
e-mail :

This document (Part XXIX Sequel-34) further elaborates on, and prepares for, the analogy between crystals and organisms.
Philosophical Context of the Crystal Analogy (IX)
In order to find the analogies that obtain between the Inorganic and the Organic (as such forming a generalized crystal analogy), it is necessary to analyse those general categories that will play a major role in the distinction between the Inorganic and the Organic : Process, Causality, Simultaneous Interdependency, the general natural Dynamical Law and the dynamical system (intrinsic inorganic being). This was done in the previous documents. In the present document we will consider some organic categories.
Introduction
The entities of the Organic World, which are organisms, that is to say, organic dynamical systems, form a natural extension of inorganic dynamical systems (discussed in previous document), especially inorganic 'totality-generating dynamical systems', which are (inorganic) dynamical systems that generate, from a more or less disorderly collocation of elements, one or more coherent, inernally structured, spatial beings (such as crystals).
Organic dynamical systems -- organisms -- are different from inorganic ones, also from inorganic totality-generating systems, and especially from systems that generate crystalline totalities -- single individual crystals -- by the fact that they involve an inherent instability (as was already stated in the previous document) :
Organisms -- as products of organic dynamical systems -- are primarily unstable material configurations that are secondarily stabilized by flexible regulations ( In this way the causal nexus is over-formed, resulting in the nexus organicus).
All processes, such as (chemical) assimilation, dissimulation, reproduction, death and phylogenesis (by genetic mutation and natural selection) are a necessary implication of the fact that organisms are primarily unstable structures. The fact that organisms, despite their unstable nature, nevertheless exist is due to these processes, which effect recreation at several levels and, in addition, transformation (when external conditions have changed).
(Solid) crystals, on the other hand, as products of a certain class of inorganic dynamical systems, are relatively stable configurations (because they are lowest-energy configurations while organisms are not). They do not, therefore, need the above processes in order to remain existent at all. Especially they do not need natural selection, because crystals, taken generally, can (in contrast to organisms) be generated and maintained in a great variety of conditions. And because they are not products of a long evolution, they can easily be recreated after temporarily adverse conditions have returned to more favorable conditions again. But, as a result of them being so, crystals cannot evolve.
All this also applies to all other inorganic things. Therefore the primary organic functions, or, better, the analogues of primary organic functions, are scattered over the inorganic domain, i.e. they are never (found to be) together present in one and the same inorganic being (many of them in crystals, but not all of them).
The association (within a single organism) of all these primary organic functions, resulting, not just in their summation, but in a coherent and functional whole within one and the same being, is the organic NOVUM.
Liquid crystals (discussed in Part XXIX Sequel-14 ) are an example of inorganic intrinsic beings that, probably, are -- like organisms -- more or less unstable. So they will be important when comparing the Inorganic with the Organic.
The organic process, like the inorganic process, consists in the interaction of material elements. The existing spatial configuration of these elements (which configuration continually changes in the course of time) is the material form of the process (as distinguished from its energetical form, the flow and transformation of energy). Like in a machine, any material subordinated form, as one of the material parts of the organic system, performs a specific function. But in contrast to a machine, the organic material part only comes into being and is sustained by performing its (or some interim) function, which, taken generally, means that the organism already functions while still building itself up. And this is only possible when all the subordinated processes (subordinated with respect to the whole) r e g u l a t e themselves, as if they were aware of forming one single unity. It is here that wholeness determination (discussed in the previous document) comes in. It is one of the implications of the organic NOVUM.
When, in the following, we mention "typically organic features" we do not beforehand mean that these features are not found in the Inorganic. We only mean that they are especially evident in the Organic. In the ensuing 'crystal analogy' (which will, despite its name, include all Inorganic-Organic comparisons) we will see whether some or all of these features can also be found in the Inorganic.
Every organic process is embedded in, and sustained by, an inorganic dynamic context, which here means that it solely consists of inorganic (partial) processes, i.e. when we analyse an organism -- breaking it up into its constituent processes, all we find is inorganic (partial) processes. And only when these (partial) processes have themselves organized in a specific and repeatable way, resulting in a coherent whole acting like one single process, we have an organic process. Life has originated at one or several occasions on the surface of the planet. Probably only one line survived and gave rise to the diversity of life now seen to be present. Apparently, from an early point on, new life is only generated by already living entities, that is to say the inorganic world does not generate life anymore. Therefore, if life is to be continued more or less indefinitely, and given the fact of the inherent unstable nature of its representatives, life must reproduce itself again and again from still existent life. And this indeed is what has been observed. So life, once produced, basically consists in reproduction (in its broadest sense). Life is a constant replacement of parts by new parts having (nevertheless) the same nature, but also (a replacement) of whole individuals by (new) individuals of the same species. And also the species is replaced from time to time (involving longer time spans). However, it is not replaced by a species of the same nature again, but by a different species, because after longer times the environment has changed, rendering the species as a whole unstable (resulting in its transformation, or in a termination of its line of descent).
So it is constant reproduction (which, at the chemical level -- the level of metabolism -- we could call re-assimilation or repeated assimilation) that generally distinguishes living things from non-living things. And reproduction and re-assimilation must be 'deployed' in order to meet the disintegration (de-production and dissimulation) that constanly takes place.
The problem of Begging the Question, when distinguishing life from non-life.
When comparing the Inorganic with the Organic, and trying to distinguish them from each other, one easily falls into the trap of already presupposing the fundamental difference between life and non-life. This we see in HARTMANN, Philosophie der Natur, 1950, pp.519. There he first of all sets the i n d i v i d u u m as a new organic category, which appears when entering the Organic Layer from the Inorganic. But it is clear that the basic tenets of something being an individuum (and not just individual) are already found in the Inorganic : (single) crystals.
Also many other so-called exclusively organic features, allegedly not occurring in the Inorganic, that HARTMANN puts forward, are probably not as such valid.
In the ensuing crystal analogy we will discuss whether these features do or do not occur (or, can or cannot occur) in the Inorganic. Here we only mention some of them.
