindrotomidae. This form is in fact neatly following upon Tanypremna which is a little higher up in the present line. R1 still ends up in the wing-margin. R2 recurrent and ending up in R1. Maybe there is left a remnant of R3. M1, M2, M3, and M4 present. |
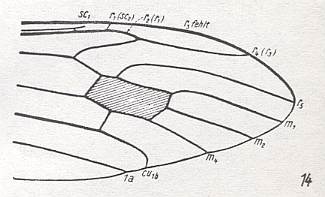
|
Triogma trisulcata, Cylindrotomidae. R2 recurrent, ending up in R1. R3 absent. The cross-vein ta (= r-m) is suppressed. M3 absent. |
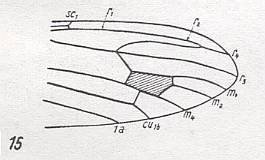
|
Stibadocerodes tasmaniensis, Cylindrotomidae. R1 not ending up in the wing-margin anymore. R2 + R2+3 is now the prolongation of R1. R3 absent. R5 still present as independent vein. M3 absent (as in Triogma). Discoidal cell still present. This is an end-point of the transformation of the (R1 - R2 - R3 - R4) system. |
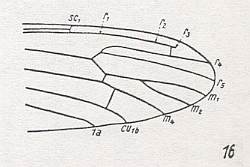
|
Stibadocerodes australiensis, Cylindrotomidae. Like Stibadocerodes tasmaniensis the present form can be more or less derived from Triogma : R4 and R5 still present as independent (from each other) veins, but the discoidal cell has vanished. The very short vein stump indicated as "r3" may be a new formation (new sclerotization in the old bed of R3) and thus not a remnant of R3. This is another end-point of the transformation of the (R1 - R2 - R3 - R4) system. |
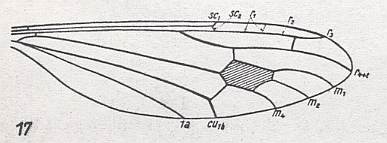
|
Stibadocerella albitarsis, Cylindrotomidae. This form may be derived from Phalacrocera (a little up in the present line). R2 recurrent, ending up in R1, which itself ends up in the wing-margin like in phalacrocera. R3 still present. R4 and R5 coalesced, forming one single vein. Like in Phalacrocera M3 is missing, and the discoidal cell is present. This is yet another end-point of the transformation of the system (R1 - R2 - R3 - R4) but now having involved R5. |
In the corresponding interpretation of this derivational line (involving wings of Limoniidae, Tipulidae and Cylindrotomidae) in terms of morphoklines we again use the letters A, B, C, D, ... for the genera (and species). There can be no confusion with genera in other diagrams, because in all diagrams the genus names (with their corresponding letters) are explicitly given. And, for reasons of convenience, the number of ' signs, as in B'''', is replaced by numbers ( B4).
Click HERE (and click on image) to see the diagram of morphoklines of this derivational line (7).
In the diagram we have the venation of Phalacrocera formally derived from that of Pales. But, as in virtually all such derivations (and certainly when we let involve all characters of the respective species), there are specialization-crossings present, that forbid strict derivation : In Pales the Medial system is primitive (as compared to Phalacrocera), whereas the place of origin of the Radial Sector is derived (as compared to Phalacrocera). Further, we must note that, although the venation of Tricyphona is indeed very primitive (and therefore having been placed at the beginning of Line 7), it has lost the discoidal cell. All this again reminds us of the fact that a derivational line of wing-venations does not represent their actual origination one from the other (and let alone that of the species possessing these venations), but only gives some indications through which states the venation of each species might have gone in its noëtic history.
As we saw in the derivational lines 1-6, in the Limoniidae the fate of the (R1 - R2 - R3 - R4) system is different from that in the Tipulidae-Cylindrotomidae (line 7). This is remarkable because, as in line 7, also the limoniid lines (1-6) start with Tricyphona (or with Limnophila having a discoidal cell).
