Origin and development of Organic Strategies in the Implicate Order, and their appearance in the Explicate Order.
Our theory is, we must admit, complex and somewhat confusing, not only for the esteemed reader but also for the author himself. This is because the problem --- to describe the principles of organic evolution, and remaining thereby sincere, and not taking refuge behind theological 'solutions', which are no solutions at all but merely increase the degree of miraculousness still more --- is so immensely complex and so difficult to solve if that is possible at all. Our theory contains elements that have been later refined or changed without removing what was established earlier. This is because the theory is in full swing of development, and it is also this very development which we want to describe. And, moreover, ealier, later revised, results, may in the future turn out to be valuable again. So this is the reason why we retain earlier results even when they have been revised later on, and despite the confusion this retention may cause.
What follows still belongs to our summary of the present state of theory, but even in this summary some things need to be summarized again in order not to loose track in this tangled theory.
We are still trying to work out how organic strategies of how to exist in the Explicate Order are actually constructed in the Implicate Order. In order for strategies to be so constructed, the Implicate Order must somehow be itself geared to the Explicate Order, because the construction of organic strategies in the Implicate Order cannot take place in complete isolation from the Explicate Order : The strategies, as they finally appear in the latter Order in the form of organismic species (material strategies), turn out to be very 'sophisticated', 'devised', 'designed', 'thought up', and 'invented' indeed. And this was one of the first reasons to question the Neodarwinistic explanation of organic evolution. But in devising an alternative general theory of evolution, we insist upon dismissing two general views, which might figure in such a theory :
All noëtic patterns, existing in the Implicate Order, which are themselves mere Forms (that is, beings without matter) possess an inherent tendency toward ontological completion (by in-forming Prime Matter, becoming individual, material, etc.), that is, toward existence in the Explicate Order, which existence is accomplished by the 'projection' of such a pattern from the Implicate Order into the Explicate Order. As a result of this ontological tendency all noëtic patterns are predisposed, not necessarily to exist as such in the Explicate Order, but at least predisposed to explicate existence as such, that is, they at least 'are about' explicate existence.
For some patterns this predisposition goes more specific : They either are, just like that, such that they directly project (or 'are projected') into the Explicate Order, or, if not, noëtically develop into strategies-to-exist-in-the-Explicate-Order. And because they are all generally predisposed, i.e. geared to, the Explicate Order, the mentioned development in the Implicate Order of these noëtic patterns into organic strategies (and thus into other patterns) is not a mere chance event, but a constant and inherent tendency. And, as we know, such strategies appear in the Explicate Order as organismic species (material strategies).
But the noëtic development, and thus construction, of strategies is not as such the noëtic counterpart of organic evolution, because evolution is the successive appearance of 'ready-made' strategies in the Explicate Order. Subsequent to their production in the Implicate Order the strategies together constitute an order of formal derivability, a formal system. And only upon projection (in reversed order) into the Explicate Order they constitute organic evolution, implying that the formal system of noëtic strategies (strategy-strings) becomes 'interpreted' : Each element, strategy, obtains meaning, that is, the significatum of a given noëtic strategy-string [of noëtic symbols] is the organic species existing in the Explicate Order. And such a significatum is then by definition isomorphic with its signifying string of symbols, its significans.
In fact, what is the case here is that we have merely defined noëtic-strategy-strings-having-a-meaning. We're not saying that such a strategy-string acquires meaning only after it is projected into the Explicate Order resulting in the actual existence of its significatum. The Implicate Order is timeless, so the meaning of its strategy-strings 'was always there'. Only from the viewpoint of the Explicate Order we may say that a noëtic strategy-string has 'acquired' a meaning after it has actually appeared in this Order.
We have said that many noëtic patterns that, although being generally predisposed to the Explicate Order, cannot (among others that can) as such be projected into that Order (because they cannot as such exist there), may develop into strategies (which then can be so projected). And this development is supposed to take place through noëtic reactions. And these reactions inherently tend to produce strategies, and are driven so by the general tendency ('aspiration') of all noëtic patterns to attain ontological completion.
However, this idea of noëtic strategies in the Implicate Order to be developed from non-strategies runs into difficulties because we then are forced to let re-appear the phenomenon of ecological setting of some strategy in one or another guise in the Implicate Order. As we shall see later, this opposes against the true nature of the latter Order. We'll solve the problem of the origin, or of the existence, or of the development, of organic strategies in the Implicate Order by still further concentrating on the phenomenon in noëtic patterns to have meaning as a result of their inherently tending to become ontologically complete. And the true role played by ecological setting becomes clear after having considered the following instructive analogy (its validity and its limits).
An analogy.
In order to better visualize the ecological setting of organic strategies (organismic species) in the Explicate Order we may consider an analogy.
True strategies -- neatly and stably fitting, upon projection, in some existing ecological context or niche (in the Explicate Order), a context that really constitutes their ultimate existential condition, and thus these strategies each appearing in the form of a multitude of individuals of a given organismic species -- may be compared with snow-crystals being generated in normal atmospheric conditions (that is, especially normal atmospheric pressures and (winter) temperatures in which ' Ice-I ' is stable, where the latter is the normal form of ice, while fundamentally other forms are stable only at very high pressures not occuring in the earth's atmosphere).
Like all the different organic species do, these snow-crystals [they are extensively discussed in Fourth Part of Website ] all possess a common basic structure, which here consists in them being structured by a specific coupling of water-molecules resulting in a particular configuration of them, a configuration having a symmetry as described by Ice-I 's Space Group. In organisms, on the other hand, this common basic structure consists in the possession by all of them of DNA and its associated universal Genetic Code, and in their carbon-based chemistry.
And while organisms appear as different morphological, physiological, and behavioral, types (different material strategies) despite their common basic structure, snow-crystals also appear as different morphological types (concerning their shape, all being different variations on a common hexagonal groundplan), despite their common internal structure.
And while the different organic types -- organismic species -- are a response to correspondingly different ecological conditions or niches, the different snow-crystals are a response to correspondingly different atmospheric conditions (temperature and humidity).
So while the prevailing ecological conditions constitute the existential conditions of given organismic species (material strategies), conditions that is, prevailing somewhere in the Explicate Order, the prevailing atmospheric conditions constitute the existential conditions of given types of snow-crystals, that is, existential conditions, prevailing somewhere in the Earth's atmosphere, of freely floating solid H2O (water).
From all this we can now pursue the analogy further as to the projection of strategies from the Implicate Order into the Explicate Order.
As was established in Fourth Part of Website , snow-crystals are holistic entities in virtue of the fact that the six antimers [counterparts, all coming together in the main axis of the crystal] of each one of them are [in any given individual snow-crystal] non-locally coupled : If a branch (an outgrowth) of a certain morphological type is developed at one of the six corners of the hexagonal ice-plate of the snow-crystal, the same type will develop at the other five corners of that crystal as well.


Snow-crystals
(After BENTLEY and HUMPHREYS, 1931)
Even if one of these branches is longer or shorter than the other five, it is still of the same morphological type as that of those others. The growing crystal tries to attain and preserve some morphological equilibrium. So the generation of a snow-crystal, like all holistic structures, involves the Implicate Order, because as far as we know, any holistic feature of a given material structure is something 'extra-physical'.
Also any organismic species -- any material strategy -- has holistic features, for instance the non-local connectedness of functional parts in the body of an organism, especially evident in morphogenesis [individual development], evident that is, in the phenonena of regulation and regeneration. As in (snow) crystals, the w h o l e of the organism is a determining factor and as such an extra-physical factor.
A given organic strategy is formed in the Implicate Order in order for it to exist in the Explicate Order, and, more specifically, to exist in a certain relevant and compatible ecological setting ( Recall that a "strategy" is not a strategy possessed by the organismic species concerned. The strategy IS the organic species, that is, IS the total of all chemical, physiological, morphological, and behavioral features commonly possessed by that species' individuals). If this ecological setting is actually present in the Explicate Order, the strategy may be projected and will then appear as a growing population of individuals of some particular organismic species. So we may say that a particular organismic strategy 'crystallizes' in a correspondingly particular ecological environment, just like a particular snowcrystal-type appears -- crystallizes -- in a certain correspondingly relevant and compatible atmospheric setting.