The organic process has (and, as HARTMANN maintains, in contrast to physical processes) its specific type of boundedness. And it has it toward both sides. It has its intrinsic beginning and end. And both, beginning and end, are implied by the system itself which passes through this process. Here, especially the boundary that marks the end is problematic. We should investigate what happens at such a boundary, and whether it is a boundary at all. Simply saying that life ends there is begging the question. Also in the Inorganic an initially generated intrinsic being can be disintegrated again, and in both cases the overall natural process continues.
HARTMANN (p.526) maintains that the natural death of an individual organism (i.e. the natural and intrinsic end of an organic process) is not determined by mere dynamic conditions of the environment, not determined by the height of some energetic fall, which exhausts itself in the process, and consequently is not intelligible in energetic terms. Although the energy stored in the food surely constitutes an energetic fall, and the organic individual that uses this energy links itself up with this fall, beginning and end of individual life are not initiated by the origination and consumption of such a fall, but also imply totally different and much more complex conditions. The presence of food does not generate the organic individual, does not initiate the organic process. And the death of the individual normally takes place much earlier than the chemical disintegration would demand it.
All this, however, is just an admission of the great complexity of the organic process. It does not testify that there exists a fundamental difference between such an organic process and a(n) (complex) inorganic process.
And begging the question about what life is, and how it differs from non-life, we can see in HARTMANN, 1950, p.527, where he says that in the inorganic process we can speak, it is true, about origin and termination, as we see it in the coming and going of process states, or in that of temporary concrete entities (things) formed by the process. But origin and termination is not -- according to HARTMANN -- as pronounced as it is in organisms.
In inorganic nature the continuance of the process is automatic and unstoppable. Here the one entity or condition turns into just another such entity or condition. And this "other" still belongs to the same order of Being (HARTMANN). Here we see that the fundamental difference of life and non-life is already presupped, instead of having it investigated. And especially with respect to the continuance of the inorganic process, matters are -- according to HARTMANN -- different in the organic. At the death of the organic individuum life does not automatically continue, when there hasn't been formed a new individual beforehand. The living system does not transform itself into another living entity when it is about to perish, but returns to the non-living, to atoms and molecules. It passes over into the negation of its Order of Being , i.e. it passes from the Organic to the Inorganic (Ibid., p.527). But this means nothing more than : Because organic processes, in their natural termination, return to the Inorganic, i.e. to another Order of Being, while inorganic processes do not (pass to another Order of Being when naturally terminated), organic processes are fundamentally different from inorganic ones. And, in short this says : Life is fundamentally different from non-life, because it is fundamentally different from non-life.
In both cases, the living and the non-living, the overall process continues. To find the difference, we must investigate that process section (of the overall process) which we associate with life, i.e. investigate what precisely is so special about it. We find it in the phenomenon of wholeness determination about which we had found out -- by analysing a dynamical system as to what it generally and essentially is -- that this type of determination does not, and cannot, exist in the Inorganic. In the Organic, however, we seem to be forced -- especially by many facts and features in embryological development -- to accept the existence of wholeness determination. But also here, i.e. in the domain of the Organic, wholeness determination is irrational. Also here we cannot reasonably separate the system-as-a-whole from its interacting elements. This irrational aspect can only be 'explained' by interpreting it as one of the implications of the organic NOVUM, or even by seeing it as (the concretum of) this NOVUM itself. And categorically it is this NOVUM that distinguishes the Organic from the Inorganic, while wholeness determination (the Whole regulating and steering all subordinated processes) phenomenologically distinguishes them. This NOVUM was called -- following HARTMANN -- the Nexus Organicus and was discussed earlier, namely in Part XXIX Sequel-24 and (more generally) in Part XXIX Sequel-25 .
Boundary (or definition) of the organic (dynamical) system.
As an important feature of organisms, categorically (according to HARTMANN, Ibid., p.532) distinguishing them from inorganic systems, is the fact that the material definition of the organic body (i.e. its concrete material boundary) does not coincide with the boundary or definition of the organic individual. The organism is, with its life functions, extended a long way into the physical world. Respiration, metabolism, search for food and self-preservation (being threatened, spontaneous defense and getting security) form an interwoven complex of actions and reactions with which the organism actively extends itself into the environment. It is irretrievably connected with a section of the real world, and is, by its partial processes of its overall life process, dependent upon it.
Here, according to HARTMANN, we see a radical difference of organic systems with respect to inorganic systems. A planetary system, continues to exist, once originated, the more undisturbedly, the more it is isolated from other entities or systems. But a flower-bearing plant, isolated from the soil, air, sunshine, rain, and even from the local insect world, not only will perish, but is no more than an abstraction.
The living individuum not only has its inner or intrinsic aspect 'in it', as is the case in all natural systems (for only fragments have it outside them), but also communicates it onto its whole domain of living (i.e. the relevant section of the environment). Surely, the organism adapts itself far-reachingly to what it encounters, but it also chooses its appropriate section of that encountered environment, and transforms it into its own domain of living. Here one finds the ultimate reason, why an animal, outside its natural domain of living -- for instance placed in a cage, or even in mimicked environments like zoo's -- often looks so miserable. Deprived of its natural domain of living (which has involved millions of years of evolution and adaptation) it just stops to be what it essentially should be according to its own nature. It is, as it were, only a mere shadow of itself. And it takes a lot of imagination and zoological knowledge to recognize in it something that indicates what it in fact was in its proper environment. The animal cannot transform the artificial environment that is forced upon it into its proper domain of living. It is then no more than a held-alive fragment of itself. Man, when caging wild animals, tears apart, without knowing, or willing to know, the essential parts of a living whole, parts, that are, as a result of a long evolution, fine-tuned with respect to each other. And matters are even worse when he trains the animal to perform tricks that oppose its nature. He is anaware of the unity of the intrinsic system that is the animal, and perhaps he lacks esteem for the animal's intrinsic wholeness, beauty and perfection.
Analogy and homology, over-forming and transformation.
