(Takes a little while loading the images)
e-mail :

Sequel-12 to the Summary and Evaluation (of the documents XVI -- XXVIII concerning the promorphology of complex-shaped two-dimensional crystals).
This document (Part XXIX Sequel-12) further elaborates on the analogy between crystals and organisms.
Sequel to Biological Morphogenesis and Organic Lattices
Further notes about the compatibility and necessity of an organic lattice structure within the developing organism.
(Here we continue to discuss relevant features of embryology, relying heavily on HADORN & WEHNER, 1977 (complete reference in previous document)).
Many organizational processes in morphogenesis are based on local more or less straightforward attraction and repulsion forces between cells as a result of changing cell-surface properties : probably specific chemical compounds present in the cell membrane or present as a thin film on the membrane (HADORN & WEHNER, p.159--161). So these cell affinities are local organization factors in morphogenesis (comparable with demixing phenomena in mixed crystals). However, the change of surface properties takes place with respect to several particular different cell groups, and the developing germ somehow determines which cells must change their surface properties. This then is the global pre-ordering, which is then automatically followed by congregation and separation of cells, resulting in a definite pattern, and this congregation and separation is the local process of ordering mentioned above.
HADORN and WEHNER, p.159--161, describe all this as follows :
The generation of definite structures in blastemas [ = cell groups consisting of not yet differentiated cells], as for example the splitting off of the chorda primordium from the surrounding mesoderm that constitutes the roof of the primitive gut (See the Figure in the previous document ), necessitates a change of the c o n t a c t b e h a v i o r of the cells. One can experimentally demonstrate that an initially positive affinity, that, during gastrulation guaranteed the connection of the cells, flips over to a negative affinity [ This process must, of course, itself be patterned], leading up to a separation. What in fact, as a result of the embryonic differentiation process, changes, are the surface properties of the cells, probably specific compounds that are present in the membrane, or covering the latter as a thin film [...]. In this way the cells are capable to distinguish identical (isotypic) tissue from alien (heterotypic) tissue.
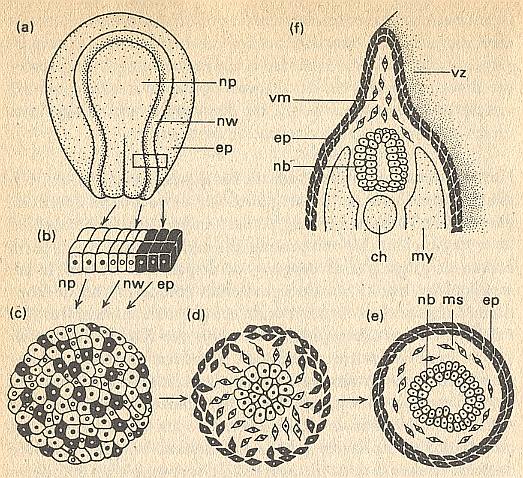
In what way c e l l a f f i n i t i e s can function as organization factors in organogenesis, is made clear in the next Figure.
Figure above : Morphogenesis in a collection of cells as a function of cell affinities.
(a) Extraction of three cell types : ep = epidermis, np = neural plate, nw = neural embankments, from the neurula of an amphibian.
(b) Before dissociation.
(c) Re-aggregation in the explantate.
(d) Beginning demixing.
(e) Differentiated end stage with neural tube (nb), mesenchym (ms) and epidermis (ep).
(f) The region of the larva that corresponds with the explantate. ch = chorda. my = myotome. vm = mesenchym of fin edge. vz = fin edge.
(After TOWNES, and HOLTFRETER)A part of the ectoderm from a neurula of an amphibian is isolated [See Figure above] and dissociated into a collection of individual cells in an alkaline solution (or by way of trypsine). Cells, which already have undergone a certain differentiation, will now become randomly mixed. In a normal culture medium they together form a disordered spherical cell mass, which one can, as explantate (Figure above, (c)), breed further. But then, however, one sees that a change is taking place in the chaotic cell mass. The e p i d e r m i s c e l l s move to the periphery and they only come to rest when they find themselves unified in contact with identically differentiated cells, in a surface layer (above Figure, d, e). In the same way the p r e s u m p t i v e n e r v e c e l l s [emphasis mine] keep moving till they have concentrated in the center of the cell mass, after which they together form a piece of the spinal cord. They avoid all contact with the epidermis cells. The third type of cells, to which also belong the cells of the n e u r a l l e d g e [...], manoeuvres itself between epidermis and neural tube as mesenchym. The result of all this, is a structure showing correspondence with the positioning of the organs in a larva [of an amphibian] (above Figure, f).
As one can see in (e) of the above Figure, cell affinities do not really create a genuine pattern. The resulting structure is just an expression of attraction and repulsion of cells. In ordinary crystallization this is also the case (here with atoms or atomic groups). But the intricate macroscopic patterns that result from non-equilibrium crystallization are not just the result of attraction and repulsion, but the result of incomplete growth, in turn resulting from rapid aggregation of elements. In the same way the true organization (i.e. organized distribution) of the cells that have already been differentiated earlier as to their surface properties, leads to a structure like in (f) of the above Figure. And it is this genuine organization (which makes use of the difference in cell-surface properties) that asks for an explanation. And this explanation will necessarily involve global factors, determining the distribution of cells according to some overall implicit design or pre-design. The latter could be provided by the fast-growth result of a pre-existing organic lattice.
When one mixes cells from skin-ectoderm, mesoderm and endoderm of an early gastrulation in this way, one obtains in principle the same result. The ectoderm cells are being moved to the periphery to form an epidermis there. The mesoderm cells unite to form an intermediate layer as muscle or chorda primordium, and the endoderm cells concentrate in the center to form a gut primordium. Thanks to such affinity-reactions a sponge, which one presses through a finely meshed sieve, is able to restore its original organization from the chaotic cell-mass that is the result of such a sieving treatment. Also the gastrula of a sea-urchin is able to reorganize after dissociation.Apart from these short-range processes, there are also chemotaxic long-range effects, that result in the arrangement of cells into a pattern (HADORN & WEHNER, p.162). This probably takes place by (concentration) gradients emanating from certain cells, which are target cells when chemotaxis is positive. A negative chemotaxis results in the involved cells to stay away from each other (HADORN & WEHNER, p.162). Our hypothetical pre-lattice points (chemical units, that are in the process of representing these points) are assumed to work in a comparable manner.
The m e c h a n i s m s that are fundamental for the morphogenetic cell migrations, are still insufficiently known. It is, however, without qualification, certain that in embryonic cells a tendency to migrate appears as soon as they are isolated from each other or being bred together with certain heterotypic neighbor cells. If they come into contact with other cells, then, dependending on the nature of these partners, the migratory movement is enhanced, weakened or halted completely. An encounter with isotypic cells generally results in halting. In this c o n t a c t-b r a k e the undulating motion of the membrane [...] stops. In this way the cells can come together resulting in a stable cell connection. In the case of c a n c e r c e l l s such a contact-brake is probably absent, which can lead to uncontrolled growth and to metastases.