So far, the analogy between snow-crystals and organic strategies holds fine. But organisms show evolution, while snow-crystals do not. Organismic species not only appear in appropriate ecological settings, but show a definite succession (consecutiveness) in their appearing in the Explicate Order, thereby not strictly following a supposed succession of ecological settings. Apart from some major transformations of those settings (for instance the evolutionary appearance of flowering plants [angiospermae] in the Cretaceous Period, or the first colonization by living forms of the terrestrial environment), making it possible for some special strategies to be finally fitted in, -- a simultaneous diversity of ecological settings has always existed, at least since the time that the colonization of the Earth by living organisms was well underway. So in the successive appearance of organismic species and types there is a non-ecological factor at work as well. As to its nature, this factor is (consequently) purely formal in the sense that the successive appearance of organismic species is partly driven by a formal factor (and thus not a physical factor). And this factor, because it is formal, not material, must ultimately reside in the Implicate Order. Indeed, it is precisely the earlier-described formal order in which the noëtically-produced strategies (strategy-strings of symbols) find themselves to be, an order that is, of formal derivability. That is to say, these noëtic strategies together constitute a formal system which we had called the " FSNSS " (formal system of noëtic strategy-strings), itself over-forming ontological number theory. And, as has been said, upon projection we see, in the Explicate Order, the evolution of organisms. And although they have, as a result of all this, a definite order of appearance, a given strategy only appears in its corresponding ecological setting where this setting is actually present in the Explicate Order.
We have described the appearance of material organic strategies in their corresponding ecological context in the Explicate Order, and have visualized this with the appearance of snowcrystal-types precisely there where their corresponding atmospheric conditions are present.
But the key-lock relationship between a given organic strategy and its proper ecological setting is at the same time also a constitutive factor playing a role in its very construction in the Implicate Order (but see later). In that Order the ecological setting is supposed to be present as a noëtic 'environment' simulating as it were such a possible ecological setting. The fitting-in into such an 'environment' is the constitutive force in constructing strategies in the Implicate Order (we shall revise this idea later), just like the prevailing atmospheric conditions (temperature and humidity) form the constitutive force in the construction of the corresponding snowcrystal-type.
But in organisms the particular ecological setting, as supposedly simulated in the Implicate Order, is not the only constitutive factor of strategies, because, judging from how things are in the Explicate Order, a given ecological setting may support a number of different strategies, that is, existence can be achieved in a given ecological setting in several different ways. This is a result of the organism's high degree of complexity, freeing it of too great a rigidity.
The internal structure, including its intrinsic symmetry, of a snow-crystal, like any crystal for that matter, represents a material lowest-energy state, a lowest-energy configuration of material elements, H2O molecules, -- lowest energy that is, within the confines of a certain thermodynamic range. Therefore, within those confines this particular configuration of material elements is a stable structure. So within these confines, within the range of these thermodynamic conditions, and only within these conditions, the crystal is stable.
If no other chemical substances interfere, the snow-crystal keeps on existing as long as these thermodynamic conditions (and conditions of humidity) prevail. No action by the crystal itself is required in order to maintain its structure. And within that thermodynamic range local variations may and will occur : small variations of temperature and pressure. Also chemically, local small changes may occur, such as changes in humidity which is the available amount of 'food' matter, H2O, out of which the snow-crystals are being built. To these local variations the growing snow-crystal responds by adjusting its shape (not its symmetry). This change of the crystal's shape is brought about solely by differential growth-rate resulting from differential accessibility of the different sites of the crystal to surrounding water-vapor : some sites of the crystal in a higher degree stick out into the nutritive environment and can thus receive and bond more water-molecules per unit of time resulting in a local increase in the crystal's growth rate, in turn resulting in a change of the shape of the snow-crystal, while its internal structure remains the same.
So the intrinsic structural pattern of the snow-crystal and generally that of solid water, ice, is able to exist as long as the mentioned environmental conditions prevail. The pattern is static, purely morphological.
But the intrinsic pattern of an organismic species (its material strategy) also, like snow-crystals, represented by individuals, here organic individuals, cannot permit itself to be static. It must actively maintain itself. If it doesn't do this, it will disintegrate. Therefore, such a pattern is not a mere particular configuration of material elements, but in addition consists of processes, it is dynamic. Indeed, an organism is a process-structure. And only as a process-structure of a particular kind it can exist, and here " it " is the process-structure itself, the organic strategy. And to be an organic process-structure means that it actively and, especially, selectively, deals with the environment. Also in the inorganic world such process-structures are known (much is written about them and their mathematical analogues in First Part of Website, Dissipative Structures ), such as the famous Belousov-Zhabotinsky Reaction. Here also 'food material' is selectively extracted from the environment (as is the case also in crystallization), but this (ongoing) reaction, like a growing crystal (but the latter being dynamic only at its periphery), does not take any special measures when the food-material surrounding it becomes scarce or depleted. Organisms, on the other hand, respond to such a situation by either actively fanning food-particles toward them (sessile marine organisms), growing their roots longer or moving their leaves (plants), or even go out in search of food (animals).
All these activities in their generality intrinsically belong to the process-structure, the organic strategy. This process-structure first of all consists of assimilation of appropriate, but nevertheless alien, elements (in contrast to crystallization), that is, accepting these elements and then transforming them into its own building-blocks and tools. But the acts of collecting these elements and transforming them, and the acts of excretion of matter and entropy, by the organism, at the same time bring with them the using-up of the organism's own internal resources of matter and energy : dissimilation. So these antagonistic processes must be balanced : The individual organism must be in metabolic equilibrium in order to maintain its overall structure. This active maintenance of metabolic equilibrium is an important element of the organism's strategy.
But this equilibrium cannot for alsways be maintained. Sooner or later it will be disturbed (by accidental or intrinsic factors). The individual will disintegrate and die. And so the strategy of the organism includes yet another factor to cope with this existential hazard : The individual will reproduce itself, guaranteeing the intrinsic pattern, representing the species, to continue to exist. In organisms this reproduction indeed plays a major part in the conservation of the species concerned, and is thus an essential element in the species' strategy to exist and persist. And also for reproduction special activity of the individuals is demanded, as we saw, for instance, in our exposition about wasps and ants in their care for their offspring. And indeed, all this activity, assimilation (counter-balancing dissimilation), and reproduction (counter-balancing the death of the individuals), can only take place successfully in appropriate ecological setting of the organismic species concerned.
And only when this reproduction of individuals fails, or fails to be sufficient, as a result of changing ecological factors, new measures must be taken. However, the species as such cannot reproduce itself (in order to maintain life), but must transform into one or more new species. In the context of our theory we say that new strategies will appear in the Explicate Order. And even without the mentioned failure, new species may nevertheless appear, in order to increase organismic diversity, and so guaranteeing Life to persist, because diversity protects Life against it being extinguished by possible environmental catastrophs. And the appearance of new strategies constitutes organic evolution.
But, as has been said, organic strategies cannot be constructed in the Explicate Order, they are constructed in the Implicate Order as a result of directed noëtic reactions, directed to produce precisely these strategies. And earlier we suggested that the noëtic counterpart of the necessary ecological setting (embedding) of such a strategy will be a constitutive factor or force of this strategy, that is, this setting partly determines that strategy as to what strategy it will actually be. We were tinkering with the idea of 'noëtic environment' simulating such an ecological setting (inspired by the snow-crystal analogy). However, this image will not do, because it suggests with the term "noëtic environment" some quasi-spatial relationship obtaining between a noëtic strategy and its noëtic environment. We may not, in theorizing, repeat, duplicate or copy given essentially spatial configurations (such as ecological settings) of the Explicate Order in the Implicate Order, because the latter is non-spatial in its very nature.
So in order to understand the existence of strategies in the Implicate Order, or (noëtic) generation of them from non-strategies in that Order, we must think along other lines.
In what follows we shall try to work out such another line.
As has been expounded earlier (part LXa, LXb, and the present part, LXc) the Implicate Order is in fact 'ontological (natural) number theory', and the essence of what natural numbers in fact are is described by TNT, the formalization of number theory. And noëtic patterns in the Implicate Order are in fact strings of numbers. But, of course, any sequence of numbers whatsoever is then a number-string, a noëtic pattern. This would imply an infinity of noëtic patterns to be present in the Implicate Order. However, there are factors -- ultimately based on the noëtic patterns' tendency to attain ontological completion by coming to exist in the Explicate Order -- that prevent such an unlimited proliferation of noëtic patterns. There are in fact two categories or types of noëtic patterns (number-strings) :
1. Number-strings expressing basic relationships between (natural) numbers. These number-strings all belong to TNT and together constitute the true nature of (natural) numbers. They have no meaning other than themselves, that is, they coincide with their significatum (= that what they mean) (significans = significatum), which is the same thing as saying that all the TNT number-strings, that are about what numbers are, have no genuine meaning at all. They do not signify or mean the essence of numbers (and with it the essence of the Implicate Order), but CONSTITUTE this essence, ARE, this essence.