In certain organisms we will encounter complex features, while primitive versions of these same features are known in other organisms. For example, the wings of Pterosaurs (fossil flying lizzards) are in fact highly specialized fore-limbs. These fore-limbs (now as non-specialized limbs) can be found in many other (non-flying) dinosaurs. Such structures we call homologous. Here the wings of the Pterosaurs are the result of trans-formation or, equivalently, of trans-forming. They are not the result of over-forming, because no appearance of a categorical NOVUM is involved here.
Only when something is the result of over-forming, and thus when a categorical NOVUM is involved, we speak of analogy. This latter term is also used in biology, but then having a slightly different meaning : The large flat external organs of the Pterosaurs are analogous to the generally transparant membrane-like external organs of many insects, because both are flight devices. So also here two disparate structures have some common (conceptual) core. In the theory of category Layers, however, we will use the terms analogy and analogous exclusively in cross-layer relations : A certain structure or dynamic pattern in a higher Layer can turn out to be an over-forming of something in a lower Layer. In this case the respective structures in the lower and higher Layers relate to each other as analogues. The same applies to their respective categories or If / Then constants.
A structure that is the result of over-forming can be further trans-formed, which means that it changes (if we go from object to object) without leaving its Layer. We then obtain a series of homologous structures. See next diagrams.
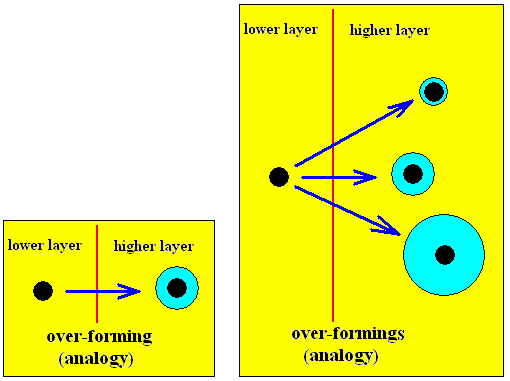
Diagram above : Over-formings
In the right-hand diagram we symbolize different over-formings of a same core, or different degrees of over-forming (of that same core).
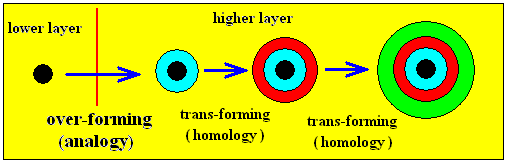
Diagram above : Over-forming and trans-forming
Organism as process
In First Part of Website we have interpreted an organism as the result of a 'totality-generating dynamical system', where a "totality" means an intrinsic being, which in turn means that it fully represents the dynamical system that has generated it.
In the inorganic domain a totality could be a (single, solid) crystal, which is generated from a melt, from vapor, or from a solution. Also here we have to do with a totality-generating dynamical system, and also here the resulting crystal can represent that dynamical system.
An organic totality (i.e. an organism) differs from a crystal by the fact that it is as such unstable, which implies that it must constantly be sustained by sub-processes that neutralize its decay. Therefore the organism necessarily is a process-structure, i.e. it is a process itself.
Although the organism in this way differs from a (solid) crystal, its process nature, i.e. its being a process, is not an exclusive feature of the organic world. Also in the inorganic world there are processes that generate definite structures or patterns that, when left alone, are unstable, and must therefore be sustained constantly by processes that counteract their decay. Said differently : when we see such structures more or less enduring, they are apparently sustained by processes that neutralize their decay.
The dynamical systems, organic or inorganic, that generate such unstable structures, are called 'dissipative systems', and the structures generated by them are called 'dissipative structures'. We treated them in First Part of Website (First Series of Documents) : Non-living Dissipative Systems (inorganic dissipative systems or structures ), and Organisms I, II, III and IV (organic dissipative systems or structures). In such systems there is a constant import of matter and energy from the environment, and a constant export of matter and entropy to the environment. In fact such systems characterize themselves by a constant renewal of substructures, be it certain chemicals or larger structures, often accompanied by growth.
Only after HARTMANN's last work (1950) much and important research into such systems (special inorganic chemical reactions, computer simulations and mathematics) has been carried out, largely by Ilya PRIGOGINE. So, not knowing the results of this research (or otherwise differently interpreting them), HARTMANN, considers the process nature of organisms, and especially the interwoveness of form and function, as exclusively organic features. But, as far as I am able to judge (from simulations) we do not have to do here with totally new categories, determining these features, but at most with over-formed categories.
Wholeness Determination as representing the organic NOVUM, revisited.
The NOVUM in the organismic realm involves, or is, wholeness determination : In addition to the determination exerted by (any) one element on other elements of a dynamical system (i.e. the interaction of those elements), where, consequently, each element has its share of determining the Whole, there is (in organisms) also a determination, exerted by the Whole (which here is, first of all, the individual organism), of every element of that Whole (i.e. that Whole influences its interacting elements).
This latter is in fact irrational, because, basically, the Whole is nothing else than the total of its (interacting) elements, which means, that apart from the elements there is no Whole. The 'Whole' is its (interacting) elements. The 'Whole' is nothing more than an abstract entity. Therefore -- as such -- it cannot exert any physical influence whatsoever.
However, in the case of an o r g a n i s m, we -- on the basis of countless observations pointing to its holistic nature -- postulate that a c o n c e t e Whole exists : Any individual organism is a true (and thus concrete) Whole, not a mere abstract whole. It is a Whole that is more than the sum of its (interacting) elements or parts. It is a real entity which as such can exert influence on its elements. And as such it is the organic NOVUM. And in whatever way we will (manage to) describe it, it will continue to contain an irrational aspect.
So in organisms the Whole (representing a single individual organism) is a concrete entity. And when this entity, albeit partly, determines the interactions of its elements, i.e. organizes these interactions, we have wholeness determination.