The mutual attractive properties are not species-specific [yet another indication that they are not the very organizing factors] , as mixtures of embryonic cells, from mouse and chicken demonstrate. The cell partners, though coming from very different animal Classes, together form, for example, chimeric kidney channels or fragments of cartilage.
[. . . ]
Positive cell affinities not only result in the formation of homogeneous cell groups (for instance epidermis). Often they also play a role between heterotypic cells. In this way endoderm and mesoderm together constitite the liver [...]. Even ectoderm can react positively to endoderm, which, among other things, guarantees the contact between oral aperture and cephalic gut [...].
The contact behavior of the cells is based upon a hereditarily programmed developmental dynamics. The latter forms the basis of the early morphogenesis as well as of the subsequent organogenesis.
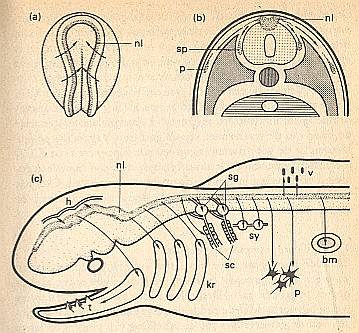
Figure above : Neural ledge (nl) of an amphibian (enlargement of (c), see LINK below).
(a) Neurula (arrows indicate the direction in which the neural 'embankments' move).
(b) Beginning transposition (migration) of the cells (arrows) after the neural tube has detached itself from the skin ectoderm. p = pigment cells. sp = spinal ganglion.
Cells migrate (arrows, see also (c)) from the neural ledge (nl) to (the regions of developing)
brain skin (h), to (the regions of developing) dentine germs of teeth (t), to (the regions of developing) cartilage of the visceral skeleton (kr), further, to the cells of the sheath of Schwann (sc), to the spinal ganglia (sp, sg), to sympatic ganglia (sy), to the pigment cells (p), to the marrow of the adrenal gland (bm), and to the mesenchym of the fin edge.
(c) Diagram of the formed organs and tissues.
bm = marrow of adrenal gland. h = brain skin. kr = cartilage of the visceral skeleton. p = pigment cells. sc = cells of the sheath of Schwann. sg = spinal ganglia. sy = sympatic ganglia. t = dentine germs of the teeth. v = mesenchym of fin edge.
For an enlargement of (c), where the arrows are more clearly visible, click HERE (and close the window to get back).
(After HADORN & WEHNER)
About the cell migrations from the neural ledge, as just depicted in the Figure above, HADORN & WEHNER, p.162/3, say the following :
During neurulation the original blastema [of the neural ledge] lies on the inside of the neural fold, between the neuro- and skin-ectoderm (Figure above : (a)). When the neural plate has folded itself into a neural tube, the cells for (constituting) the neural ledge are situated in the dorsal connection zone (Figure above : (b)). From here they disperse, partly under the epidermis, partly between neural tube and myotomes, across different regions of the embryo. Thanks to special cell affinities they come to a halt at certain places. Here they often undergo very different differentiations. These derivates of the neural ledge are depicted in the Figure above (c). Especially conspicuous is their participation in the formation of the skeleton, which, after all, for its remaining parts has a mesodermal origin. The cells of the neural ledge produce cartilage and bone of the visceral arches [...], certain parts of the skull base and the dentine of the dental germs [...].It is probably here that the presence of an organic lattice is important with respect to the maintenance of the eudipleural ( = bilateral) symmetry : The migrations of the cells must take place symmetrically, and end up at several definite locations.
In some animal groups, for example in Ascidians (Tunicata) of the genus Cynthia, but also in Ctenophora and other animals, we see a mozaic development : Already in the egg cell there are a number of cytoplasmatic regions that are predisposed to develop specifically differentiated cells. They differentiate independently from neighboring cells : autodifferentiation. It is the cleavage of the egg cell that distributes these cytoplasmic types. The differentiation is independent from the positions of the cells and dependent only on the cytoplasm type that gave rise to them. So at least in these early stages no active (differentiation-determining) position information is present (HADORN & WEHNER, p.172).
But when it comes to organizing those differentiated cells and cell groups into organs that come to be located at definite places within the embryo, some sort of localization medium must be present.
The embryo of Cynthia develops according to predetermined cytoplasm regions in the egg. There are at least four different types of cytoplasm present in the egg. After fertilization these types arrange themselves in a polarly layered and eudipleural way as a result of flowing movement. And as a result of cleavings they end up in certain zones of the embryo, i.e. they are ordered. And only from this initial order they can be organized after having been differentiated. Left and right body halfs develop independently. If the cytoplasm types are initially mixed (by centrifugation techniques), the types still develop into their corresponding differentiated cells, but they do not organize themselves into a genuine embryo. It seems that after centrifugation there is no functional localization medium anymore to which the cells could respond in order to organize themselves into a genuine gastrula, neurula, etc. The next Figure illustrates the phenomenon of mozaic development.
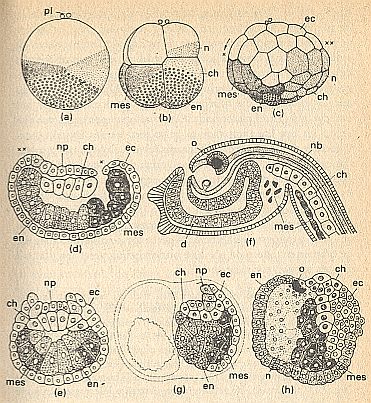
Figure above : Determination of development in Ascidians.
(a) Two-celled stadium, seen from left side (right-hand blastomer not visible). pl = polar bodies above animal pole.
White (signature) : transparent animal plasm for ectoderm (ec).
Small circles (signature) : vegetal plasm rich in yolk, for endoderm (en and d = gut).
Lightly speckled (signature) : grey crescent for neural tissues (n = presumptive neural tissue or material, np = neural plate, nb = neural tube) and chorda (ch, in later stadia signatured by white cells + large nuclei).
Darkly speckled (signature) : yellow crescent for mesoderm (mes, musculature).
(b) 8-celled stadium.
(c) Blastula.
(d) Median section of a gastrula (neurula). x, xx = corresponding places in (c) and (d) before and after gastrulation.
(e) Transverse section of the same stadium as in (d).
(f) Young larva with eyes (o).
(g) Half a neurula, developed after the killing of the left cleavage cell in the 2-celled stadium.
(h) Abnormal embryo, developed from an egg that was centrifuged during the first cleavage.
(After HADORN & WEHNER, from CONKLIN, altered.)
The cases of embryonic regulation -- more or less in contrast to mozaic development -- can be globally explained by the presence of an organic lattice (organic crystal), which expresses the totipotent nature of the germ. Partial cleavage in the median plane results in something like a siamese twin, like the case in crystals : Baumhauer twin. See next Figures.
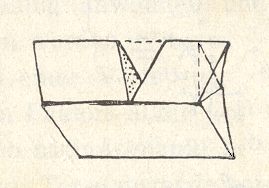
Figure above : The Baumhauer calcite twin, generated by locally applying pressure.