2. To the second, and at the same time last, category or type of number-strings belong the ones that truly have meaning (and that this is an 'easy' to obtain property is because all pure forms tend to become ontologically complete, that is, to become existents of the Explicate Order), that is to say, they do not coincide with their significatum (significans not identical to significatum). They have their significatum (that what they mean) outside themselves. And where, then, can we find this significatum? Well, of course, in the Explicate Order. Only then the significatum of the number-string does really lie outside it. And only then this string distinguishes itself from other strings or from other numbers, that is, only then it is a true noëtic pattern. In fact such number-strings are descriptions of possible states of affairs in the Explicate Order. And also when the significatum of a given number-string is 'not yet', or 'not anymore', present in the Explicate Order, the absence of this significatum is only seen as such from the perspective of the Explicate Order, because only there we can say that it 'now' is not present. From the perspective of the Implicate Order, on the other hand, the significatum is 'always' present in the Explicate Order. And we now know why any number-string (if being a truly distinguished unit, and if it is not a TNT number-string) has its significatum in the Explicate Order : Any noëtic pattern naturally tends to become ontologically complete, that is, to become material, and consequently to become an existent of the Explicate Order.
So all non-TNT number-strings have meaning, and are therefore descriptions of states of affairs in the Explicate Order. The meaning of the description (number-string), a description which is not written by someone or something -- ( Because of the fact of them necessarily having a meaning in the Explicate Order and together with the connected fact that they intrinsically tend to ontologically complete themselves, the descriptions come forth spontaneously, without any 'designer'), -- lies outside the description itself, it lies in the Explicate Order. And a part of these spontaneous descriptions will turn out to be strategies. And every such strategy will necessarily refer to the Explicate Order, because they all have their significatum in that Order. So they are true and relevant strategies. They are not internally random, that is, they are not randomly produced, but true and relevant prescriptions of how these very prescriptions themselves may exist (as isomorphic material strategies) in the Explicate Order.
So thanks to the fact that non-TNT number-strings do only exist in the Implicate Order when they have meaning (and so are indeed definite and delimited number-strings, noëtic patterns, distinguished from other such number-strings), that is, when their significatum lies outside them, and thus lies in the Explicate Order, -- all strategies that do exist (as number-strings, noëtic patterns, noëtic descriptions) in the Implicate Order, are true, genuine, strategies, fully relevant to the Explicate Order, and so not existing side by side with a multitude of randomly-produced quasi-strategies, or generally, quasi-descriptions.
So we have now, at last, solved the vexed problem of the "Origin and Development of true Strategies in the Implicate Order", and have seen that the strategies have not necessarily "originated" from some non-strategy. We can safely say that one or more such strategies were there in the Implicate Order right 'from the beginning', and that many other true strategies have been produced by noëtic reactions between existing strategies (or other noëtic patterns). Of course, these noëtic reactions do not take place in time, and moreover only exist when their products have meaning in the sense described above. Otherwise they would not be distinguished number-strings and thus not be products at all.
The above discussed analogy between the meteorologic stability-fields of snowcrystal-types and the ecological stability of material strategies (organismic species) indeed only holds for states of affairs in the Explicate Order. Only there it may be instructive. So while we may say, thinking about this analogy, that strategies 'crystallize' in their proper ecological settings and are stable there, we may not transpose such an image to the Implicate Order.
Although a noëtic strategy-string (each strategy being a string of symbols, numbers) is a relevant description of how for it to exist in the Explicate Order, it cannot just like that exist in that Order. From the perspective of the Explicate Order it can only do so when it can be received in its proper ecological setting or environment, and the latter must then be present. So we cannot expect projections of strategies (from the Implicate into the Explicate Order) to take place in some random way. And because projected strategies (organismic species) play a chief role in the constitution of ecological settings of other strategies (organismic species), a certain definite sequence or succession of projections may be expected, especially a succession beginning with lower strategies (primitive organisms) followed by higher and higher (that is, more sophisticated) strategies (higher organisms), and this is indeed accomplished by the earlier-described fact that the noëtic strategies find themselves to be ordered according to some formal system in which lower strategies are formally derivable from higher ones (by subtraction of elements), followed by a projection of them (into the Explicate Order) in reverse order (first primitive organisms, then higher organisms). And this then IS organic evolution, and in it ecological settings are more or less gradually built up in virtue of the ordered succesive appearance of organismic species that can going to make up these settings.
As to the inorganic setting of things (things existing in such settings) in the Explicate Order, we may hold that
the Explicate Order is simply there (together with the Implicate order). This means that there is an existential domain having a space-time framework. And then all the things in this Order that are directly or indirectly implied by this space-time framework, such as individuals and individual cases, matter, gravity, radiation, movement, thermodynamic conditions, etc., also exist in the Explicate Order just like that. But insofar there are holistic aspects in these things and features, these aspects will involve the Implicate Order (such as the holistic aspects of crystals). And it is especially organisms that display many holistic aspects, so they are irretrievably connected with the Implicate Order in addition to their purely physical, material, non-holistic features connecting them with the Explicate Order. And, as we saw, especially organic evolution is irretrievably connected with the Implicate Order as our theory has described it.
Origin and Development of Organic Strategies in the Implicate Order Revisited.
We return to the question of how strategies come to exist, as noëtic patterns, in the Implicate Order. To answer it we must make use of analogies, such as "language", "description" and "meaning", without in fact knowing whether such things actually exist in the Implicate Order. Further, we have 'ontologized' certain concepts of mathematics and mathematical logic : We did this with the concept of meaning, with the concept of symbol, and with the concepts of (natural) number and number theory. Finally, we were forced to repeat earlier findings and assumptions in order not to lose track of our complicated theory.
Assuming (as we have done) that "noëtic space" (as we may call the Implicate Order) is in fact "number space", where numbers, i.e. natural numbers, objectively exist in some way, the Implicate Order IS "ontological number theory", which here means not a 'theory' in the usual, cognitive, sense, but a coherent structure consisting of natural numbers. And it is here supposed that its very essence (like that of "ordinary number theory" as a branch of mathematics) -- this essence being the intrinsic whatness of natural numbers -- is, in Mathematical Logic, described by formal typographic number theory, TNT. The typographical symbol-strings of the latter, as well as their transcription into Gödel-numbers, describe what natural numbers essentially are. And, correspondingly, something like Gödelized TNT must reside in the Implicate Order as the true kernel or essence of ontological number theory. This kernel thus consists of a set of certain "number-strings" which we may call " TNT-strings". In what way numbers do actually exist in noëtic space, we of course don't know. But that doesn't matter so much. For convenience we may assume that they are composed of digits ('symbols') which together form digit-strings with place-value of digits as in the decimal number system. And because individual digits represent numbers as well, we may call digit-strings also "number-strings". And, as has been said, some of such strings form the set of TNT-strings, constituting the kernel of number theory. These TNT-strings, TNT-number-strings, have no meaning beyond themselves. If they have one, it resides in the Implicate Order only. All remaining number-strings, however, are only then true definite intrinsic strings (with an intrinsic beginning and an intrinsic end) when they do have a meaning in the Explicate Order ( Their significata, as seen from the timeless Implicate Order, existing in the Explicate Order [even when they happen not to exist there at some given point in time seen from this Order] ). This meaning, then, that is, the meaning of number-strings, lies in the Explicate Order, but to conceptually grasp this semantic phenomenon we must artificially insert an extra, intermediary, semantic stage. For this we semantically connect with such number-strings some one existing language, say, English. And then we consider every individual, say, triplet of consecutive number-symbols existing in such a number-string to be a Gödel-number (a number coding for something), each, consequently, consisting of three digits, a Gödel-number that is, arbitrarily (but then constantly) assigned to some definite typographical symbol of the English language : letters, interpunction-marks, etc.
So, for example, the string "512,123,210,811,754" (not reading the commas) means the word "CAUSE" after earlier having assigned "512" to "C", "123" to "A", "210" to "U", "811" to "S", and "754" to "E" (and other number-triplets having been assigned to the remaining letters and interpunction-marks of the English language). And the word "CAUSE" in turn has a meaning in the Explicate Order.