Life originating from the Inorganic, means that the Whole (i.e. an originated organic individual) becomes concrete (while wholes are abstract in the Inorganic). This initial concrete Whole, once originated in the form of some primordial organism, will subsequently be copied and transformed in the course of time. But, because of its inherent irrationality, we cannot even speculate about its (ultimate) origin. It just as such emerges.
Is it possible -- albeit speculatively and incompletely -- to describe such a concrete whole? The assumption of such a Whole in biology is equivalent to the claim of many researchers that life cannot be totally reduced to physics and chemistry. Organisms are, according to them, 'holistic'. We can read this in the books of writers such as Steven ROSE, Lifelines, 1997, and Brian GOODWIN, How the Leopard Changed Its Spots, 1994, 2001. However, they did not (dare to) reflect on what exactly this holism means. Instead some such authors tend to ridicule some others who did. Other writers, speaking about 'non-reductionism' (for example, Jack COHEN & Ian STEWART, The Collapse of Chaos, 1994) pitifully confused (1) methodology (and approach) and (2) ontology (i.e. (1) how, and how far, science is actually able to penetrate the mystery of complex systems, and (2) what this mystery actually is, or, in other words, (1) how and how complete can we know Nature, and (2) how Nature really is in itself, independent of her being partially or wholly known).
There are, however, writers who took the trouble to investigate the consequences of the alleged 'holism' (of course in an ontological sense) in organisms. They indeed recognized the necessary introduction of an irrational aspect, when accepting (ontological) holism (and also for them 'holism' is equivalent to wholeness determination). Among them are David BOHM (Wholeness and the Implicate Order, 1980, 1988) and Rupert SHELDRAKE (A New Science of Life, 1981, 1987, The Presence of the Past, 1988).
I think that in SHELDRAKE we can find ideas for a further description of our 'concrete whole'. However, SHELDRAKE extends the assumed phenomenon of a concrete Whole over all of reality, i.e. including the inorganic world. Although this could be true, we here assume that the concrete Whole only appears after the transition from the Inorganic Layer to the Organic and then reappears in the higher Layers (Psychic and Super-psychic), while it is totally absent in the Inorganic.
And now indeed we have our organisms such that they are not totally reducible to inorganic entities.
So let us now, following SHELDRAKE ( NOTE 1 ), describe these concrete wholes (which SHELDRAKE calls morphogenetic fields or morphic fields for short), as they are present in the organic world, representing the organic categorical NOVUM as a new nexus category (nexus organicus), a nexus that consists in the fact that such a (concrete) whole co-determines the interactions of the elements of this whole (wholeness determination), resulting in a specific organization, which as such is Life. And the reason why we investigate this NOVUM further (as far as it goes) is because we ask ourselves in what way a concrete whole co-determines the interactions of its elements. A general answer to this question is that this determination by the concrete whole consists in selecting causal factors in order to let a certain structure to be generated (by causality!). We do not assume that the determination exerted by the concrete whole fits in where the physico-chemical processes are themselves intrinsically indeterminate (because these processes are not indeterminate in themselves, only we cannot always predict their outcomes). By themselves, these processes can result in different results on different occasions, because of different prevailing conditions. Wholeness determination stabilizes these conditions, resulting in a definite and repeatable outcome. And to delve still deeper in the way wholeness determination goes its business, we follow SHELDRAKE
and his concept of morphic resonance.
The Hartmannian theory of the organic NOVUM, as well as SHELDRAKE's theory of the morphogenetic field (which we can here identify with the concrete whole) oppose the so-called mechanistic (s.l.) theory. This latter theory asserts that in principle all organic phenomena can totally be explained by the known laws of physics (and chemistry). Let us cite SHELDRAKE on this point (A New Science of Life, 1981, p.45--47 of the 1987 Paladin Edition [ comments between square brackets ] :
However, the mechanistic theory does not merely suggest that these and other physical processes [ crystallization processes, surface tension processes, processes involving colloids, and, further, patterns arising from statistically random fluctuations ] play a part in [ organic ] morphogenesis. It asserts that morphogenesis is entirely explicable in terms of physics. What does this mean? If everything observable is d e f i n e d as being physically explicable in principle, just because it happens [ i.e when 'observable' means the same as 'physically explicable' ] , then it must be so by definition. But this does not necessarily mean it can be explained in terms of the k n o w n laws of physics. In relation to biological morphogenesis this explanation could be said to have been achieved if a biologist who was supplied with the entire sequence of bases in the DNA of an organism and a detailed description of the physico-chemical state of the fertilized egg, and of the environment in which it developed, could p r e d i c t in terms of the fundamental laws of physics (e.g. quantum field theory, the equations of electromagnetism, the second law of thermodynamics, etc.) first, the three-dimensional structure of all the proteins the organism would make. Secondly, the enzymic and other properties of these proteins. Thirdly, the organism's entire metabolism. Fourthly, the nature and consequences of all the types of positional information that would arise during its development. Fifthly, the structure of its cells, tissues and organs and the form of the organism as a whole. And, finally, in the case of an animal, its instinctive behaviour. If all these predictions could be made successfully, and if, in addition, the course of processes of regulation and regeneration could also be predicted a priori, this would indeed be a conclusive demonstration that living organisms are fully explicable in terms of the known laws of physics. But, of course, nothing of the sort can be done at present. So there is no way of demonstrating that such an explanation is possible. It might not be.In order to account for phenomena like regulation and regeneration in organisms, one has put forward the idea of fields that are somehow associated with developing organisms. However, the nature of these fields remained obscure. SHELDRAKE, 1981, came up with his version of a field theory, namely his 'hypothesis of formative causation'. In his second book about this subject ( The Presence of the Past, 1988, p.108/9), he wrote :
Thus if the mechanistic theory states that all the phenomena of morphogenesis are capable in principle of being explained in terms of the known laws of physics, it might well be wrong : so little is understood at present that there seem to be no good grounds for a firm belief in the adequacy of the known laws to explain all the phenomena. But at any rate this is a testable theory. It could be refuted by the discovery of a new [ and relevant to our problem ] law of physics. If on the other hand the mechanistic theory states that living organisms obey both known and unknown laws of nature, then it would be irrefutable. It would simply be a general statement of faith in the possibility of explanation. It would not be opposed to organicism and vitalism. It would include them.