(After PRZIBRAM, H., Die anorganischen Grenzgebiete der Biologie, 1926.)
The next Figure illustrates some aspect of the internal structure of such a twin crystal.
Figure above : In the glided structure (B) of a crystal, successive planes of the atoms in the unglided structure (A) have moved a short distance, and the atoms have settled in new positions symmetrical to the old positions. We must see the twins as follows : up and below the right half of the array depicted in the right-hand image of the Figure. Twining is recognized by the presence of receding angles.
(After HOLDEN, A. and MORRISON, P., Crystals and Crystal Growing, 1960, 1982.)
There is no sharp difference between mozaic development and regulative development, only the fact (and its consequences) that in mozaic development the cellular system is determined to the several specific functions in an early stage of development, while in a regulating germ this is taking place at a later point in time, in virtue of cellular interactions.

Figure above : Diagram of a two-dimensional zoned crystal. It consists of (chemical) motifs that are arranged according to a rectangular point lattice. The upper half of the crystal consists of motifs that are slightly different from those of the lower half. So the crystal consists of two zones. As such it represents the two-dimensional analogue of a possible organic lattice containing two types of motifs that differ chemically and morphologically, but have the same symmetry, all arranged according to one and the same rectangular point lattice. In such a zoned organic lattice the direction of a partition (i.e. division into two halves) can make a difference, as the next Figures show.
Figure above : The zoned crystal of the above Figure yields two identical halves when partitioned vertically as indicated. These halves can be seen as 'monovular twins'. When the cut (and the subsequent shift of atomic layers) is only partial, we get 'siamese twins', a Baumhauer crystal twin (See Figure above ).
Figure above : If the above two-dimensional crystal is partitioned along the boundary between the two zones, then two different halves will be the result.
Instead of zoned crystals (organic or otherwise), we could also have to do with 'patched' crystals, and compare these perhaps with cases of mozaic development where it occurs in organisms.
In all cases where artificial separation and isolation of parts of a developing egg or embryo, or of an adult organism, nevertheless lead to the development or restoration of the Whole again (and also in all cases where we have the restriction that certain specific parts of the developing germ or adult organism must be present in minimal quantities), we must assume that any 'pre-design', supposed to be pre-existent in the form of a system of inhomogeneities, and as such constituting a position-information system, is necessarily periodic, or at least quasi-periodic, with respect to its internal structure. Only such a structure (of the pre-design) allows for the continuance of its (successful) development (even) after partitions have taken place. Or in other words, the internal structure of our supposed 'pre-design' must be periodic in order to survive partitions. This pre-design can regenerate and thus extend again its lattice, and, under the right parameter settings, restore its macroscopical pattern. Of course this can only happen when there is a sufficient supply of the right material. In this way it is supposed that the phenomena of regulation and regeneration as they occur in organisms, are partly dependent upon the restoration ability of the pre-design thanks to its periodic structure.
It is instructive to dwell a little longer at the phenomenon of regulation, which is, in any case, the most conspicuous ability of many developing germs, and, until now, defies detailed explanation. Let us quote HADORN & WEHNER, again (p.174/5).
In a large number of developing germs the components of eggs and embryos have prospective developing capabilities that go far beyond their actual prospective function. If, for example, one separates the two cells of the first cleavage of an amphibian embryo, that normally would have formed a right or left body half, then a complete larva develops from each of the two blastomers, and from this larva a completely normal frog or salamander. The egg material which was supplied to each half of the germ, is applied (used) totally anew. A new plane of symmetry is being formed and a new blueprint for the predisposition of organs emerges, of which the organization and developmental potency represent a reduced version of the developmental plan that originally was meant for the whole germ. In the case of such regulation processes factors are involved of a structured continuum which exerts influence on the cells that are sensitive to it [ HADORN & WEHNER then refer to a section where they discuss position-information, and which section we have reproduced in the previous document .] . Such a development, regulated by neighbor cells, is known as dependent differentiation.
Determination, Differentiation and Organization.
In the following we must clearly distinguish between determination, differentiation and organization, despite the fact that there are no sharp boundaries between them.
Determination of a cell means an acquired predisposition of it to differentiate at an appropriate point in the course of the development of the embryo. Differentiation means an actual change of the cell resulting in it to belong to a certain definite cell-type. And although changes of cell shape, as a result of differentiation, can play a role, and then more or less automatically entail supracellular structures to emerge, for example tubular structures, which is already a form of organization, differentiation generally is not the same as supracellular organization : While a given set of individual cells differentiates in this direction, and another in that direction, etc. (and as such having to do with some reorganization within these individual cells), the differentiated cells must now be spatially distributed such that a definite pattern of these cells is the result. Only then organization is achieved.
So determination takes place at a certain point in the development of the germ, later followed by differentiation and organization. Differentiation is in most cases irreversible, and also determination is often irreversible. Let's quote again HADORN & WEHNER, p.177--179, with respect to experiments with amphibian embryos.
At the beginning of gastrulation a part of the presumptive epidermis [ presumptive epidermis, i.e. a cell group that will, in normal development, later become epidermis (which, roughly, is skin tissue) ] , for example the future abdominal skin, is exchanged for a piece of presumptive neural-plate tissue of another individual (See next Figure, (a), (b)). The implantates grow together (fuse) with the surrounding tissue and locally develop, in correspondence to their new environment, into brain or skin (next Figure, (e), (f)). Evidently, the pieces of ectoderm were, at the time of transplantation, not yet determined. If, on the other hand, one transplants blastemas from the same area, but not until after gastrulation has taken place, when the roof of the primitive gut already lies below the ectoderm (next Figure, (c), (d)), then, it's true, the transplanted pieces of tissue are still accepted, but now they develop according to their o r i g i n . The, according to the developmental plan (See Figure in the previous document ), presumptive epidermis also in the new situation becomes epidermis and as such later rejected from the brain. The presumptive neural material develops within the abdominal skin further into neural-plate tissue, invaginates and subcutaneously forms some part of brain, as it would also have done at its original position. Clearly, during gastrulation, both ectoderm types are irreversibly determined.
As a result of the determination, the ability to autodifferentiate is concluded. Therefore, in the determined blastemas, also when one breeds them isolated as explantate in a tissue culture further, histogenesis starts [And in the latter case this means : outside the embryo.] and does not end until the full differentiation, resulting in the specific cell-type, is completed.
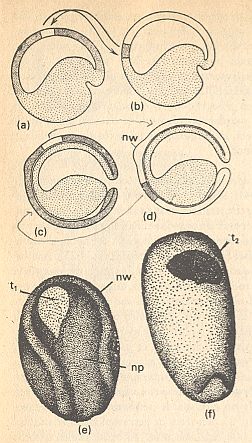
Figure above : Transplantation experiments on embryos of salamanders (Triturus). Blastemas differently marked by natural egg pigments.
(a) Lightly colored transplantate from the presumptive abdominal epidermis of early gastrula, develops within the (presumptive) brain area in accordance to its location (i.e. in accordance to its newly acquired place, the presumptive brain area). This location is t1 in (e) .