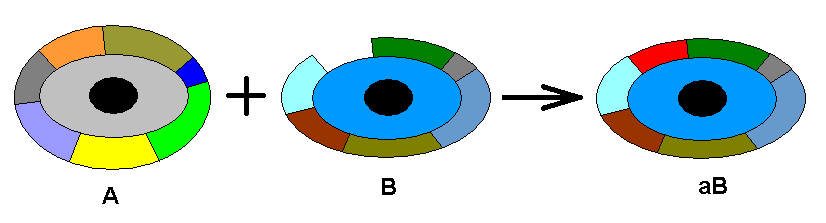
Figure 1 : Diagram of a noëtic reaction of the qualitative content of a species B with that of a (noëtically) closely related species A. In this reaction the qualitative content of the species B is completed by taking up elements (while at the same time changing them) from the qualitative content of species A.
Every species has its own strategy sensu stricto (represented by the two innermost areas in the diagrams). And every species has its own set of peripheral contents not belonging to the strategy sensu stricto.
Let us elaborate on this diagram.
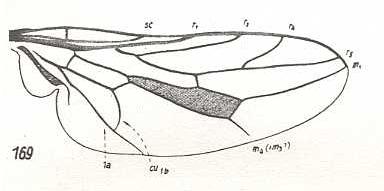
|
Omphrale fenestralis, Omphralidae. R4 and R5 curved upwards. M2 and M3 absent. M4 originating from discoidal cell, not reaching wing-margin. CuA and 1A with long common end-stalk. |
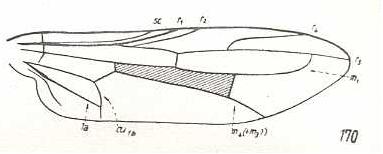
|
Pseudatrichia chilensis, Omphralidae. M1 ending up in R5 well before apex. M2 and M3 absent. M4 originating from discoidal cell, reaching wing-margin. CuA and 1A with shorter common end-stalk. Wing elongate. |
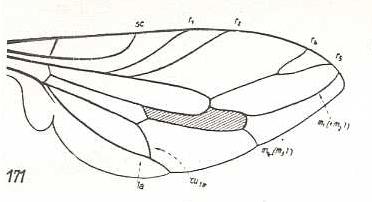
|
Apiocera maritima, Apioceridae. SC, R1, and R2, strongly curved upwards. Fork of R4+5 very short. M1 ending just above wing-apex. M2 and M3 absent. M4 originating from discoidal cell, reaching wing-margin. CuA and 1A with a moderately long end-stalk. Wing broadened. With respect to Pseudatrichia there exists a specialization-crossing : Apiocera represents a derived condition with respect to the curvings of SC, R1, and R2, while Pseudatrichia is in a derived condition as a result of the course of M1. |
We may assume that in noëtic reactions between qualitative contents of different species (existing in noëtic space [Implicate Order] as noëtic descriptions [the meaning of which appears in the Explicate Order upon projection] ) the adaptive and species-specific elements form a little- or non-reactive core in each such a qualitative content. In fact this core IS the qualitative content of the species. This results in the fact that these contents will not be affected by any reaction between them. Only the fact that in the qualitative content of a given species (in the Implicate order) one or more non-adaptive elements are not yet present renders this content to be still reactive (Any affection of the qualitative content of a given species, whether as a result of noëtic reactions with that of other species, or as a result of some other cause, may break down the strategy and seclude it from projection into the Explicate Order. Therefore, even if such reactions occur, we do not deal with them).