In practice, the mechanistic theory of life is not treated as a rigorously defined, refutable scientific theory [ better, a theory of the theory of science, or, epistemological theory ] . Rather, it serves to provide a justification for the conservative method of working within the established framework of thought provided by existing physics and chemistry. Although it is usually understood to mean that living organisms are in principle fully explicable in terms of the known laws of physics, if a new law of physics were to be discovered, and thus became known, the mechanistic theory could easily be modified to include it. Whether this modified theory of life were to be called mechanistic or not would only be a matter of definition.
When so little is understood about the phenomena of morphogenesis and behaviour, the possibility can by no means be ruled out that at least some of them depend on a causal [ better : determining ] factor as yet unrecognized by physics [ and even one that is so because it is a non-physical factor such as wholeness determination ]. In the mechanistic approach, this question is simply put aside. Nevertheless it remains entirely open.
[comments between square brackets]Using these ideas of SHELDRAKE's to (further) characterize the organic NOVUM, we get the following :
What is new in the hypothesis of formative causation is the idea that the structure of these fields is not determined by either transcendent Ideas [ In the sense of Plato ] or timeless mathematical formulae, but rather results from the actual forms of previous similar organisms. In other words, the structure of the fields depends on what has happened before. Thus, for example, the morphogenetic fields of the foxglove species are shaped by influences from previously existing foxgloves. They represent a kind of pooled or collective memory of the species. Each member of the species is moulded by these species fields, and in turn contributes to them, influencing future members of the species [ SHELDRAKE, speaks here of "the morphogenetic fields of the foxglove species", because he recognizes a field for each level of complexity within a single organism (for instance a field for the structure of its cells, then a higher-level field for the structure of its organs, etc., up to the field that determines the organism as a whole). We do not think that this is a fruitful idea (at least not within our conception of a morphogenetic field as representing wholeness determination, exclusively present in organisms, and referring to the individual organism as a whole). So we prefer "the morphogenetic field of the foxglove species". And such a field represents a "kind of pooled or collective memory of the species".].
How could such a memory possibly work? The hypothesis of formative causation postulates that it depends on a kind of resonance, called morphic resonance. Morphic resonance takes place on the basis of similarity. The more similar an organism is to previous organisms, the greater their influence on it by morphic resonance. And the more such organisms there have been, the more powerful their cumulative influence. Thus a developing foxglove seedling, for example, is subject to morphic resonance from countless foxgloves that came before, and this resonance shapes and stabilizes its morphogenetic fields [ Again, we prefer field ].
Morphic resonance differs from the kinds of resonance already known to science, such as acoustic resonance (as in the sympathetic vibration of stretched strings), electro-magnetic resonance (as in the tuning of a radio set to a transmission at a particular frequency), electron-spin resonance, and nuclear-magnetic resonance. Unlike these kinds of resonance, morphic resonance does not involve a transfer of energy from one system to another, but rather a non-energetic transfer of information. However, morphic resonance does resemble the known kinds of resonance in that it takes place on the basis of rhythmic patterns of activity [ Indeed Reality can be thought of as patterns of vibrations ].
[ . . . ]
Because all past members of a species influence these fields, their influence is cumilative : it increases as the total number of members of the species grows. Since these past organisms are similar to each other rather than identical, when a subsequent organism comes under their collective influence, its morphogenetic fields [ Once again, we prefer field ] are not sharply defined, but consist of a composite of previous similar forms. This process is analogous to composite photography, in which "average" pictures are produced by superimposing a number of similar images [...]. Morphogenetic fields are "probability structures", in which the influence of the most common past types combines to increase the probability that such types will occur again.
The Concrete Whole of an organism as its organizing field.
A developing organic individual (say, an embryo) is itself not yet the final whole. During individual development its (complete and final) whole is as such (only) present in similar (but fully developed) p a s t organisms. This (final) whole now becomes the (morphological and physiological) Whole of the (presently) developing organism, i.e. it becomes inherited by, and immanent in, the developing organism by morphic resonance between the latter and the (morphological and physiological) whole that is represented by all past similar organisms. It is now the developing organism's concrete (final) Whole. As a field of information (while not being energetic) it issues a determination (wholeness determination) that organizes the elements and dynamical subsystems of the developing organism by way of selecting and organizing available causal factors.
This organizing determination is -- as has been explained earlier -- itself irrational (i.e. it contains an irrational aspect), but once this organizing determination is accomplished (i.e. has taken place), we have (obtained) just an 'ordinary' physico-chemical dynamical system, which now automatically and necessarily, and in a purely physico-chemical way, produces its generic form. And this form we indeed call (following GOODWIN, in How the Leopard Changed Its Spots, 1994) 'generic form', because it will be reached equally well (by the dynamical system) from a range of more or less different initial conditions and parameters (See also below), resulting in the fact that it appears as a common trait in a group of related organisms. So this stable form is not the direct result of morphic resonance, but (the direct result) of the physico-chemical dynamical system, itself resulting from the organizing wholeness determination.
Because the concrete whole of the organism, as well as its interacting elements, are concrete, not only this concrete whole can act on the interacting elements, but the latter can also act on that whole. Said equivalently : The field not only acts on the (developing) organism, but the organism also acts on its field. And this implies that the field can, and will, e v o l v e, which generally means that it becomes (still) more complex.
SHELDRAKE tends to 'physicalize' his morphogenetic fields as much as possible. And this is understandable, because he wants to present his hypothesis an an experimentally testable theory of natural science. We, on the other hand, use the concept of morphogenetic fields to characterize the organic NOVUM, and this theory of the NOVUM is not a theory of natural science. It is rather an ontological (or, if one prefers to express oneself, metaphysical) theory (which allows for an irrational aspect to be involved).