Having done so, we consider only those number-strings that then possess, as a result, some r e a l meaning in the English language, such that these number-strings in fact point to elements, forms, or conditions in the Explicate Order. Each such (non-TNT-)number-string is then a word, a sentence, or a sequence of sentences, in that language, and of them the more complex number-strings are then descriptions, descriptions in that same language. But a description of what ? Well, a description of general patterns or conditions as they exist in the Explicate Order ( "exist" as seen from the Implicate Order, that is, persist as seen from noëtic space). So, and now skipping again (for a while) the intermediate meaning-stage (the English language) of the number-strings (in the Implicate Order), we can say that the mentioned general patterns and conditions as they exist (and persist) in the Explicate Order are the meanings (significata) directly of those number-strings. And from here we can see that -- adding back again the meaning of them in the English language -- transforming one description into another is in fact some arithmetical transformation (namely from one number-string [i.e. number] into another). This is especially the case in number-strings whose meaning is a material (organic) strategy-to-exist-in-the-Explicate-Order (in this Order appearing as organismic species). And the number-patterns representing such strategies now turn out to be formally transformable into one another by derivational operations involving special arithmetical rules. And so it is that these number-strings -- descriptions of strategies -- together turn out to constitute a formal system in which 'theorems' are noëtically produced from other 'theorems', and ultimately from 'axioms', by applying relevant rules from the system's set of arithmetical production rules. Upon projection, then (from the Implicate Order into the Explicate Order), we see in the Explicate Order organic evolution to take place. But, as has been said earlier, it is not so that in this way the (noëtic) strategies are actually created. They merely happen to form a system with an inherent order of formal derivability.
How, then, are these strategies actually created in the Implicate Order? This is a deep and important question, because, as we can see in their Explicate-Order-counterparts (organismic species), they always are very complex, and especially, very 'sophisticated' indeed. We ask, and try to answer, this question, because it is problematic to suppose that these strategies are 'just there' (or, even worse, are made by some 'intelligence'), just like that, in the Implicate Order. In fact, they do exist, just like that, in that Order, and trying to establish how they are 'formed' there, should in fact mean trying to find out in what way they exist there just like that, and thus not being formed there but (how they are) constituted there.
We may safely assume that the dimensions of noëtic space (the Implicate Order) are ultimately qualitative in their nature as to meaning-possessing number-strings residing in it. This implies that the constituent numbers (elements) of such a string may be scattered over wide 'areas' of noëtic space. And this fact makes it virtually impossible to actually graphically depict the noëtic creation of any given strategy, that is, creation by way of it (= the beginnings of that strategy) appropriating certain initially (to it) foreign elements, elements, that is, of, and taken from, the inorganic conditions (as they are present in enfolded ('coded') form in the Implicate Order), and also of elements of other strategies, which noëtic act of appropriating is the very noëtic counterpart of organic adaptation in the Explicate Order. So for graphically depicting this, we must resort to some spatial diagram depicting, more or less symbolically, the creation of organic strategies in noëtic space. We will do that in due course.
As has been said, each (non-TNT-)number-string, if it is to be a definite number-string (with a beginning and an end) at all, must have a significatum persisting (as seen from the timeless Implicate Order) in the Explicate Order. We can see this significatum as being the (true) meaning of a description present as a number-string in the Implicate Order.
Now, a loose group, or mere incoherent aggregation, of disconnected mere numbers or of short number-strings, may become, that is, may come to constitute, a fully-fledged true and coherent number-string, representing a strategy, in four ways :
The mentioned appropriation, encapsulation or seizing of alien noëtic elements by some given incoherent collection of [shorter or longer] number-strings (and the dropping of some, or the rearrangement of some of its own elements), or the appropriation of parts of already existing (in the Implicate Order) strategies (descriptions), or the encapsulation of whole such strategies (descriptions) is, we might say, a throughout ' r a t i o n a l ' process. The initial incoherent group of 'words' (short number-strings with meaning in the Explicate Order) seeks to become a true coherent description (= having a definite meaning in the Explicate Order) by seizing alien noëtic elements, etc., this seizing ultimately driven by the 'desire' to create a noëtic pattern that is such that it can, upon projection, acquire o n t o l o g i c a l c o m p l e t i o n, that is, the mentioned incoherent aggregation of number-strings seeks to become a true strategy (to exist in the Explicate Order). The mentioned processes of inclusion, dropping, or rearranging, do not, however, take place when the result would fail to be a coherent description having its meaning in the Explicate Order. So all compound noëtic patterns finally existing in the Implicate Order are true strategies, strategies 'formed' in virtue of the mentioned processes, that is, by self-organization of pre-existing elementary noëtic forms. And because such self-organizations are timeless, we can say that all strategies do 'already exist' in the Implicate Order. And 'subsequently' they -- as has been expounded earlier -- together turn out to constitute a special formal system, a system, that is, of strategies that are noëtically related to one another in virtue of their formal derivability from one another. And, as has been said, this derivational order determines the "noëtic trajectory" leading from higher [more sophisticated] strategies to lower ones. This is 'e-volution' the literal sense of the word. And the subsequent projection of these derivationally consecutive strategies will then take place in reverse order, resulting in the evolution (in its present sense) of higher organic forms from lower ones, in the Explicate Order. And this successive appearance of organismic species (micro-organisms, as well as plants and animals) is at the same time responsible for the ongoing construction, transformation and destruction of ecosystems.
We spoke about the 'desire' of noëtic forms (patterns) [number-strings] to acquire "ontological completion" (= going to in-form [prime] matter (as a substrate), and becoming thereby individual, local, and temporal ), and said that this completion is realized after projection of that noëtic form into the Explicate Order. And indeed, when it is so projected (unfolded), it exists as a material pattern in the Explicate Order, and as such being then the significatum of that noëtic form (number-string), that is, its meaning then existing in the Explicate Order. But for a noëtic form to be ontologically complete, this (actual existence) is already more than sufficient. If a given noëtic form or pattern truly is a description, residing as such, that is, as description, in the Implicate Order, then it has, as seen from this timeless Order, a significatum persistent in the Explicate Order, even when at some given point in time, or during a certain period of time (now seen from the Explicate Order), such a significatum does not actually exist in the Explicate Order.
To the Explicate Order the projections of noëtic patterns take place at definite points or periods in time, and in a definite successive order.
Not so to the Implicate Order. For it a true description or strategy has, by implication, a definite meaning in the Explicate Order, and so already is ontologically complete. For the Implicate Order the projections are timeless as to 'when' they take place, they are just projections.
We may depict the creation of a strategy in the Implicate Order spatially as follows :
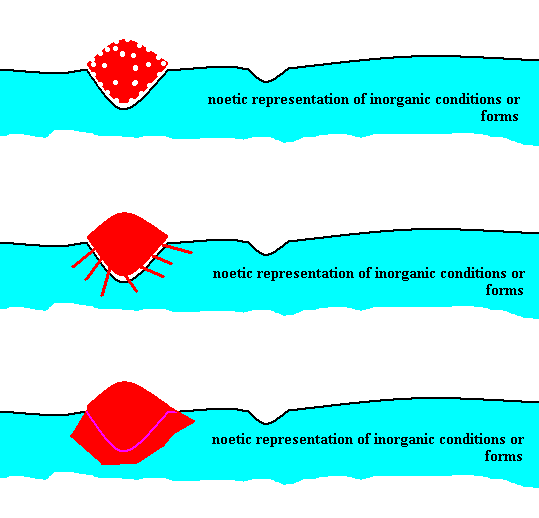
Figure 1 : Diagram depicting, reading from top to bottom, the noëtic transformation of an initially incoherent aggregation of numbers and number-strings (red) into a single, true, fully-fledged coherent number-string, a description, having a definite significatum in the Explicate Order. For this, the initial aggregation (red) appropriates (seizes, takes up) certain elements from the noëtic representation (description) of inorganic forms and conditions (blue) (without, of course, these elements being lost for these latter forms and conditions). The resulting coherent number-string (tangle) (bottom image, red) represents the result of 'adaptation' of (part of) the initial incoherent aggregation to inorganic forms and conditions. The initially incoherent number-string (tangle) has now become a true strategy : a strategy prescribing how for this very strategy itself to exist in the Explicate Order.
A new strategy, may, as noted earlier, also be created in the Implicate Order when it seizes elements of other existing strategies, existing that is, in that Order :
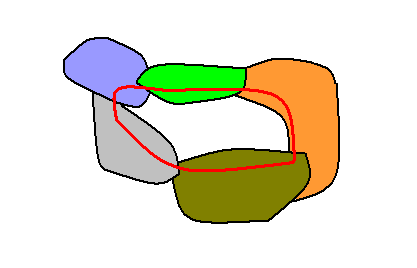
Figure 2 : Diagram depicting the creation of a new strategy (encircled by a read curve) amidst other already existing strategies : An initially incoherent aggregation of noëtic elements organizes these elements and takes up elements from five other existing strategies (which do not, of course, lose these elements), resulting in a sixth single (tangled) number-string. And if this new number-string is a prescription of how for this very description itself to exist (i.e. to unfold) in the Explicate Order, then it is a (new) strategy, existing (in the Implicate Order) together with the original five strategies (drawn as colored areas).