The next derivational line is about the family Psychodidae (moth-flies, sand-flies) but should begin with the venation of the primitive limoniid Tricyphona protea :
LINE 8 :
|
|
Tricyphona protea, Limoniidae. R2, R3, R4, and R5 present. M1, M2, M3, and M4 present. No intermedial cross-vein, and therefore no discoidal cell. The cross-veins r-m and m-cu present. CuA, 1A, and 2A present. So this venation is almost identical to the order-prototype. |
|
|
Nemopalpus zelandiae, Psychodidae. The venation of this wing can be derived from a venation that is very close to that of Tricyphona (Limoniidae). SC shortened but 2-branched. R2 ending up, not in R1, but at the wing-margin. Fork of R2+3, as in Tricyphona, branches off from the trunk of RS (which is the primitive condition of it). As in Tricyphona, M1, M2, M3, and M4 present, and no discoidal cell. CuA and 1A drastically shortened. 2A absent. So especially in the posterior part of the wing we see a shift of main bifurcations towards the wing-base. |
|
|
Phlebotomus papatasii, Psychodidae. SC still further shortened and reduced. Main bifurcations still further shifted basad. Fork of R2+3 branches off from R4. CuA touching, and partly coalescing with M4, and shortened still more. Anal veins absent. The apex of the wing has become pointed. |
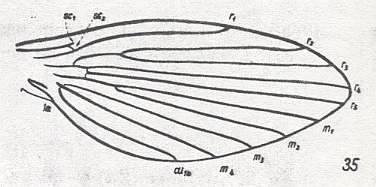
|
Ulomyia fuliginosa, Psychodidae. Shift toward the wing-base of the main bifurcations now also involving R2 and R3, and M1 and M2. Fork of R2+3 branching off from the trunk of RS again (which makes it difficult to derive this form from Phlebotomus. But it is hard to derive it from Nemopalpus too, because of the latter's short CuA ). The wing is significantly broadened, and its apex is pointed. CuA is long, but anal veins vestigial. |
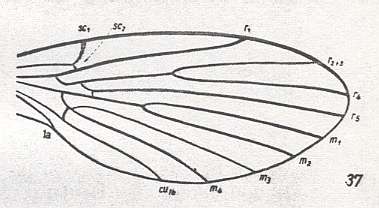
|
Trichomyia urbica, Psychodidae. Contrary to the labeling of the veins in the drawing I believe that in this form R4 and R5 have coalesced, and R2 and R3 still present. CuA still long, and 1A short. 2A absent. The wing is still broad, but the apex less pointed. If the labeling is correct, then this form can be derived from Phlebotomus : The fork of R2+3 is still branching off from R4. R2 and R3 have completely coalesced, forming one single R2+3 ending up at the wing-margin. As to the condition of CuA, it (Trichomyia) cannot be derived from Phlebotomus. |
|
Psychoda spec., Psychodidae. If the labeling of the veins in the drawing is correct, then M4 has vanished and 1A is still a long vein (as in Tricyphona [Limoniidae] ). If, on the other hand, we maintain that M4 is still present, then CuA is still long, but 1A is vestigial. 2A absent. The wing is still broad, and the apex is pointed again. The shift of the main bifurcations toward the base in this wing has reached its very limits. |
Here, in this line, we have seen clearly that the derivations are not strict derivations. The line nevertheless shows derivational relationships between the six venations.
The corresponding diagram of morphoklines of this Line 8 is the following :
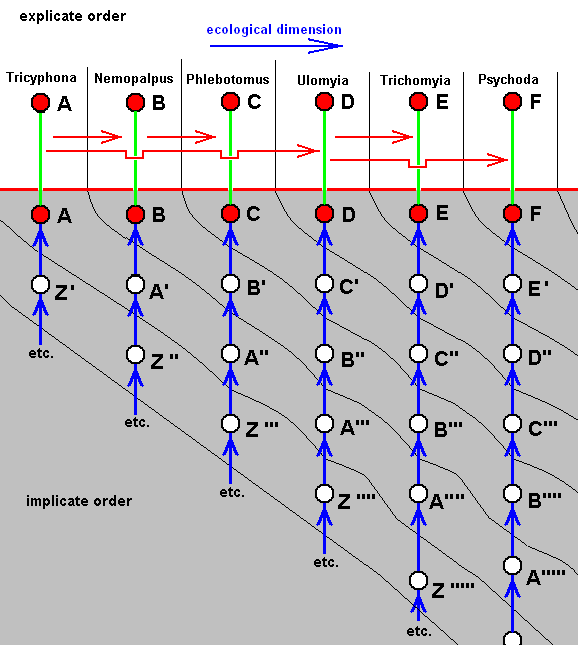
Also here we must realize that the derivational relationships between the above wing-venations -- (as in all our considerations) not the individual material objects, but the patterns they represent -- do not represent true genealogical relationships between these venations (and their possessors), that is, they have not necessarily originated one from the other, but show us the stages through which the wing-venation might have passed in the noëtic history of each of the species, as indicated by the corresponding lettering of these stages in the implicate-order-part of the diagram.