(b) The dark transplantate fom the presumptive neuro-ectoderm develops, according to its location, into abdominal skin : t2 in (f) .
(c),(d) Corresponding exchange experiments with transplantates from a late gastrula (nw = anterior edge of the neural embankments).
(e) Receptor of implantate (a) in neurula stadium (np = neural plate. nw = neural embankment).
(f) Receptor of implantate (b) as neurula, ventral view.
(After HADORN & WEHNER, from SPEMANN.)
Induction
It is known that in the course of the embryological development of many animals a certain cell group induces changes in an adjacent cell group. This is called differentiation by induction.
This induction (i.e. the fact that it indeed is a form of induction), can be demonstrated by transplantation experiments on amphibian embryos. HADORN & WEHNER, pp.181, describe them as follows (See also Figure below) :
A part of an early gastrula from the presumptive mesoderm, which, before gastrulation, lies, as edge zone [...], above the upper lip of the primitive mouth, and later forms the roof of the primitive gut (See,in the previous document the Figure of fatemap and the Figure of gastrulation and neurulation ), is being transplanted into the future abdominal epidermis (next Figure, (a),(b)), or carried into the blastocoel (next Figure, (a),(c)). In the first experiment the tissue invaginates [...], in the second experiment it is, during gastrulation, pushed against the ventral ectoderm (next Figure, (d)). In virtue of autonomous capabilities the implanted piece of tissue differentiates into a chorda and remaining mesodermal tissue. [ But, and this is what induction is all about, ] Moreover, it exerts an influence onto the adjacent parts of the receptor-embryo, which one calls induction : [ To be noted first ] The implanted tissue completes itself with the help of the surrounding material of the receptor, insofar as it needs it for primitive segments and lateral-plate mesoderm. [ And, to be noted secondly ] In the ectoderm, which lies on top of the implanted tissue, moreover, the formation of a neural plate is induced in the receptor (next Figure, (e)). In the endoderm [ second experiment ] a secondary primitive mouth can emerge, and within the mesoderm, mobilized for its realization, pronephros channels can develop (next Figure, (f), ev) [ by induction ].
The fragment, originated from the edge zone, of presumptive mesoderm organizes a secondary embryonic primordium by regulative formation of somites and through induction, an embryonic primordium that lies symmetrically around a new body axis. In the formation of this secondary embryo, parts of the receptor-embryo can be involved that are so extensive, and the realization of the secondary body parts (head, trunk, limbs, tail) can reach such a high degree of internal and external organization, that the receptor grows into a double monstrum (next Figure, (g)). The area in the blastula and the early gastrula, from which this inductive and organizing influence emanates, was called 'o r g a n i s a t o r ' by Spemann [ It is perhaps better to call it a 'differentiator'. Only -- as I assume -- when induction takes place within an embryo, organization (of the differentiated cells) will follow, in virtue of just this embryo.] . The cell material of this primary induction-system lies concentrated in an organization center. [ And ] This is identical to the edge-mesoderm that, as roof of the primitive gut, invaginates [...]. This edge-mesoderm autonomously differentiates mainly into chorda and muscle tissue [ while it induces the ectoderm which lies on top of it ].
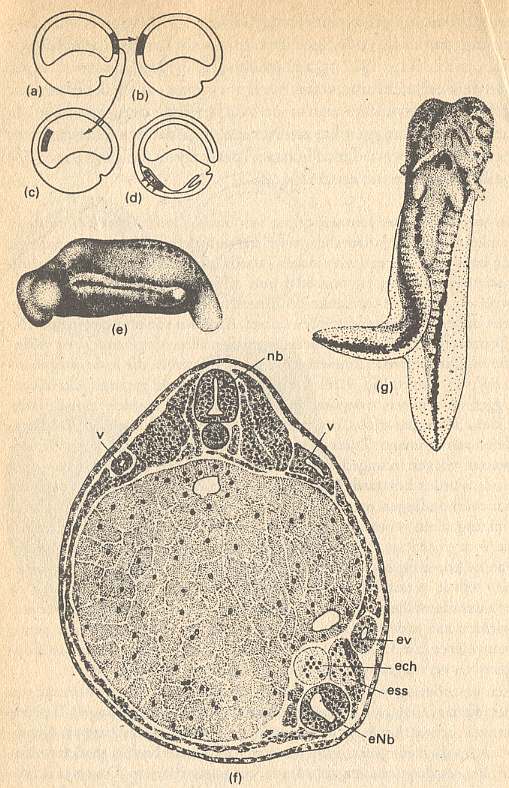
Figure above : Transplantation of organizer (organisator).
(a) Extracted area from the dorsal lip of primitive mouth.
(b),(c) Different kinds of implantation.
(d) Shift of implantate by gastrulation and effect on ectoderm.
(e),(f) Germ of Triturus taeniatus, in which, while still in the state of a young gastrula, was implanted left ventrally a piece of the dorsal lip of primitive mouth from a gastrula of Triturus cristatus (From Spemann).
(e) Germ seen from left side, with extra-embryonal primordium containing neural tube, primitive segments, auditory vesicle and tail, all induced by the organizer.
(f) Transverse section through the center of the germ. Implantation site : bottom-right. The implanted tissue develops a(n) (extra) chorda and a part of the right-hand primitive segment.
(g) Double development. Larva of salamander with a secondary embryo primordium, induced at the abdominal side by an implanted piece of the dorsal lip of primitive mouth (From Holtfreter). ech = extra chorda. enb = extra neural tube. ess = extra muscle segment. ev = extra pronephros channel. nb = normal neural tube of receptor. v = normal pronephros of receptor.
(After HADORN & WEHNER)
(Continuing quote)
Based on the results of transplantation experiments one can ask oneself what the significance and the necessity of the organization center are in normal development. Immediately after fertilization the cytoplasm of the future edge-mesoderm lies concentrated in the grey crescent of the frog egg (See next Figure).
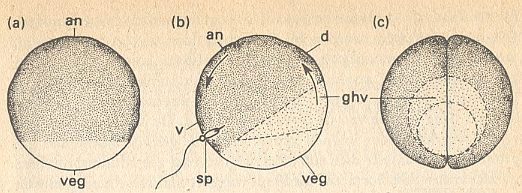
Figure above : Symmetries in the amphibian egg (frog).
(a) Egg cell before fertilization, with an animal and vegetal pole (an and veg).
(b) The appearance of the grey crescent (ghv) after fertilization (sp = location of sperm cell penetration) and orientational rotation (arrows). d = dorsal. v = ventral.
(c) View at vegetal pole (posterior) after the first cleavage in the plane of symmetry.