Every such new strategy, is, moreover, as all organic strategies, dependent on, and adapted to, certain inorganic elements, forms, and conditions, first of all being so dependent through such dependencies of the other strategies on which the new strategy depends, but also directly so dependent. We may symbolize all these dependencies by arrows :
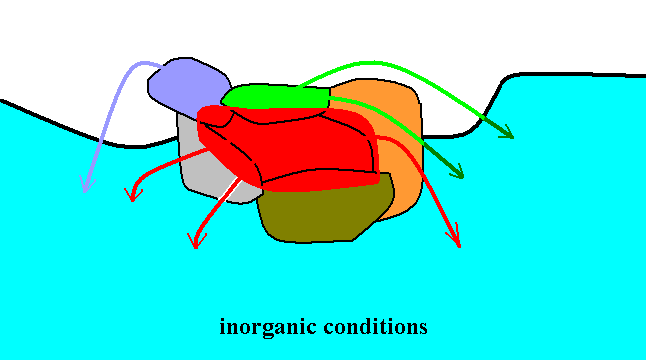
Figure 3 : Diagram depicting the direct and indirect dependencies (arrows) of the original strategies and of the new strategy (red) upon inorganic elements, forms, or conditions.
Projection into the Explicate Order.
Turning now to the projection of these noëtic strategies into the Explicate Order, we must expound what such a projection in fact is.
While in the Implicate Order there is no difference between principle and that what is determined by it, that is, between category and its concretum (because in the Implicate Order there are no individual cases and no accidental structures), the Explicate Order contains categories ( If / Then constants), each determining its own concretum. The complete set of such categories determines what, in fact, the Explicate Order really is (characterized by its set of determinative elements). And, in addition, the Explicate Order has several 'categorical layers' each with categories of their own. For the time being we distinguish two such layers, the Inorganic Layer and the Organic Layer. And in addition to categories that are commonly possessed by these two Layers, such as the categories of Space and Time (and some others), they each have their own specific categories. They, in determining their specific concreta, at the same time collectively determine what these Layers essentially are (as characterized by their determinative elements).
So upon projection into the Explicate Order, the noëtic strategies (and other noëtic forms) come to stand under the Explicate Order's categories, having them, first of all, unfolded along space and time dimensions, and having them also determined by the other general categories of the Explicate Order. Certain noëtic patterns will be projected into the Explicate Order's inorganic category layer and will then unfold into physical forms and conditions. Other noëtic patterns, on the other hand, such as organic strategies will be projected into the Explicate Order's organic category layer. Also they will come to stand under the categories commonly possessed by the two layers, but in addition, also under the special categories of the organic layer.
Any true noëtic pattern will, when the conditions of reception are right, be projected. And then a sequence of alternating injections and projections will take place as long as the mentioned conditions (in the Explicate Order) remain right. When they cease to exist, the sequence will be terminated. And in the case of organismic species we see this as their extinction.
About "projection into the Explicate Order" we can theorize much more than has been done so far. We will do this now.
Organic strategies exist in the Implicate Order as noëtic patterns, which are just Forms without matter. And these Forms are descriptions of how, for these very descriptions themselves to exist in the Explicate Order and so obtain ontological perfection, because there these Forms 'in-form' Prime Matter resulting in real beings with all the paraphernalia that are implied by the fact that each such Form now exists in the form of individuals in Time and Space. Existing in the Implicate Order, these Forms, strategies, are not actually derived from each other, but already pre-exist in the way described above and constituting just a set of strategies. And this set turns out to have a derivational structure, meaning that its members are formally derivable (not derived) from each other such that lower strategies are derivable from higher ones. So they form a branched sequence of strategies, and the members of this derivational sequence will be projected in reverse order into the Explicate Order, visible there as a branched evolution of organisms. But because a given strategy can only project into the Explicate Order when it can be received there in an existing appropriate ecological setting or context, the sequential order of the successive appearance of organismic species in the Explicate Order does not precisely reflect the (reverse) derivational order of them in the Implicate Order. But in the main lines it does. The projection of strategies was diagrammatically depicted in the last drawing of the previous document , not depicting, however, the mentioned imprecision of the reflection of the (reverse) consecutive order in the Implicate Order in the successive order in the Explicate Order. We reproduce that Figure here :
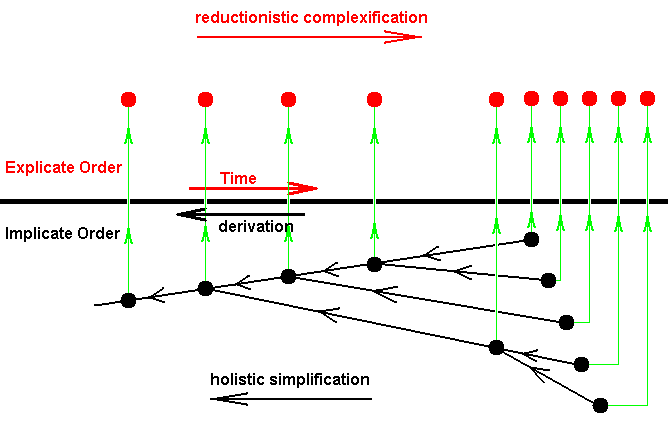
Figure 4 : Diagram depicting the projection of different strategies from the Implicate Order into the Explicate Order.
We will now discuss the phenomenon of projection more closely. When relevant and appropriate ecological conditions actually exist in the Explicate Order, a copy of some definite strategy will be projected into the latter Order. This means, that seen from the Explicate Order this strategy appears at some definite point in time of the Explicate Order's history. And as long as the mentioned ecological conditions continue to be present, that strategy will continue to exist in the Explicate Order. It will exist there as a certain organismic species. And the strategy of this species, or better, this strategy embodied by the individuals of that species, may represent a particular instance of a more general strategy of some higher taxon, say a family. Projection of such a strategy is always directly followed by its injection back into the Implicate Order. Also this injection is only an injection of a copy of the strategy. And this injection is again directly followed by a projection again into the Explicate Order, and so on and so on. Here we have to do with a continuous series of projections and injections. Only when the existing ecological conditions in the Explicate Order cease to exist, this series will come to a halt. While the mentioned series of projections and injections is 'seen' in the Explicate Order as the continuous existence of the organic species from the point of time of its first appearance (i.e. of its origin), in the Implicate Order this is not a temporal series. The next diagram depicts the appearance and persistence of a given organismic species resulting from the mentioned series of projections and injections :
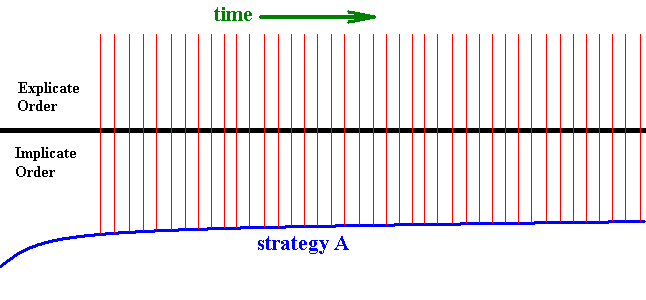
Figure 5 : Diagram depicting the projection of a given strategy, the strategy A, and the latter's continued existence in the Explicate Order since the point in time of its first projection. Each vertical line symbolizes a projection plus a subsequent injection. Although drawn as a discrete series, these lines in fact form a continuous series (of projections and injections). Seen from the Implicate Order, this series is not a successive series in time, as it is seen from the Explicate Order.