Because we consider wholeness determination as just a (albeit irrational) type of determination among other types (which are rational), and in this case a determination that only consists in selecting and organizing prevailing causal factors, we should not, in our theory, let the field (which is the concrete whole, exerting wholeness determination) determine too much, that is to say, we should let its scope be limited such that (1) its determinative role within a single individual entity or being is limited to a certain aspect of the overall determination of that being, and (2) its role is confined to organisms, and excluded in inorganic beings. As has been said, it results in the formation of an 'ordinary' but complex overall dynamical system, which as such is (now) purely physico-chemical, and which in turn generates, among other things, a generic form, which is an essential part of the overall form (including structure) of the corresponding organic species (group). In our First Part of Website we have called this generic form, but then as it is subsequently and additionally (further) determined to represent a particular species (of the group), the phenotypical intrinsic expression of the organism's ' Essence', where the latter is the organism's intrinsic genotypical aspect or dynamical law.
The g e n e s of a given organism are part of this dynamical law : They are ( We here follow GOODWIN ) specific parameter settings of this law (where the latter is the law of the overall dynamical system which in fact (categorically) constitutes that (given) organism).
The configuration and organization of causal factors (i.e. wholeness determination) and the product that is generated by the dynamical system resulting from that configuration (i.e. resulting from that wholeness determination), can be illustrated by the following analogy given by SHELDRAKE in his A new science of life, 1981, p.125 :
[comments between square brackets]
Consider the following analogy. The music which comes out of the loudspeaker of a radio set depends b o t h on the material structures of the set and the energy which powers it a n d on the transmission to which the set is tuned. The music can of course be affected by changes in the wiring, transistors, condensers, etc., and it ceases when the battery is removed. Someone who knew nothing about the transmission of invisible, intangible and inaudible vibrations through the electromagnetic field might therefore conclude that it could be explained entirely in terms of the components of the radio, the way in which they were arranged, and the energy on which their functioning depended. If he ever considered the possibility that anything entered from outside, he would dismiss it when he discovered that the set weighed the same switched on and switched off. He would therefore have to suppose that the rhythmic and harmonic patterns of the music arose within the set as a result of immensely complicated interactions among its parts. After careful study and analysis of the set, he might even be able to make a replica of it which produced exactly the same sounds as the original, and would probably regard this result as a striking proof of his theory. But in spite of his achievement, he would remain completely unaware that in reality the music originated in a broadcasting studio hundreds of miles away.
In terms of the hypothesis of formative causation, the 'transmission' would come from previous similar systems, and its 'reception' would depend on the detailed structure and organization of the receiving system. As in a radio set, two types of change in the organization of the 'receiver' would have significant effects. First, changes in the 'tuning' of the system could lead to the reception of quite different 'transmissions' : just as a radio set can be tuned to different radio stations, so a developing system can be 'tuned' to different morphogenetic fields. Second, just as changes within a radio set tuned to a particular station can lead to modifications and distortions of the music coming out of the loudspeaker, so changes within a system developing under the influence of a particular morphogenetic field can lead to various modifications and distortions of the final form.
Thus in developing organisms both environmental and genetic factors could affect morphogenesis in two different ways : either by changing the 'tuning' of morphogenetic germs [ initial stage of organic development that associates itself with the morphogenetic field of the whole organism ], or by changing the usual pathways of morphogenesis in such a way that variants of the normal final forms are produced.
The role of the genes in morphogenesis.
We have found out that an organism is the product of, and at the same time represents, a very complex dynamical system. This dynamical system is forged by wholeness determination where the latter has selected and organized causal factors. The dynamical law of this system is the organism's Essence (as was found out in First Part of Website) or genotypical aspect or domain, while the product of this dynamical system is the phenotypic aspect or domain of the organism. The dynamical law contains a great many parameters, which are magnitudes that remain constant during one run of the dynamical system ( Here "one run" of the system means the complete individual development of an organism). There are good ground to assume that the organism's genes do not directly create or affect macroscopic features of the organism (all what genes do is code for specific proteins and their orderly production in the right amounts at the right place and at the right moment). Therefore we assume, following GOODWIN, 1994, How the Leopard Changed Its Spots, that the organism's genes represent parameter settings of the overall dynamical system that generates and maintains the organism. Some other parameters are set by the environment.
An egg cell of an organism can be called an 'exitable medium' which means that it will develop patterns after it has been stimulated in some way (something analogous we see in the case of the famous Belousov-Zhabotinsky Reaction, which is an inorganic chemical reaction system that develops spatial patterns). So the exitable medium is in fact the initial condition of a pattern-generating dynamical system.
We can indicate diagrammatically the role of the genes of an organism in its morphogenesis (See Diagrams below). These genes (together with certain environmental factors) define the parameters of the dynamical law of the dynamical system generating and maintaining the organism. Now we can concentrate on an intrinsically robust generic form or structure that is generated by the system and is common to a group of related organic species. The diagram indicates all species that have this generic structure in common, and concentrates on three of them (indicated by colors : light blue, red and green). Let us consider the space of parameters (parameter settings, genes). The total set of parameter settings (genes) of one individual of a given organic species defines one point in a multidimensional parameter space : If we have, say, 100 parameters (i.e. 100 parameter settings, represented by values), then each value is a point on one particular dimension of the (multidimensional) parameter space. So all 100 values define a point in this space. In our diagram we have very much simplified all this, resulting in a two-dimensional parameter space, represented in the diagram by a plane.
This parameter space, together with a given dynamical law, defines a particular organic exitable medium, which can be identified with a certain taxonomic group -- let us call it A -- of related organic species. This (particular) exitable medium only gives viable life-cycles when the parameter settings (genes) fall within the (large) oval area. Otherwise the life-cycle is not viable, or is a transient leading to another exitable medium. Each point in the oval area of the parameter space can give rise to a morphogenetic trajectory of a species of A. All parameter values that lie within the oval area produce trajectories that lead to (a whole organism possessing) the generic form, described as the narrow neck (small oval in diagram) of the cone through which these trajectories pass. This form represents a structure that is typical of a whole group of species, describing a high-level taxonomic characteristic that all the members of the group A share. Often such a generic form or structure is absent in the adult form of such a species, but then its morphogenesis passes through it as a juvenile, and the generic form undergoes secondary modification later, or even disappears. This radiation of secondary forms is shown in the diagrams as the diverging arrows leading to the different species labeled light blue, red and green. These species originate from parameter domains labeled with the same colors. They all lie within the oval area of the parameter space.