(After HADORN & WEHNER)
(Continuing quote)
Its inductive influence [ i.e. the inductive influence of the future edge-mesoderm ] evidently goes back to the functional structure of the egg [...] which [ functional structure ] is predetermined by maternal genes. Irradiation with ultraviolet rays of the grey crescent in an egg which has not yet undegone cleaving, leads to a failure of the induction in a later stadium. Embryos appear without nervous system. This damage effected by ultraviolet rays does not strike the nucleus -- which can be proved by nuclear transplantation [...] -- but the cytoplasm. The effect of irradiation can be neutralized by injection of cytoplasm from the grey crescent of a non-irradiated donor. If eggs of salamanders are being divided into two pieces, while these eggs are still in an arbitrary early stage before the beginning of gastrulation, then only median halves regulate which have received sufficient cell material from the edge zone [...] into a complete embryo. If the germ is divided frontally into two pieces, then only the dorsal half receives the inducing blastema. Only this half gastrulates and regulates into a normal larva [...]. The ventral 'partner' stays, it is true, alive, but misses a neural tube, a chorda and somites. Only an amorphous abdominal part develops, which consists of endoderm cells rich in yolk, and a skin layer [...]. Also in humans sometimes such an amorph is born, in addition to a normal twin.
That the roof of the primitive gut is, as primary induction system, indispensable for normal development, is made evident by the so-called sandwich combinations. One isolates ectoderm, which is still not yet determined, of an early gastrula (See next Figure, (a), (d)). In the nutritient solution one places a part from the organizer-area onto one of the explantates. The ectoderm of the latter quickly grows around and over it (next Figure, (b)). In the presumptive ectoderm subsequently the most divergent organs and tissues, characteristic of a normal development, and especially parts of the nervous system, are induced (next Figure, (c)). The control explantate (next Figure, (e)) develops into a cell heap, consisting only of non-specific epidermis cells, arranged in rows.
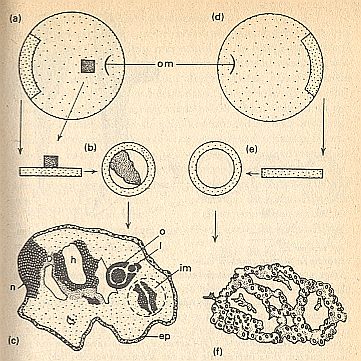
Figure above : 'Sandwich' experiment.
(a) and (d) Donor (early gastrula with primitive mouth (om)), from which a piece of ectoderm (signature : intermediate density of speckling) and a piece of mesoderm (densely speckled) is taken.
(b) and (e) Explantate with and without added mesoderm.
(c) Inductive effect of the added mesoderm (im = encircled implantate [ I assume that the authors mean : encircled area = original explantate, which because of growth is now embedded within a larger whole.] ). Differentiations in the target tissue : neural tissue (n), brain (h), eyes (o) with lens (l) and normal epidermis (ep).
(f) Atypical epidermis.
(After HADORN & WEHNER, from HOLTFRETER)
(Continuing quote)
From this it is evident that without the influence of the roof of the primitive gut, there can be no determination into nervous tissue. This is also demonstrated by the abdominal parts [ referred to earlier ] [...]. The inductive effect appears during the course of gastrulation, when the ectoderm is internally lined with endoderm (See Figure on gastrulation and neurulation in the previous document). It continues until after neurulation (See next Figure).(End of quote)
In (c) of the above Figure one can see that indeed differentiation has taken place (by induction), but only a little degree of (higher-level) organization. It seems therefore that the initial isolation of presumptive tissue from the embryo, resulting in an explantate and allowed to grow, has destroyed (most of the) the pre-existing global predesign (organic lattice grown by non-equilibrium crystallization).
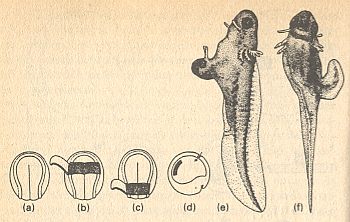
Figure above : Transplantation of different regions of the organizer of the neurula of a salamander, to show position-specifity of inductors. Different pieces (fragments) of the roof of the primitive gut, lying far (away) from its anterior end, of the neurula, are transplanted to embryos that are in a state of beginning gastrulation.
(a) The embryo that supplies the pieces, in dorsal view.
(b)--(d) A slice of the neural plate is cut away, and an anterior (b) or posterior (c) part of the roof of the primitive gut is exposed, and (d) implanted into an embryo that is beginning gastrulation.
(e),(f) Larvae.
(e) After implantation of an anterior part of the roof of the primitive gut.
(f) After implantation of an posterior part of the roof of the primitive gut.
(After HADORN & WEHNER, from MANGOLD)
From the experiment depicted in the above Figure it is evident that position-specific properties are present in the roof of the primitive gut. An implantate, consisting of mesodermal tissue which develops into primitive-gut roof of head (i.e. into that part of the primitive gut that lies in the region of the future head), induces parts of the brain, eyes and attaching threads. On the other hand, material from the presumptive primitive-gut roof of trunk induces in the ectoderm a spinal cord, trunk muscles, fin edge and parts of the tail. The same position-specific properties are demonstrated in sandwich experiments (HADORN & WEHNER, p.185).
The induction itself turns out to be non-species-specific. Apparently in all vertebrates the determination of some tissue, or group of cells, to develop into a nervous system takes place according to corresponding mechanisms and under the influence of the same or replaceable substances. So inductors act, like hormones, within broad limits non-species-specific (HADORN & WEHNER, p.184).
According to WADDINGTON, 1961, p.70 (of work cited below), evocation (which is induction s.str.) can be obtained by chemicals, like methylene-blue, which surely does not occur in a normal embryo. WADDINGTON continues that this means that the cells that react to the organizer, are, in one way or another, already prepared to develop into the various tissues of the embryo, and that they can be stimulated to do this, not only by the organizer itself, but also by several other stimuli, a number of which could work in a way very different from that of the natural stimulus.
So according to these considerations of WADDINGTON, and also to those given earlier, we see that induction -- at least pure induction, or 'evocation' -- is a very local process, presupposing some (pre-existing) disposition of the target cells, which are only to be triggered to commence their differentiation. And together with the above mentioned position-specifity of the inductors, things point to a pre-existing design or scaffold, which we call the organic lattice, macroscopically shaped by non-equilibrium crystallization. As such it forms the lay-out of the spatial distribution of organ primordia.