Now we may theorize that injection takes something back to the Implicate Order in addition to the original strategy. In fact, the strategy as residing in the Explicate Order, will be changed a little in the course of time, because it is subjected there by local and individual influences, such as external agents and because of its intrinsic degree of instability. This is in fact what we see in organisms : in addition to changes that cannot be transmitted to the offspring, the organism is subjected to genetic mutations. Some of these will be transmitted to the offspring or to a part thereof, and because the genome (the hereditary material) of an organism (but not its genome only!) contains information not only as to what the individual, carrying this genome, in fact is, but also what the species, and thus the strategy in fact is, this strategy will keep on changing -- albeit only little -- as long as that species exists in the Explicate Order. And because it is a process of change taking place in the Explicate Order and by the agents of that Order, it is not a formal process, but just a physico-chemico-biological process. And where such changes have biological significance, natural selection will affect the phenotypical result of these changes. But because the only genetic mutations we can think of are random genetic mutations, forming successive, but random series, often interfering with each other (as far as such mutations become phenotypically expressed at all, and as far as they are not immediately lethal), they will never result in long-term directed changes in the phenotype, meaning that the history of changes of a given organismic species is not some straight line or a bundle of such lines, but just an irregularly zig-zagging sequence of unrelated small changes, not leading anywhere whatsoever. All sophisticated changes and adaptations, as we see them in all organisms, come from the Implicate Order in the way described. The Explicate Order cannot bring forth or sustain such changes. So the only thing what we see in a given organic species is a slight morphological (and maybe also physiological and ethological) drifting during its existence. All the characters of such a species largely remain what they were, only their states will slighly change. As an example of such drifting we may mention the gradual shortening or widening of some forks in the wing-venation of some given insect species, or the partial or complete reduction of some particular vein. Only for such more or less simple transformations we may assume some degree of rectilinearity or unidirectedness in the series of elementary changes constituting such a transformation. These transformations, which we may call "explicate order transformations", remain within the strategy in which they occur. They do not lead it to another strategy. Further, they are expected to be present in all species of a given taxon. That is to say, the character-to-be-transformed is, in its generality, present in all these species, but its state differs in them. So the character actually changes by substituting one state of it by another. This step-by-step substitution of one character-state by another such state of the same character, say, of character "Zetha" (i.e. this transformation of character Zetha), may take place, independently, in all species of a given taxon, but possibly with different speeds and with a slight qualitative difference existing between the corresponding transformational series. So in every species of such a taxon we see the sequence Zetha-1, Zetha-2, Zetha-3, Zetha-4, etc. And letting this taxon to be the strategy (organismic species) A of our previous Figure, we can depict this "explicate order transformation" in it as follows :
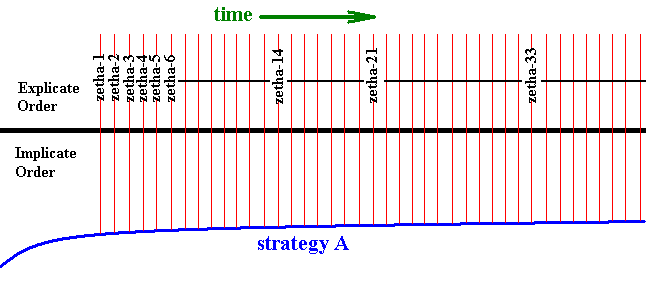
Figure 6 : Diagram depicting the projection of the strategy A.
A certain character Zetha of it changes its state as time goes by, resulting in the character sequence Zetha-1 ==> Zetha-2 ==> Zetha-3 ==> Zetha-4, etc.
and more simply we can depict this as follows :
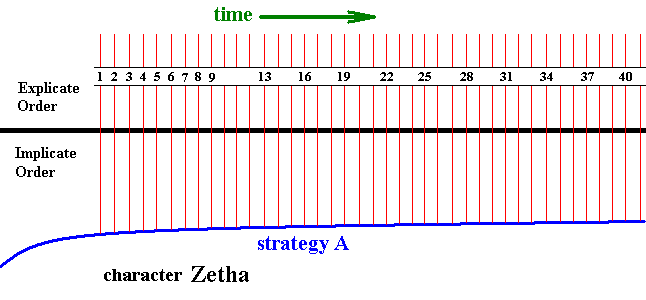
Figure 6a : Diagram depicting the projection of the strategy A.
A certain character Zetha of it changes its state as time goes by, resulting in the character sequence Zetha-1 ==> Zetha-2 ==> Zetha-3 ==> Zetha-4, etc.
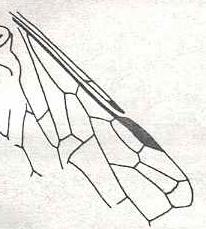
Characters like Zetha we find, for instance, in the wing-venation of insects. There, in every taxon, say an order or family, we can see, or reconstruct, in each case, the specific fate of each of the seven main-vein systems : the fate of the Costa (C), the fate of the Subcosta [subcostal system] (SC), that of the Radius [radial system] (R), of the Radial Sector (RS), of the Media (M), of the Cubitus (Cu), and, finally, the fate of the Anal system of veins (A). Each such main-vain system changes its state in the course of time. And e x c e p t for broader, more encompassing, venational changes, -- such as extensive reduction or expansion of the venation, or [except] for very specific features such as the development of 'joints' in the Costa or elsewhere in the wing, the development of a large pterostigma (wing-spot), or the development of special folds in the posterior part of the wing-membrane, and, of course, the concentration of strong longitudial veins at the anterior wing-margin ('costalization'), etc., -- these changes do not, apparently, have any aerodynamic or mechanical significance. So they cannot be directed by natural selection. Also, they have no ecological significance. They, apparently, are spontaneous changes, and the precise nature and course of each one of them is certainly dependent on the very 'whatness' of the taxon in which such a change takes place. A given venational character may be subjected to such a gradual change in more than one sequence of character-bearers. These parallel sequences may be simultaneous or not, and the changes in them may be slightly different. It is roughly known what the most primitive state of each main-vein system is, that is, how they looked like in the first (winged) insects. Already at the origin of the (higher) insect Orders the seven main-vein systems appear in a more or less slightly transformed condition. And so it is, for example, with the fate of the Subcostal-vein-system (SC) in the lower Hymenoptera (Symphyta). Although it is not more than just an example (of the many existing cases), it is perhaps instructive to follow this fate within the (families of the) Symphyta (saw-flies and allies). For this, we cite from RASNITSYN, Origin and evolution of the lower Hymenoptera, published in 1969, but we here illustrate things with only one figure. (All fossil and recent superfamilies, families, and subfamilies that are mentioned in this example belong to the suborder Symphyta).
The most primitive state of SC we find in the subfamily Archexyelinae (Upper Triassic) of the family Xyelidae :
where the vein (the Subcosta), giving off several anterior branches (in impressions [Rasnitsyn here relating to fossil Hymenoptera] are visible 1-2 branches, but, possibly, their number was large), reaches the tip of the Costa [vein along the anterior wing-marging] before the pterostigma [wing-spot]. The next evolutionary stage is demonstrated by, apparently, Pararchexyela where the base of the last branch of SC bends down and unifies with the Radius (R) lying posterior to it, but then, at the same place, leaves it again and ends up at the end of the Costa, as in Archexyelidae. At the point of unification of SC and R originally there was a cross-vein, secondarily lost in Archexyelidae : merging of parallel veins usually is preceded by a shortening of the vein connecting them (more often of a cross-vein). Veins not connected by anything, seldom merge together.
The further evolution of the subcostal system is characterized by the extension of the region of coalescence of SC and R, such that of the distal branch of SC only its basal piece remains, often looking like a cross-vein between SC and R, and its end-piece, distally closing the costal field (as one calls the region between C and R in Hymenoptera). The end-piece of SC very often completely vanishes. It is well developed, often even widened, only in Macroxyelinae and, apparently, in Tenthredinidea. And here the tip of SC with its base tightly adheres to the pterostigma, such that the fracture ['venational joint'] of the forewing at the base of the pterostigma seems to go through inside it.
As a result of the described processes a straight SC with a fork at its end appeared [at the origin of the Symphyta], held to be typically of the Symphyta. In this, in the majority of forms also the extra anterior branches disappeared, being preserved only in some Xyelidae. In Caenolyda (Pamphiliidae), in addition, the anterior branch of SC was strongly lengthened and secondarily reached the end of C. However, also the typical two-branched SC is far from preserved in all Symphyta and is characteristic only of a part of the Xyelidae, of Pseudoxyela (Xyelotomidae?), Pamphiliidae, Xyelydidae, Gigasiricidae, some Siricidae, and, perhaps, Myrmicidae. The subsequent evolution of SC is in many cases characterized by a merging with R also of its basal piece, such that of SC only remains its anterior branch in the form of a cross-vein (in many Tenthredinidea, in Sepulcidae, Xiphydriidae, Anaxyelidae, some Siricidae), or even this cross-vein disappears. By the way, this latter result -- total absence of a free SC -- may [in other cases] also be accomplished by its direct reduction, without merging with R. This process took place in some Xyelidae, and, probably, also in Megalodontidae, Parapamphiliidae, Cephidae, Orussidae, and in some recent Siricidae.
Other (different) histories can be reconstructed for the Radius, the Radial Sector, the Media, etc. in Symphyta (See our document on fossil Hymenoptera ), and again other such histories of these main-vein systems can be found in other taxa of winged insects.