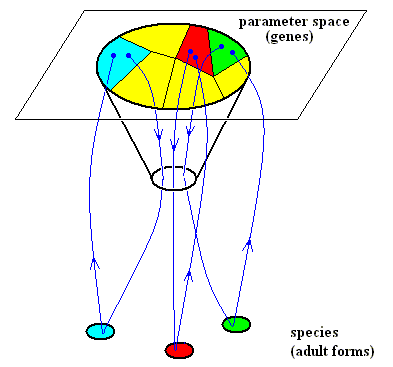
Diagram above : The role of genes in the morphogenesis of organic species of a given taxonomic group. Each point of the parameter space is a set of parameter settings and corresponds to the genetic constitution of a given organism (After GOODWIN, 1994). See this diagram again, now with some more indications :
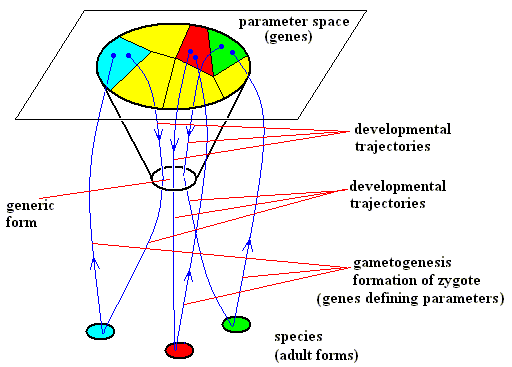
Diagram above : The role of genes in the morphogenesis of organic species of a given taxonomic group (See also text above and below). Morphogenetic trajectories, gametogenesis and zygote formation (reshuffling and possible mutation of genes) and generic form, explicitly indicated. After GOODWIN, 1994
During reproduction, each species produces gametes and then zygotes with (reshuffled or mutated) genes defining parameters that specify what morphogenetic trajectory the zygote will follow. This is indicated by the upward trajectories. This process completes the life-cycle. Each species has a closed loop, with a range of tolerance (indicated by a color) for genetic variation that defines the viability limits of the species. The whole group lies within a set of tolerance parameters, indicated by the large oval in the diagram. Of course, random variations can result in trajectories that fall outside the set (for example, a trajectory starting from a point within the oval area, but returning to a point outside it), giving either non-viable organisms or transients leading to another exitable medium (representing another taxonomic group) ( See for all this, GOODWIN, B., How the Leopard Changed Its Spots, 1994, p.112/3 of the 2001 Princeton Science Library Edition).
Historical and Rational Taxonomy.
All documents in our website (four Parts) are about p a t t e r n s. Where we encounter them in Nature they generally are stable patterns, such as crystals and organisms, but also atoms, molecules, stars and other astronomical objects. Even tornados are such stable patterns.
With "stable pattern" we mean dynamically stable patterns. Such a pattern is generated by a dynamical system. Its stability means that also when the initial condition of the dynamical system is more or less different, that same pattern will be generated nevertheless. It means further, that it will also be generated when one or more parameters (magnitudes that remain constant during a run of the system) are set more or less differently from those of a previous run of the system. Also the subsequent endurance or persistence of such a pattern, once it has been generated, must, in order for it to be called stable, be possible within a range of external conditions, rather than only one particular condition.
In physics we have, among other things, to do with several kinds of atoms. They are patterns that can persist under a range of conditions. Their properties define certain possibilities of connection (chemical bonding) resulting in relatively stable patterns called molecules, which are studied in chemistry and are called compounds when present in bulk quantities. Atoms, but also some (types of) molecules can combine to form crystals, which are in fact giant molecules with a periodic structure and without a strictly defined size. Also these crystals are stable patterns. Physics, chemistry, crystallography and mineralogy are trying to classify these patterns, resulting in classificatory systems.
But also organisms are relatively stable patterns. But in order to endure they must actively maintain themselves, by constantly renewing their components, and in the end, by reproducing themselves, and finally by transforming themselves in order to adapt to changed external conditions (biotic and abiotic). This is because an organism as such, i.e. totally in and by itself, is intrinsically an unstable pattern. Only this pattern together with its maintaining activities is the true organism. And only then it is a relatively stable pattern. Such a pattern, that is to say an organic pattern (i.e. an organism) can be generated equally well from more or less different initial conditions, its morphogenesis has the capacity of regulation (resume its course after a perturbation has taken place, temporarily diverting the system's trajectory), and this same pattern will be produced when parameters are slightly changed. Such an organic pattern, in conjunction with these inherent capacities, is, like atoms, molecules, crystals, etc., an objective intrinsic entity which intrinsically is what it is. When conditions are right it will appear, when they're not it will not appear. This status of objectivity is precisely the same as in the case of atoms, molecules, crystals, etc. Also they will only appear when certain conditions are right. For (heavier) atoms to be generated extremely high temperatures are needed. Once formed, they can maintain themselves in a broad range of moderate temperatures. The generation and persistence of molecules generally depend on relatively mild conditions. Where very high temperatures prevail no molecules can exist. The same goes for crystals. For organisms even milder conditions are needed for their generation and persistence.
So organisms can be considered as relatively stable patterns, and thus objective and intrinsic entities, alongside those that are represented by atoms, molecules and crystals.
But how then does biological evolution fit into this picture of stable patterns?