HADORN & WEHNER, p.188, report (1977) that despite years of intensive research the many problems with respect to the mechanism of induction are still unsolved. About the supposed mechanism WADDINGTON, C., The Nature of Life, 1961, says the following (taken and translated from the Dutch edition 1966, Het Leven, een biologisch-filosofische benadering, p.73/4) :
It is already known for a long time that, generally speaking, the explanation should be looked for in the fact that the fertilized egg is not an amorphous mass of cell plasm, in which a nucleus is present, but structured in a certain degree. This implies that the egg consists of a number of different parts, mutually arranged into a fixed proportion [ See Figure above ]. If the egg is partitioned into smaller cells, then some nuclei end up in cells of a certain part and other nuclei in cells that come from another part. We must assume that these initial differences between the parts of the egg activate different groups of genes (or perhaps we in fact should speak of different groups of cistrones [ sub-genetic units ] , but this modern refinement of terminology is in the present context barely necessary). In fertilized salamander eggs, for example, one can distinguish a part having a grey color and which is called the grey crescent [ See again Figure above ]. Precisely the cells, which are formed from this part of the egg, later form the third quadrant that was discussed earlier [ In an ealier drawing, WADDINGTON, p.65, depicts the salamander egg laterally in early gastrulation, and roughly distinguishes four quadrants : top : 2 and 1, bottom : 4 and 3, where quadrant 3 corresponds to the area of the blastula or early gastrula that will invaginate in the course of gastrulation.], and function as organizing center. The material in the grey crescent must be such that in these cells a begin becomes to be established of the realization of certain hereditary capabilities. At the top of the egg [ i.e. its upper part ], [ namely ] the material from which the quadrants 1 and 2 are being formed, a different series of genes will become active. When quadrant 3 invaginates into the interior of the egg and establishes contact with quadrant 1, and induces there the formation of the nervous system, it means that this induction entails an alteration of the cell-plasm properties of the cells from quadrant 1, namely in this sense that the genes, involved in the protein synthesis of nerve cells, are being activated.According to HADORN & WEHNER, p.189, this acivity of certain groups of genes could be triggered by activation of a type of messenger-RNA already present fully-fledged, but (still) masked, as well as by the appearance of new transcriptions.
From all this it is understandable that the r e a c t i o n-part of the induction process is species-specific. As HADORN & WEHNER, p.185, say,
The head region of an embryo of the Urodela (salamanders) exerts a non-specific inductive influence, which effects, in an implanted piece of epidermis originating from Anura (frogs [...]), the formation of the structures of mouth and fixing organs that are normal for the relevant location [ within the animal ] . However, the 'execution' of this assignment takes place according to the genetic constitution of the induced cells. Therefore the horn jaws and fixation discs, characteristic of Anura, emerge in the head of the salamander. In the reciprocal experiment the head of the tadpole becomes equipped with epidermal organs of Urodela, i.e. with genuine jaw teeth and fixation threads. This genetically determined reaction is brought about autonomously. The same holds for many other transplantation experiments in which action- and reaction-system possess a different genotype (mutants, species).As has been said earlier, pure induction, or, as it is called, evocation, is a local process, and as such not involving (large-scale) organization. Therefore it can be interpreted 'atomistically' which here means the action of some local element on some other local element. It is, to be sure, a part of a broader system, but it is not steered by that system-as-a-whole. The latter case, wherever it is seen to take place, is sometimes called 'determination by a continuum' and contrasted with 'atomistic or local determination'.
There was a moment that the induction process, at least certain aspects thereof, could be explained by an atomistic theory. This would have brought the most fundamental events in embryonic development within the reach of such a theory. [...]
I had demonstrated that induction not only occurs in amphibian embryos, [...], but also in embryos of birds. For example in the embryo of the chicken there is also an organization center, and when small parts of it are cut out and implanted at another location of the embryo, they will induce the formation of a new nervous system and other embryonic organs in the surrounding material.
Studying embryos that had been abnormally influenced this way, I came to the conclusion that their construction [ structure ] sometimes demanded an explanation by a continuum theory [...]. An implanted piece of organizer can, for example, cooperate in such a way with tissues emerged under its influence, that they together form a more or less complete embryo. It here seems evident that the inducing organizer and the reacting material are both determined by the mutual relation of the parts. But there were also many [ cases of ] induced embryos in which this was not the case. If, for example, one brings an organization center in contact with material that normally would form a nervous system, and adjacent to it [ i.e. to that material ], supplies a second chunk of the same kind of material, it is possible that induction not only takes place in the normal, but also in the extra material, eventually resulting in the formation of two complete nervous systems, that is to say : much too many for one single complete and normal embryo. With respect to such a case one cannot say that induction is the visible result of a necessary relation between the organization center and the remainder of the embryo, and nothing more. The organizer seems to 'induce' totally independently of the fact whether there is, or is not, a need for more nerve tissue for the completion of the system. 'Induction' in such cases is in fact something totally different from 'organization' of the tissues, that originated under its influence, into one single embryo. Discussion about this topic in Spemann's terminology has the drawback that in it induction and organization are more or less indissolubly connected, because, according to him, the most important property of an organization center is the fact that it induces. Therefore I use the word 'individualization' to signify the construction of tissues into one system, and later my friend Needham suggested the word 'evocation' signifying something that otherwise would not have been developed, a process that I earlier had called 'pure induction'.
If 'pure induction' or 'evocation' can occur independently of organization, then an atomistic approach of this aspect of development is not expected to meet objection. One could, for example have the opinion that simply by the activity of a certain chemical substance, produced by the organizer, the surrounding cells are going to develop into something that they otherwise would not have become. But this supposition was opposed to Spemann's aversion to atomistic theories. I spent the summer of 1931 in his laboratory and in that of his pupil Mangold in Berlin. They had little confidence in an attempt to search for an active substance that would be released by the organizer. But as soon as I was back in my own laboratory in Cambridge, I began to do such experiments with my own material, the chicken embryo. And when, in the next spring, the salamanders were laying eggs, Spemann himself and several of his collaborators in Germany also performed such experiments. As a result it was immediately clear that in salamander as well as in chicken embryos organizers were capable of induction, or, as I should have to say, evocation, even if they were killed. So there was most certainly nothing vitalistic in the process and it seemed indeed all-out evident that here an atomistic theory was called for.
One should thoroughly be aware - which is not the case in all authors -- that the sole production of something new from the surrounding world does not represent all there is in the process of embryonic induction. This is in fact only that aspect of it, that I call evocation. It is very probable that this will be explained by the influence of a certain chemically active substance, released by the organizer, but even if we consider evocation atomistically, then this does not necessarily mean that such hypotheses are suitable to explain the whole developmental process. For example, a dead piece of tissue can influence its surroundings such that the latter develops into a complete and normal embryo. Because the dead tissue evidently cannot be considered a part of the embryo, nothing can stop us to explain the evocation of such an embryo atomistically. However, the embryo itself displays a strict organization in all its different components, as brain, spine, kidneys, muscles, etc., all having their normal form, and being mutually arranged in the normal way. The mutual relations between the parts of the embryonal organization evidently are of a totally different nature than the relations between the dead tissue and the embryo. In fact it seems rather clear that this individualization of the developing tissues resulting in a normal well-organized whole, asks for a kind of continuum explanation [ Our theory of organic lattices, as given on this website, represents a proposal along such lines. ]. Some people did not see this, and it is probably because of this that they thought that an atomistic approach of one single aspect of the induction process meant that the total process of embryonic development could be explained in the same way.
In addition to the above discussed primary inductor, there are many other inductors. There appears to be a hierarchy of inductors in a single developing embryo.