The next derivational line is about the family Tendipedidae (midges) (= Chironomidae), but should begin with the venation of a mosquito -- Anopheles (or Culex for that matter) (Culicidae). (Our interpretation of the venation does not necessarily comply with the naming of the veins -- by Hennig -- in the drawings) :
LINE 9 :
|
|
Anopheles maculipennis, Culicidae. R2 and R3 present. R4 and R5 completely coalesced (or : R2+3 branching off from R4. R2 and R3 completely coalesced. R5 present). R1 long, ending up near the wing-apex. M1, M2, and M4 present. M3 absent. No intermedial cross-vein, and therefore no discoidal cell. The cross-veins r-m and m-cu present, the latter long and curved. CuA and 1A both long, and 2A vestigial. |
|
|
Trichotanypus nigriventris, Tendipedidae. SC reducing. R1 shortened, ending up at the anterior wing-margin. R2 and R3 shortened, R2 ending up in R1. R3 ending up in the front margin of the wing, as well as the next vein consisting of the coalesced veins R4 and R5. M1 and M2 completely coalesced. The vein 1A strong but not reaching the wing-margin. The vein 2A present (apparently weakly so). |
|
|
Pelopia spec., Tendipedidae. Venation largely the same as that of Trichotanypus. The vein 1A weak. (CuP drawn here, being largely a mere fold [not drawn in many other figures] ). The vein, in the drawing signified as An1, may be 2A. In that case 1A absent. |
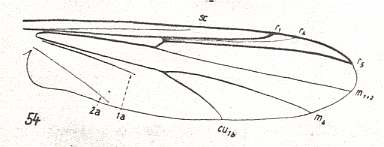
|
Polypedilum nebeculosum, Tendipedidae. R2 disappeared. As regards 1A this form is best derived from Trichotanypus . |
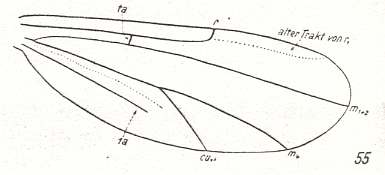
|
Thienemanniella spec., Tendipedidae. R3 and the coalesced veins R4 and R5 absent, that is, the complete Radial Sector has vanished. 2A absent. |
The corresponding diagram of morphoklines of this derivational Line 9 is the following ( Again, the use of the same letters will not cause confusion with other genera [in other diagrams], because these are explicitly given. Also the use of numbers instead of accents will not confuse us) :
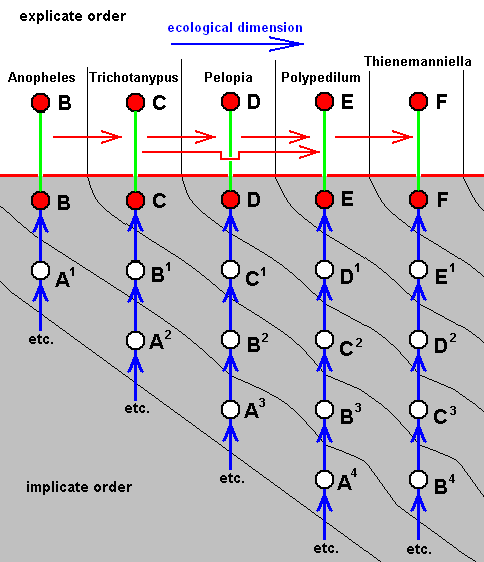
The next derivational line is about the family Heleidae (biting midges) (= Ceratopogonidae), but should begin with the venation of a representative of the family Thaumaleidae, namely Thaumalea major :
LINE 10 :
The corresponding diagram of morphoklines of this derivational Line 10 is the following ( Again (and for always) the use of the same letters will not cause confusion with other genera [in other diagrams], because these are explicitly given. Also the use of numbers instead of accents will not confuse us) :
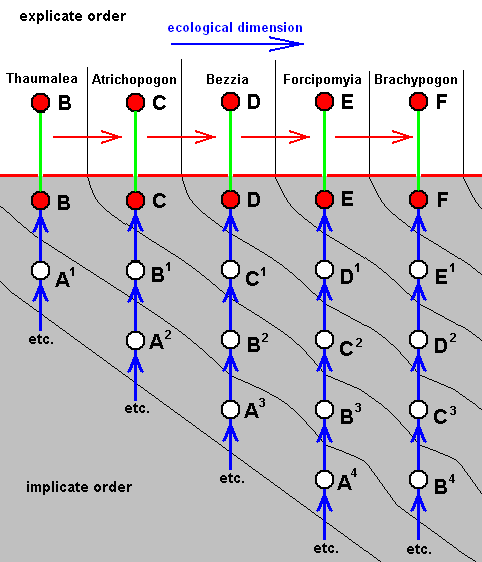
The next derivational line is about the family Blephariceridae. Its groundplan-venation cannot neatly be derived from venations of other families.