We will continue the above general considerations about the "explicate order transformations", but now with the wing-venation in insects constantly in mind, implying that the character "Zetha" represents one of the seven main-vein systems.
Because the wings of an insect are situated outside its main body, we may assume that the venational details -- apart from those 'broad' ones mentioned above -- change more or less independently of those of the insect's main body. That is to say, no other morphological or physiological change of characters interfere with the evolutionary transformation of the venational details. And to a certain extent this may even be true of each main-vein system, considered in itself, with respect to the other main-vein systems. So there is a more or less separate evolutionary 'development' of the Costal system (always consisting of only one vein, the extent of which may thereby change), the Subcostal system, the Radial system, the Radial-Sector system, etc. at least within the confines of a given lower taxon (such as a species or a family). Each insect Order of the Pterygota (winged insects) originally possesses the seven main-vein systems in the forewing (sometimes, and more often in the hindwing there is an additional main-vein system, the Jugal system ( J )). However, the general nature, the type or 'look', of all the main-vein systems together, that is, of the venation of a wing as a whole, changes from insect Order to insect Order. We can clearly see this when we go, for example, from Diptera to Hymenoptera, or from mayflies to dragonflies, etc. And, of course, then, the mentioned transformations in the details of the venation differ from Order to Order.
So the details of the wing-venation in insects go (in their transformation) their own way in each insect Order or lower taxon, without resulting in new strategies, that is, without resulting in new habits. There is no doubt that there are, in addition to the wing-venation, other such characters that independently evolve, although we do not expect them to be many.
Earlier we had interpreted an organismic species (or higher taxon) as a materialized strategy to exist in the Explicate Order. And as constituting this 'material' strategy we have reckoned-in the totality of all morphological, biochemical, genetical, physiological, and behavioral features of that species (or higher taxon). In fact this is the way that a pure immaterial strategy, a description, residing as a noëtic pattern or pure Form in the Implicate Order (noëtic space), manages to exist in the Explicate Order (material, physical space). Such strategies are forged in the Implicate Order as described above, driven by the force of ontological completion. They cannot be forged in the Explicate Order, because this Order doesn't have a 'rational' nature. But we now know that among the mentioned "totality of morphological, etc. feartures", together constituting a material strategy, there are some such features, co-constituting the organismic species, that are non-functional. A true materialized strategy must consist of (themselves more or less complex) components executing some particular function, that is, doing, or able to do, some work or task that neatly fits into a comprehensive web of tasks together guaranteeing existence, existence that is, of the given organismic species. So the non-functional features of that organismic species cannot belong to that very strategy. Or, in other words, the non-functional features, although themselves more or less complex components of the species' make-up, are not part of the strategy sensu stricto. And precisely because they do not belong to the strategy insofar true strategy, they do not come from the Implicate Order, they are not forged there. They are a product of the Explicate Order. We here do not directly mean the non-functional structures just as structures, but the non-functional changes of, and in them. In fact, all that comes to exist in the Explicate Order will be subjected to its physical or chemical agents, and will, as a result, slowly change in an uncoordinated way. Also a true strategy itself, constituted by a definite whole of integrated functional components, will tend to change slowly. But this only takes place within narrow limits, because uncoordinate change of a given strategy will not lead to a new strategy, but will be lethal. In fact it belongs to the very essence of a material strategy to be able to counteract noxious influences. So the true strategy, the strategy in the strict sense, is more or less buffered against such influences and so is relatively stable, as long as its ecological substrate remains intact. Change of the latter causes the strategy to vanish (locally or globally) and makes possible the projection (from the Implicate Order) of a new strategy, or the immigration of representatives of another strategy already living elsewhere. So a strategy in the strict sense, is always a product of the Implicate Order, while its small insignificant changes, not resulting in a new strategy, or changes that lead to its extinction (locally or globally) are products of the Explicate Order. And of course the (non-functional) changes (big or small) of non-functional structures (only insofar as they are really non-functional), or such changes of functional structures, are also products of the Explicate Order. Indeed, together with all the inorganic processes, they, although still being organic processes, represent the most conspicuous products of that Order. So, with respect to insects, we may roughly hold that the details of their wing-venation (and some other, not further considered characters), and the subsequent changes in them, are true products of the Explicate Order. And, as we may deduce from the study of existing wing-venations in insects, the venational transformations, as they are caused by, and take place in, the Explicate Order, are also confined within the morphological play of the main venational type constituted by the mentioned set of seven main-vein systems. And in each insect Order these transformations are further confined within the morphological play of the particular subtype of wing-venation inherent in, and typical of, such an Order. So here, that is, in the non-functional wing-venation, there is also some sort of buffering present, but certainly much less so than in the case of true strategies, that is, their morphological play is much greater. And, moreover, in contrast to changes in true strategies, changes in several main-vein systems in the same wing can take place more or less independently of each other insofar purely spatial conditions permit. As to precisely what transformations the main-vein systems will be subjected to in the course of time (via corresponding changes in the genetic material, or in other material of the body comparable to it, changes resulting from internal or external agents), depends first of all on the very essence -- whatness -- of the organismic species concerned, and secondly on the nature of the Explicate Order's agents acting on it. So it is to be expected that a more or less similar sequence of incremental changes of some given main-vein system will take place in different but related organismic insect species, although the rate of change may differ. Let us diagrammize, in the spirit of Figure 6a, above , the projection (from the Implicate Order into the Explicate Order) of two different but related strategies, ending up in the Explicate Order as two (different but related) insect species. The first appearance and continued existence of each of these species (and thus the continued alternation of projections and injections) are determined by the existence, in the Explicate Order, of their appropriate ecological context, a context that is, that can receive the projected strategy, resulting in the functional integration of the latter into the former. So one of these two related insect species may appear long before the other does, but if that first one has not become extinct before the time the second appears, there is a period of their simultaneous existence (in the Explicate Order). In the diagram it is supposed that a certain main-vein system is gradually changing in the course of time (during the existence of the insect species) in both species at approximately the same rate. The sequence of incremental venational changes is symbolized by a series of consecutive numbers.

Figure 7 : Diagram depicting the consecutive first projections (first red vertical line in the one, and first red vertical line in the other species), and the continued alternation of projections and injections, of two different but related insect strategies A and B.
The one venational character v (as such not belonging to the strategy sensu stricto) changes its state in both species as time goes by, resulting in character sequences symbolized by consecutive numbers. So we get two transformation sequences : v1, v2, v3, v4, ..., v41, ... and v1, v2, v3, v4, ..., v16, v17, ... .
In the above diagram strategy A is first projected at the point in time t 0, whereas strategy B is first projected at the point in time t 1. And because strategy A had its first projection long before that of strategy B, and because the rate of transformation of venational character v is here supposed to be equal in both species, the species A lies ahead, and keeps on lying ahead, of species B in this respect. So if we, with respect to venational character v, compare simultaneous individuals of both species, we see that the transformation of character v in the older species A has proceeded further than that in species B, implying that with respect to this particular venational character the later-appeared species B cannot formally be derived from the earlier species A , where a "derived species" here means a species that represents an evolutionarily more advanced state. When in setting up a system of phylogeny, one, of course, does not say that the individuals of one species have been evolved from those of another simultaneous species, that is from individuals simultaneously living with those of the other species. That's why one says "formally be derived". So when one is considering the actual evolution of species B from species A , one assumes that the transformational state of character v has remained the same in the past of species A. And then, in our present case, species B is more primitive with respect to character v (as it is in species A) and thus cannot be derived from species A , according to Dollo's law of irreversibility.
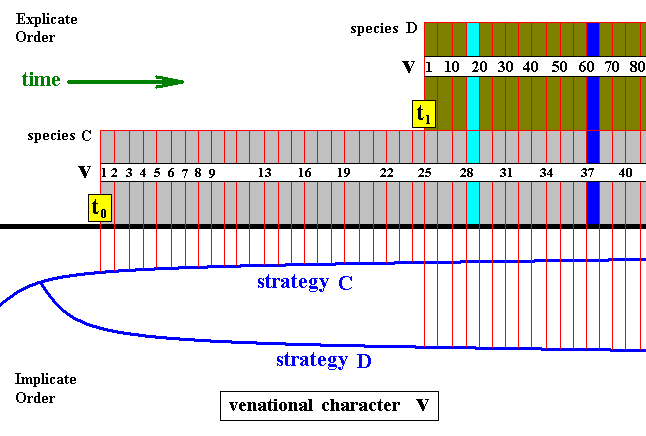
Figure 8 : Diagram, as the above diagram, depicting the consecutive first projections, and the continued alternation of projections and injections, of two different but related insect strategies C and D.