Particularly in embryos of vertebrates the spatially as well as the temporally programmed formation of organ primordia realized by a series of induction processes, in which blastemas, owing their specific properties to higher-order inductors, in turn are going to act as lower-order inductor. Such a hierarchy, forming the basis for a series of dependent differentiation processes, is depicted in the next table.
| Active Blastema | Rank of Inductor | Intrinsic Differentiation | Reacting System | Effect of Induction |
| Mesoderm of Roof of Primitive Gut | First Order |
Chorda, Musculature, etc. | Presumptive Neuro-ectoderm | Neural Plate, Neural Tube |
| Neural Tube | Second Order | Brain, Eye Cup, Spinal Cord | Presumptive Skin-ectoderm | Nasal Groove, Lens, Ear Vesicle |
| Ear Vesicle | Third Order | Labyrinth (dermal) | Mesenchym | Cartilagous Labyrinth |
Table above : Some examples of the hierarchy of inductors in the embryo of vertebrates.
This hierarchy [pre]supposes mechanisms of regulation which guarantee that the inducing influences take place at the moment at which the reacting systems are sensitive to them. For this it is necessary that in both systems the phase-specific gene assortments are switched on and off at the right moment [...].The local nature of induction is also preserved when considering a whole hierarchy of inductions, because each individual induction is based on direct contact or very narrow spaces between inducing and reacting systems.
Many differentiation processes, as a result of determination by induction, involve many reacting systems, i.e. a certain final induction is often prepared for by preceding inductive influences (HADORN & WEHNER, p.187).
All the above demonstrates that induction, as the most fundamental process in embryological development, is in fact only : determination of cells for a particular direction of their differentiation. And because differentiation of cells often entails that they acquire particular shapes (and surface properties), they can spontaneously arrange into certain definite patterns, like tubes, spines, and the like. But apart from such more or less simple structures, no global patterns result from induction, or a hierarchy of inductions, alone. So the global spatial structure of a multi-cellular organism is not explained by the phenomenon of induction. Other phenomena, like concentration gradients and/or organic lattices, must be involved in the generation of global spatial structure of such an organism. They must organize (i.e. spatially arrange or spatially distribute) the differentiated cells, first of all into organs (which always have a definite structure and shape), which WADDINGTON calls 'individuation', and then (not temporally meant) into the pattern of the whole organism. This 'first into organs then into the whole organism' is an integrated state of affaires and can collectively be called the patterned spatial distribution and arrangement of organ primordia.
Let us quote WADDINGTON, The Nature of Life, 1961, Dutch edition 1966, pp.74, where he writes about these three levels, viz., cell differentiation (autodifferentiation and induction, resulting in definite cell types), formation of organs (individuation), and arrangement of organs into a definite pattern, resulting in the internal and external morphology of the individual organism.
One of the most important properties of animals -- so evident that its importance is often overlooked -- is the fact that they are built up by a restricted number rather sharply separated cell types. There are nerve cells, muscle cells, kidney cells, liver cells, etc., and each of these main types can have a certain number of subordinated variants, but we find very few cells that can truly be considered intermediary between the main types. In animals one structural type does not grade into the other, but [ animals ] are built up by bordered, clearly mutually separated organs. The development [ of an animal ] does not lead to a continuous series of possibilities, but results in a discontinuous series of different cell types. Every cell type has a complicated houshold requiring the activity of many genes. Therefore the gene activities must be organized [ in the sense of orchestrated ] into mutually exclusive systems.So now we have seen something about differentiation of cells during embryological development. It is, as one could say, representing the micro-organizational level.
Evidently these are dynamical systems [...]. Embryonic cells, as for instance those of the top of a salamander egg are not quietly waiting [ while ] in constant possession of the property [ ability ] to develop into either nerve tissue, or skin, etc. From the moment of fertilization on, their nature changes continuously. Only some considerable time after fertilization, when these proceeding changes have moved forward quite a long way down the line, the cells are able to respond to an organizer which can trigger them to go in, or not in, the direction of the development into nerve tissue. If influencing by an organizer is delayed too long, their readiness to react disappears again. During their continuous change there is only a particular state during which they are, as we say, 'competent' to respond [...]. And if then such a system of gene activities is stimulated, either by the original nature of a particular region of the egg, or by the cell plasm, that is altered by an organizer, the cell does not transform instantaneously into a completely developed nerve cell, muscle cell, or whatever. It is just the beginning of a long, long way of development, in which one stadium will grade into the other, until the final stadium is reached. We can visualize this by supposing that in each individual stadium a certain group of genes is active, synthesizing certain proteins. The presence of these proteins, however, alters the cell plasm, resulting in the activation of another group of genes which form new proteins and thus entailing again a change of cytoplasm, resulting once again in the activation of other genes, etc.
The fact that the development ends with a series of sharply separated end results, intercalated by only a few intermediate cases, means that these continously changing gene activities interfere with each other in such a way, that development is possible only in certain particular directions. It seems to be the case that in a cell, in which genes, involved in the production of muscle proteins, work hard, the genes, [ potentially ] involved in the production of nerve-cell proteins, cannot be strongly active at the same time. When the cell starts with the production of a particular protein type, then opposite activities are rendered impossible.
Until now we have discussed only the development of parts of the egg into cells of a determined type, while having passed over the phenomena of the individualization, by which these cells are being arranged [ i.e. spatially distributed ] resulting in organs with a determined shape and pattern. I am sorry that, as a biologist, I must admit that we barely have any idea in what way this takes place. Here, undoubtedly not only chemical processes will play a role. Development starts off from a more or less spherical egg, and from it an animal develops that is anything but spherical, it has legs, a head, a tail, etc. and internal organs that also have determined constant forms. This cannot be explained with any theory whatsoever, when it limits itself to chemical concepts, like the theory that genes control the synthesis of certain proteins. We must, in one way or another, find a way to be able to point to the physical forces needed to bring the material to the right place and mould it in the right form.WADDINGTON then gives an example of a simple case, where changes of cell shape can entail a specific spatial ordering of those cells, resulting in a chorda. Certainly, there will be many cases where organs, or parts of them, are formed in a comparable manner.
Whatever precisely is the nature of the physical forces that form the developing tissues into organs, they decidedly must be organized in self-regulating systems. The 'regulation' is one of the most conspicuous properties of embryos, which means that if parts are cut away or if they are damaged in different ways, they show a stong tendency to nevertheless reach a normal end result. The forces, by which the different forms are produced, cannot go their business totally independently and without a proper response to what happens in the environment. They must have the same capabilities to compensate for abnormalities and to adapt to their environment, as we saw in the case of the chemical processes that play a role in tissue differentiation.So we have seen that, although induction (i.e. pure induction, evocation), resulting in cell differentiation, seems to allow for an atomistic approach, the organization (except for simple cases) of them into organs (individuation), and the patterned distribution of these organs, require a different approach, involving global, in contrast to local, factors.
In our consideration of development we thus arrive at the conclusion that our theories must include, in addition to the simple atomistic approach of DNA sequences and protein sequences, different levels of organization.
First of all we have to do with cybernetically organized systems [ i.e. systems that self-regulate by feedback processes ] of gene activities, resulting in a series of mutually excluding chreodes [ A chreode -- a concept introduced by WADDINGTON -- is a more or less stabilized route -- visualized by an inclined valley along which a ball, representing the state of the system, rolls down to a particular end point, and so a route which a developing cell takes, in order to become a definite cell type. ] , in accordance with which [ i.e. in accordance to these chreodes ] the different parts of the egg alter, until they finally have developed into precisely determined cell types.