LINE 11 :
Also in this line we see that the derivations are not strict derivations of everytime one whole venation from another.
And, further, the Blephariceridae still await their evaluation as to what functional type their wings belong.
The corresponding diagram of morphoklines of this derivational Line 11 is the following :
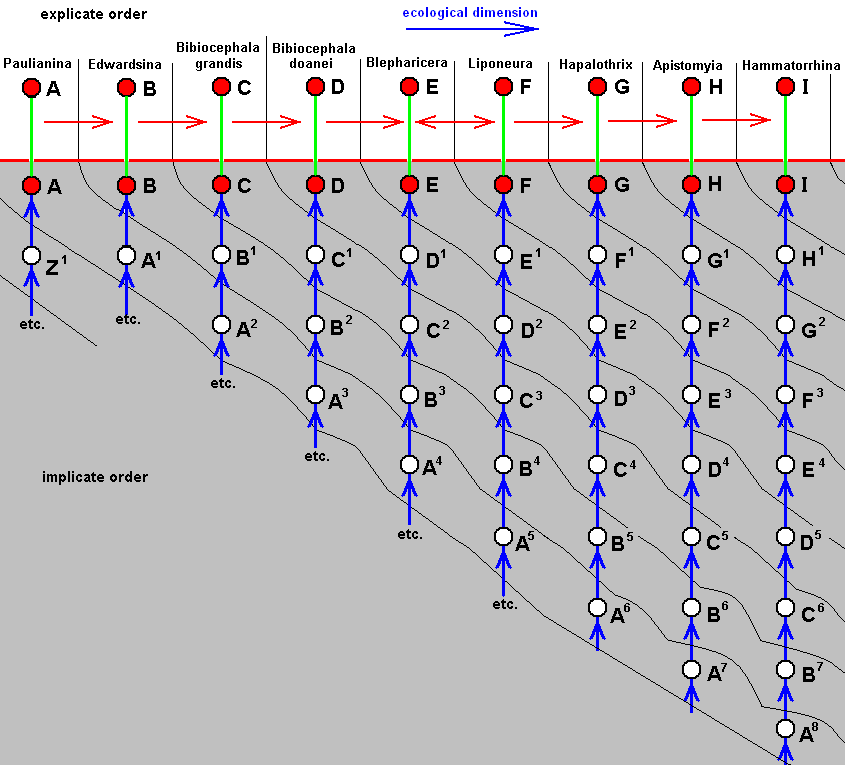
Bibionomorpha
Essentially, derivational lines of the wing-venation in the families of the Infraorder Bibionomorpha (many of them having wings belonging to the fungivoroid and bibionoid functional types) have already been discussed (and extensively so indeed!) in Part XXVIII and Part XXVIII-A . These derivational lines (= wing-venations ordered according to their successive derivability one from the other) were used to establish the phylogenetic system of the Bibionomorpha, that is, to find monophyletic groups in them and determine their phylogenetic relationships. In the present documents, however, we are investigating the possibility of polyphyletic evolution of organisms including insects. And we do this by deriving venational morphoklines from derivational lines (where a given venational derivational line of existing species tells us something about the venational transformations having taken place in the noëtic history of these species). We shall derive such venational morphoklines in Bibionomorpha in Part LXXa after we have dealt with (in fact reproduced from earlier documents) the functional wing-types that can be found in the Bibionomorpha in ( Part LXIX and Part LXX ).