What in fact does the theory of evolution investigate? Well, the theory of evolution of organic species investigates what the history of Life on the planet Earth has been so far, and why it has been so. The theory tries to answer these questions by studying changing environments, changing genes and natural selection : Under changed conditions (biotic and abiotic) those variants within a species that are best fit to live in the changed environment and having the properties, that make them so fit, genetically fixed, will, on average produce more offspring, resulting in them to outnumber the others, and finally to replace them. Eventually this will lead to a new species, well adapted to the changed environment. Generally the change of the environment is more or less local, meaning that, in addition to areas where the environment has changed, there still are other areas where the environment is in its original state. And there the species is not forced to transform. All this results in the splitting-up of some one species into two species, of which one strongly resembles the original species, while the other has become different (in such a degree that they cannot interbreed anymore). While the Earth's history proceeds, and environments change, the splitting-up of species continues, resulting in the family tree of Life (genealogical tree of life forms).
It is evident that this genealogical tree precisely reflects the history of Life on Earth. It is also evident that this history, and therefore, the sequence of all the emerging organic species is one of the effects of what has happened on Earth, i.e. the history of Life reflects the sequence of environmental changes on Earth (that include climates, land-sea relations, but also the presence of organisms themselves, resulting in evolving ecologies).
Indeed, all this is what the theory of evolution investigates. And its results are expressed in the form of genealogical trees. But a genuine t a x o n o m y of life forms should not be based on this, because the structure of these trees is strongly dependent on the (relevant) events that happened to take place on this particular planet. A genuine taxonomy of life forms should be independent of some given planet or any particular location in the Universe.
A true taxonomy should be objective, intrinsic and universal.
When we describe and classify, say, c r y s t a l s, according to what they are in themselves, we do this by indicating their chemical composition and their space group. The latter indicates the type of crystal lattice plus the symmetry of chemical motifs that occupy the points of the lattice. Together with the knowledge of the actual chemical composition of these motifs the complete intrinsic structure of the crystal is then known. When we do this for as many crystals (naturally- and laboratory-generated) as possible, we are able to set up a taxonomy of crystal species, based on their intrinsic structure. And it is clear that such a taxonomy is totally independent of a particular planet, or of a particular sequence of environmental changes during which these crystals have come into being. Or, in other words, it is totally independent of the history of crystals (minerals) on Earth, it is an objective, intrinsic and universal taxonomy of crystals. In the same way we want the taxonomy of organic species to be.
This means that also organisms must be classified according to their intrinsic structure, and not according to their history that took place on a particular planet ( We can assume that the Universe harbors much more than one planet on which life exists). This intrinsic structure is the stable product of the organism's dynamical system as the latter is set up by the organism's wholeness determination. In short, i.e. (now) already presupposing the organic NOVUM, we can say that an organismic species is a relatively stable pattern, generated by a dynamical system. We can now classify organic species according to their relations of dynamic kinship, which means that the more the dynamical systems (generating the structure of the members of given species) are similar to each other, the more the corresponding species belong together. And this is something quite different from recency of common ancestor, which is the criterion of historical (ancestral) kinship. See next diagram.
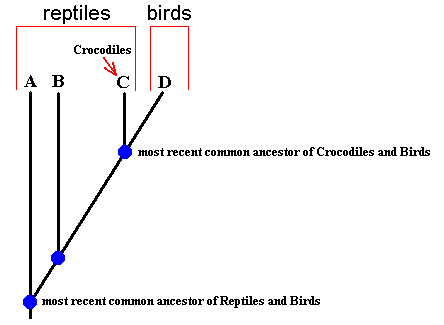
Diagram above : Sketch of the evolution of the Sauropsida (Reptiles + Birds), given in the form of a genealogical tree. A + B + C = Reptiles. D = Birds. C = Crocodiles. Conventionally, taxonomical groupings (taxa) are formed on the basis of ancestral (historical) kinship relations. It is assumed (for the sake of argument) that Birds and Crocodiles have a common ancestor. This implies that C + D is a monophyletic group, and therefore a true historical phylogenetic taxon, despite the fact that C and D (Crocodiles and Birds) look quite different with respect to each other. Because the common ancestor of Crocodiles and Birds is more recent (in geological history) than that of Crocodiles and any other member of the ' Reptiles', Crocodiles are -- historically -- more closely allied to Birds than they are to the rest of the ' Reptiles'.
The whole group of Reptiles (A + B + C) is not a genuine historical phylogenetic taxon, but only a typological grouping, while the Sauropsida as a group (A + B + C + D) do represent such a taxon, and also the Birds (and, of course, also the Crocodiles).
Here we see that on Earth a series of species bifurcations has taken place as a result of environmental changes (biotic and abiotic), which were such that Birds only relatively late evolved from Reptiles, at a time when there existed already many different types of the latter. Some one particular species of Reptile then bifurcated into two branches, of which one led to -- what we now call -- Crocodiles, while the other to Birds.
Having finished all preparations, the next document will now take up the crystal analogy.
This was in the year 2005. . . , but because we realized that in fact all the elements of the crystal-analogy (crystals-organisms) were already present in the present and foregoing series of documents, we had decided to postpone the presentatation, in the next document, of this crystal analogy because it would be no more than a summary of those foregoing documents. Meanwhile, in the year 2010, we have -- in addition to all what we had already found in the foregoing documents -- found new elements, negative and positive, contributing to the development of the crystal-analogy (we have found them in Sixth Part of Website). We will discuss these elements in the next document and present a final crystal-analogy (based on these findings including most of those already discussed in the foregoing documents).
In the document(s) after the one concerned with the "final" crystal analogy (coming up next) we leave the investigations into the latter, and, based on the results of these investigations, now accept this analogy as a general context for the previous and ensuing work. This ensuing work will be a continuation of our investigation into the promorphs of two-dimensional crystals, but now explicitly involving their internal symmetry (plane group symmetry). In addition, the crystal analogy could point (if it allows to be so interpreted) to two different Layers of Being, that are either distinguished from each other by a (mere) jump in physical complexity (and nothing else), or by some really fundamental ontological NOVUM, resulting in the organisms to be elevated or over-formed (non-equilibrium) crystals.
e-mail :
To continue click HERE for further study of the Theory of Layers (Crystal Analogy), Part XXIX Sequel-35.