The one venational character v (as such not belonging to the strategy sensu stricto) changes its state in both species as time goes by, resulting in character sequences symbolized by consecutive numbers. So we get two transformation sequences : v1, v2, v3, v4, ..., v41, ... and v1, ... v10, ... v20, ... v30, ..., v40, ... v50, ... v60, ... v70, ... v80, ... .
If we now consider these two species at the time indicated by the l i g h t blue column, we see that, with respect to the development of venational character v , the later-projected species D still lags behind the earlier-projected species C . But when we consider them at the later time indicated by the d a r k blue column, we see that species D has catched up with species C , and even has progressed beyond it. So when looking at these two species at this later point in time, we see that, with respect to venational character v , species D is more advanced than species C and thus is the "derived species". And this is compatible with its being younger (if we think of the conventional view that the younger species is expected to be more advanced in its characters than is the older species).
Later we will see that many more or less primitive conditions of venational characters, as they are seen in various taxa (for example in Hymenopterous taxa), still exist today in recent representatives of these taxa. Have they persisted as such all the time, or have they re-appeared? If we assume that the transformations, of the type described above, steadily proceed from stage to stage, then it seems unlikely that some primitive stages have persisted all the way, from, say, Jurassic times up to the present time. And, moreover, also some relatively advanced states can be observed in certain jurassic fossils. This forces us to consider the possibility that certain primitive strategies re-appear long after they had become extinct a long time ago, carrying these primitive states with them. This seems to be confirmed by the general fact that of most higher taxa (everywhere among the animal and plant kindoms), many primitive members of them still exist today, either as (specialized) relics or as (still) blooming groups. So we must consider the possibility that the formal derivational order of noëtic strategies present in the Implicate Order only partly shines through in the system of recent organisms.
No precise congruence between (1) the formal derivational order of strategies in the Implicate Order, and (2) the actual (first) appearance of organismic species in the Explicate Order corresponding with these strategies.
Our noëtic theory of evolution, thus far developed, says that in the Implicate Order all the possible strategies are already present in the form of noëtic patterns, that is, pure Forms without matter. And whether they will be projected into the Explicate Order fully depends upon the existing ecological conditions or settings in the latter Order. When, seen from the Explicate Order, at time t these ecological conditions indeed appear, the corresponding projections may take place. These ecological settings are, as to their biotic factors, of course, co-developed by the projections of organic strategies themselves. In principle the consecutivity of projections of strategies belonging to a given higher taxon (the successive appearance of organismic species), follows the consecutivity of formal derivability (but then in reverse order), in the Implicate Order, of the strategies composing such a higher taxon (for example an insect Order or Family). However, the consecutivity of formal derivability will not exactly be copied in the Explicate Order because projection of a given strategy will only take place when an appropriate ecological condition actually exists in the latter Order. This implies that 'primitive' strategies of a given taxon -- strategies, that are situated at or near the 'beginning' of the branched derivational formal system (in fact at its end, because the direction of formal derivability is in principle opposite to the corresponding successive appearance of organismic species in the Explicate order) -- may appear much l a t e r than the more 'advanced' strategies of that taxon. This explains the well-known fact that the 'primitive' members of many taxa "still exist today". But it could also be that the primitive members of a given taxon indeed appeared first, but that later on their proper ecological conditions were destroyed as a result of either the extinction of certain organismic species, or of newly appearing organismic species, or as a result of changed abiotic conditions, or, finally, as a result of all of these together. And then these primitive members become extinct. But then, at a still later point in time of geo-biological history, the appropriate ecological conditions for these primitive organisms could return (as a result of extinction of other organismic species, or as a result of other causes), resulting in their projection once again into the Explicate Order. We may expect that all newly projected strategies (organismic species) will sooner or later become extinct, implying that we will never see a complete hereditary tree of some larger taxon being present in the Explicate Order at some point in time. Many species and even higher subtaxa of such a taxon will have become extinct during the history of that taxon. But some of them may re-appear when proper ecological conditions themselves re-appear.
The next diagram depicts the partly non-correspondence between (1) the formal derivational consecutivity of a given taxon in the Implicate Order and (2) the successivity of actual appearance of the members of that taxon in the Explicate Order.
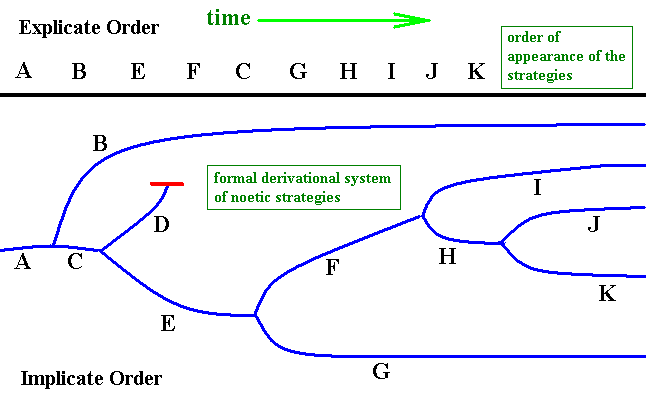
Figure 9 : Case of non-correspondence in projection. Diagram depicting the partly non-matching consecutive orders of (1) a derivational system of noëtic strategies in the Implicate Order and (2) the succession of actual appearance of the corresponding material strategies (organismic species) in the Explicate Order.
The (reversed) derivational consecutive order is assumed to be : A, B, C, D, E, F, G, H, I, J, K, whereas the actual successive order of appearance of the corresponding strategies is : A, B, E, F, C, G, H, I, J, K.
The strategy D did not project because of the non-existence of its proper ecological setting in the Explicate Order (when this setting later on happens to appear somewhere in the geo-biological history, the projection of strategy D will still take place).
In fact, as drawn, the formal derivational order is not linear, but branched : A, (BC), (DE), (FG), (HI), (JK). And if it would be that the formal and material consecutive orders exactly matched, the latter order would also be : A, (BC), (DE), (FG), (HI), (JK), or, taking into account that D is not projected : A, (BC), E, (FG), (HI), (JK). In the present diagram, however, it is taken that these orders do not match with respect to more than one strategies of this series.
But of course in many other cases the formal derivational system of a taxon in the Implicate Order will neatly be 'copied' into the Explicate Order. Or perhaps not? Experience in classifying organisms tells us that the noëtic system of strategies of a taxon (its formal derivational system) at least often leaves many traces of its consecutive order right in the Explicate Order. If this were not so, classification would be entirely impossible. But the all too well-known difficulties and uncertainties in the business of classification or of designing hereditary trees, show that in many cases there may be little correspondence between the noëtic and material consecutive orders.
In Part LXVI, on fossil Hymenoptera we will discuss the transformations of the wing-venation (as being explicate order transformations") further, especially the phenomenon of the 'take-over' (in the sense of continuation) of a given series of venational transformations from a given species to a new new species developed from the former.
(will be continued if necessary . . . )
This is then our noëtic theory of evolution so far developed, and so the end of our Summary, and with it the end of the Four-document Intermezzo on Noëtics.
[So all this also concludes our exposition (in earlier documents) of how formal systems can be embedded in number theory, and in what way certain strings of TNT -- the formalization of number theory -- can express things of other TNT-strings, and further what happens when the self-reference is tightened still more, namely the revelation that TNT is incomplete.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part LXI.
Back to Evolutionary Part XVII
Back to Evolutionary Part XVIII
Back to Evolutionary Part XXII
Back to Evolutionary Part XXIII
Back to Evolutionary Part XXIV
Back to Evolutionary Part XXV-A
Back to Evolutionary Part XXV-B
Back to Evolutionary Part XXV-C
Back to Evolutionary Part XXVI
Back to Evolutionary Part XXVII
Back to Evolutionary Part XXVIII
Back to Evolutionary Part XXVIII-A
Back to Evolutionary Part XXIX
Back to Evolutionary Part XXXI
Back to Evolutionary Part XXXII
Back to Evolutionary Part XXXIII
Back to Evolutionary Part XXXIV
Back to Evolutionary Part XXXV
Back to Evolutionary Part XXXVI
Back to Evolutionary Part XXXVII
Back to Evolutionary Part XXXVIII
Back to Evolutionary Part XXXIX
Back to Evolutionary Part XLII
Back to Evolutionary Part XLIII
Back to Evolutionary Part XLIV
Back to Evolutionary Part XLVI
Back to Evolutionary Part XLVII
Back to Evolutionary Part XLVIII
Back to Evolutionary Part XLIX
Back to Evolutionary Part LIII
Back to Evolutionary Part LVII
Back to Evolutionary Part LVIII