In the next document (Part LXX) we continue, and conclude, our exposition of the functional wing-types in Diptera-Nematocera (while from Part LXXI onwards we expound the functional wing types in Diptera-Brachycera (orthorrapha and cyclorrapha)), and the derivational lines (and morphoklines) of the venations involved in these types).
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of Organic Evolution, Part LXX.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII
Back to Evolutionary Part LXII
Back to Evolutionary Part LXIII
Back to Evolutionary Part LXIV
Back to Evolutionary Part LXVI
Back to Evolutionary Part LXVII
Back to Evolutionary Part LXVIII
Back to Evolutionary Part LXIX
Back to Evolutionary Part LXIXa
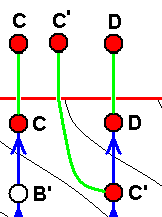
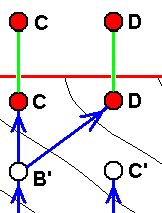
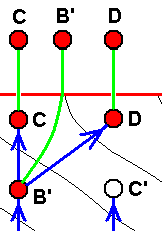
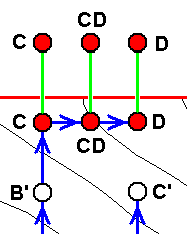
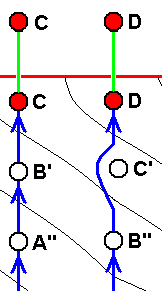
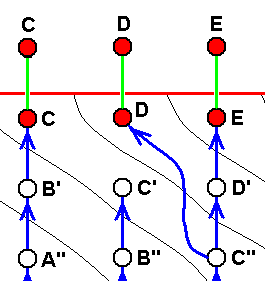
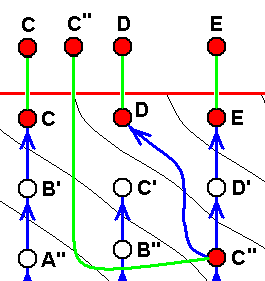
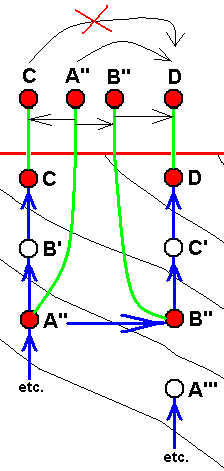

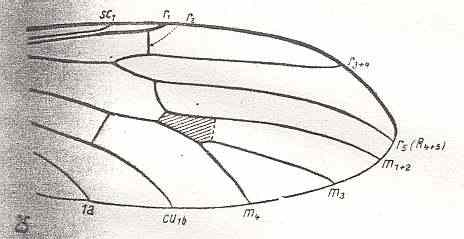
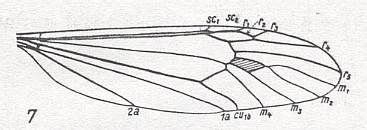
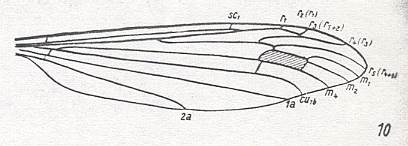
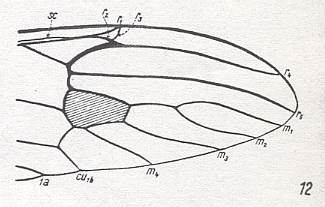
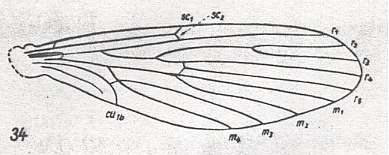
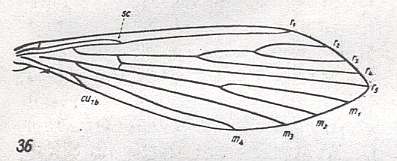
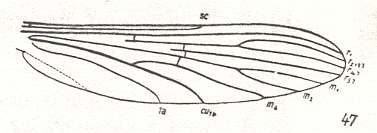
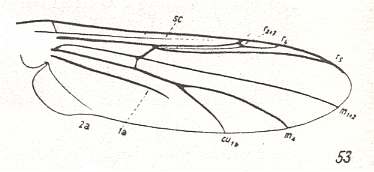
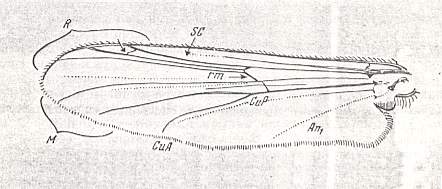
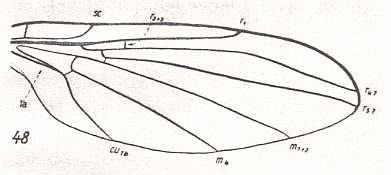
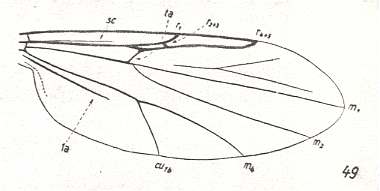
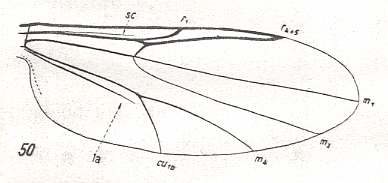
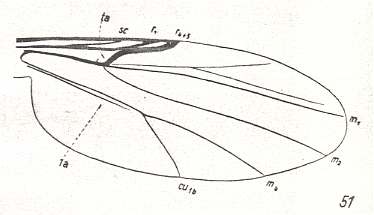
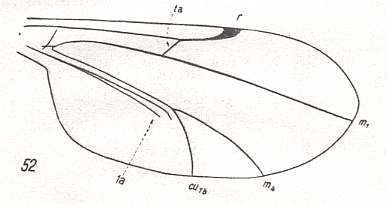
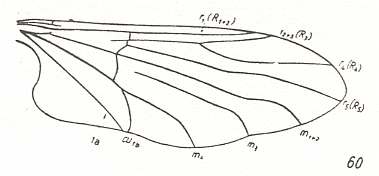
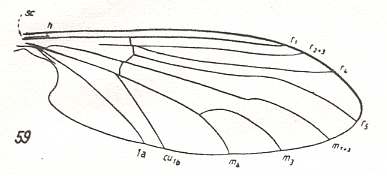
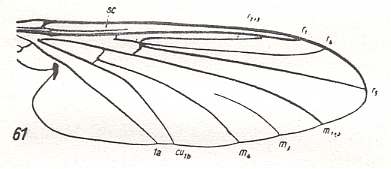
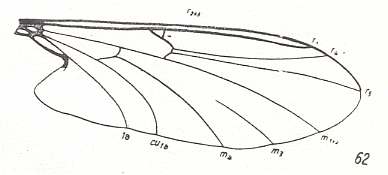
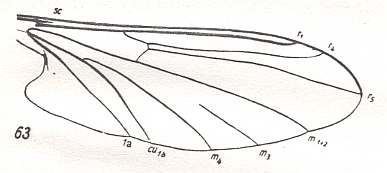
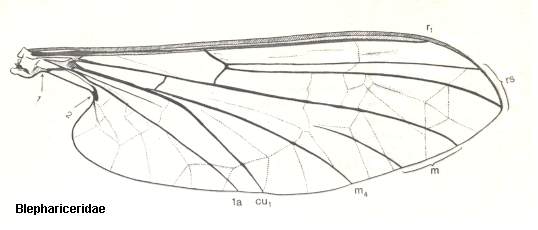
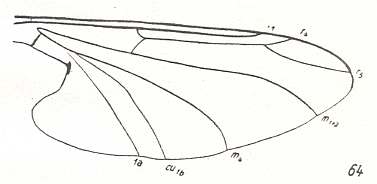
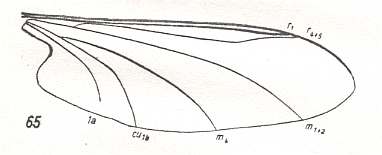
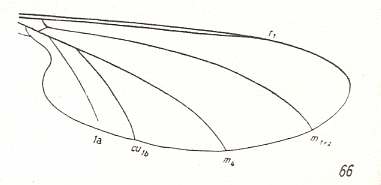
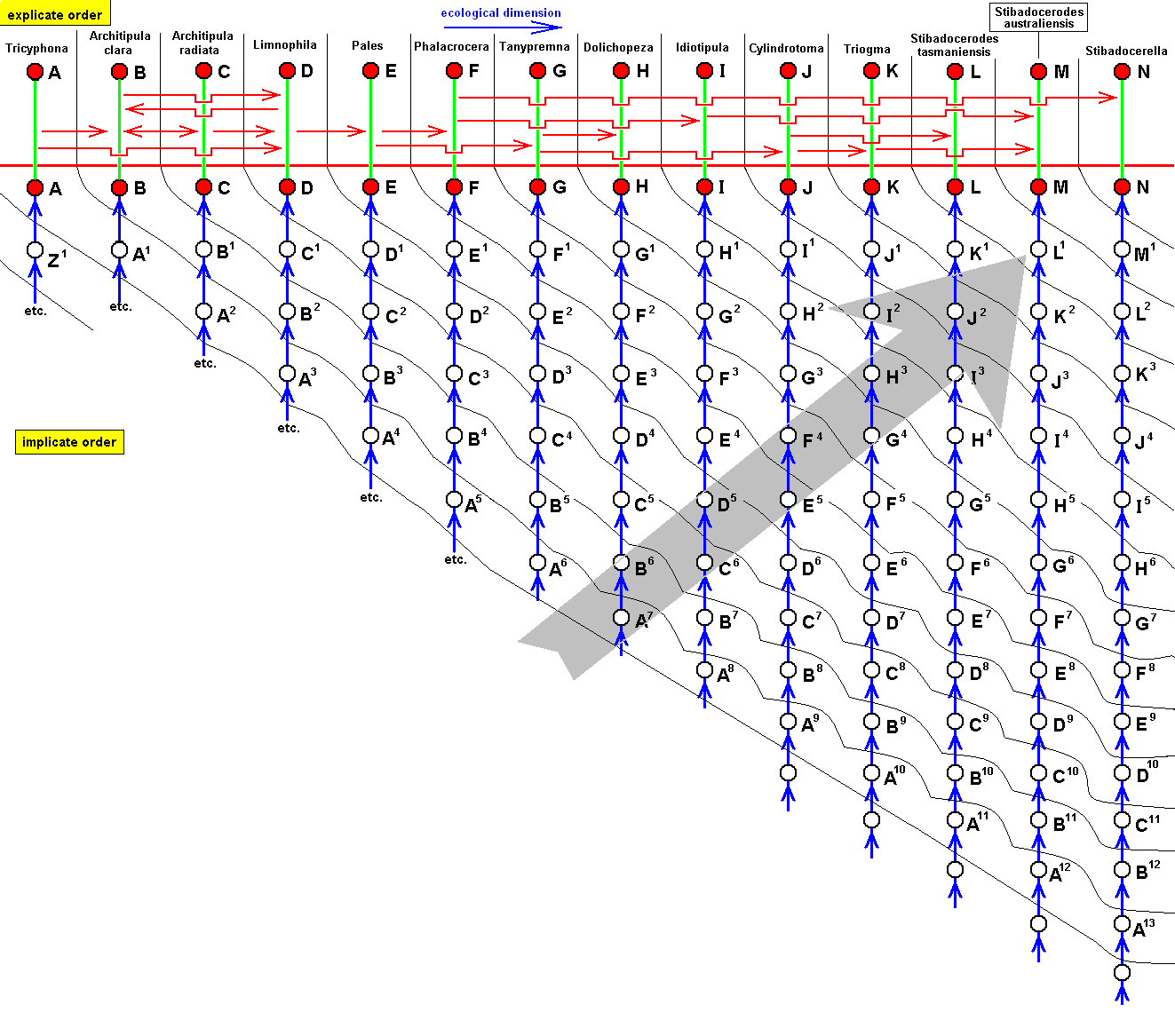{kind=link}