Next we have to assume [ the presence of ] organized systems of physical processes, that accomplish the individualization of cell groups to organs having a fixed form and size.
[ In addition to the above described levels ] there can be still other, higher levels of organization. For example there are remarkable 'regionality ' phenomena. In a fairly early stadium of the chicken embryo one can demonstrate that the posterior limbs differ from the anterior ones [ which are the wings ] in a very special way, although at that moment both are still small stumps, consisting of seemingly identical cells. If, however, a small piece is taken from the upper [ i.e. a proximal ] part of the leg and placed close to the wing tip, one could, in some cases, observe that it grew into a claw under these new conditions. This means that at the time of transplantation it was not yet determined into what part of the leg the cells would develop. Taken from the upper part (i.e. the thigh), they became the distal end (i.e. a toe), but did preserve the property 'posterior limbs' in contrast to 'anterior limbs', resulting in the fact that, although they now develop within a wing [ i.e. within the area of a developing wing ] , they do not grow into a wing tip, but into a leg tip [...]. We do not have any idea how to explain these regional properties. Is there a chemical difference between all leg parts, whether they be either thigh or claw, on the one hand, and all wing parts on the other [ because of the early determination of the cells as leg cells or wing cells ] ? We simply do not know, and it is possible that we are confronted here with a still more complex level of organization than [ connected with ] the levels that regulate tissue differentiation and individualization.
Evidently, an incipient understanding can be case only if we establish that the processes we investigate force us to apply a theoretical way of approach in which the concept [ of ] organization plays a role [ WADDINGTON points to a version of organicism where systems and levels of organization are paramount, in contrast to an atomistic or reductionistic approach. ] . Where does this organization come from? How is it possible that gene activities are regulated in such a way, that they interfere in a manner by which sharply distinguished, mutually exclusive chreodes appear? It is beyond rational sense to assume [ i.e. it would be wrong to press the atomistic approach too far into the mentioned problems of the global organization of a developing animal ] that if we take a series of random[ly selected ] genes, each of them capable of regulating the synthesis of a certain protein, and put them together in a single cell, they then inevitably get into interaction with each other in this way. But of course in biology we do not have to do with random[ly selected ] genes, that got together by accident. The organisms we see in the world around us, are the result of evolutionary processes, their organization -- and that means the organization of their gene activities, by which chreodes are determined, and their morphogenetic forces by which organs are formed in a fixed pattern -- must be accomplished by evolution.
Gradient systems in developing embryos.
Some theories speculate about systems of chemical concentration-gradients, that in one way or another appear within the developing embryo, and that represent a kind of map, a map, where the different locations in that embryo are marked by a concentration profile of one, but in most cases several different, chemical substances. So a given location within the embryo has a particular chemical nature, and a cell, finding itself at such a location, chemically reacts (by one or more of its molecular components) in a specific way with that particular chemical environment, specific for the given location. This "chemically reacts in a specific way" is, in fact, the act of recognition : The cell 'knows' where in the embryo it is. And we might say : "knowing its location", it now knows what to do". But, according to me, this is not entirely correct. The recognition by the cell of its location is a chemical event, and this will change the cell's cytoplasm in a definite direction. And this particular change of cytoplasm will cause the switching on (or off) of a particular gene or series of genes of the genome of that cell, causing the cell to behave in a particular way. So the location of the cell entails the cell's differentiation, and this location is represented by a concentration profile brought about by concentration gradients. But of course within the developing organism this connection between location and differentiation can and will often be overruled by other factors.
The importance of gradient systems lie in the fact that they, for the first time, represent something that accounts for the spatial organization of the developing embryo. However, when the (still hypothetical) gradient systems turn out to be very complicated, we are thrown back to the question as to where such a gradient system -- such a spatial system -- comes from. A second difficulty is whether such systems are robust enough. The theory of organic lattices provides for both of these difficulties.
In the following we present a long quote, concerning the organization by gradient systems in embryos, from HADORN & WEHNER's textbook, pp.189, including one of their Figures.
Cytoplasmatic gradients in echinoderms.
Fragments and blastomers [ the latter are individual cells of the blastula (stadium) ] of sea-urchin eggs only then regulate into a complete embryo [...] when they contain animal as well as vegetal regions in a certain proportion, as is also the case in uncleaved eggs. If, however, a sea-urchin egg is divided into two parts perpendicular to the animal-vegetal axis, then only incomplete larvae develop. Stainings [...] and natural pigment areas offer the possibility to find out the prospective destination [...] of the blastomers that lie in stacked rings [ and as such constituting the blastula ] (See next Figure, (a)).
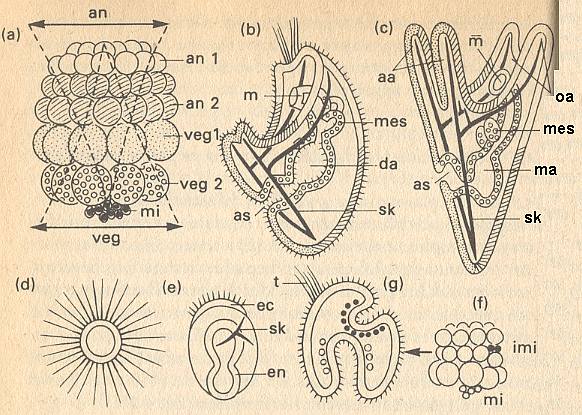
Figure above : Prospective meaning of the blastomers and gradient system in a sea-urchin embryo.
(a) 64-celled stadium. animal cell rings an 1 (white) and an 2 (hatched). vegetal cell rings veg 1 (speckled) and veg 2 (open circles). mi = micromers (dark). an = animal gradient decreasing toward vegetal end. veg = vegetal gradient decreasing toward animal end.
(b) Young larval stadium. as = anus. da = gut, in three parts. m = mouth. sk = skeleton. Signatures (hatching, etc.) according to (a).
(c) Pluteus larva. aa = anal arms. ma = stomach. mes = mesodermal sacs. oa = oral arms.
(d) Isolate from an 1.
(e) Isolate from veg 2. ec = ectoderm. en = endoderm. sk = skeleton.
(f) Implanting of micromers (imi, black). own micromers : white.
(g) Result of experiment (f). t = tuft of vibratory hairs.
(After HADORN & WEHNER, partially from HÖRSTADIUS)
(Continuing quote)
The cells of the first animal ring (an 1) yield the mouth ectoderm, that, in addition to short cilia [ hairs ] covering most of the body, also forms the tuft consisting of long vibratory hairs and being restricted to the animal pole ( [...] See Figure above (b)). This tuft consists of long, motionless stereocilia. When one cultivates this first ring of animal cells in isolation, an ectodermal vesicle appears of which the entire surface is covered by these stereocilia (above Figure, (d)). This autodifferentiation proceeds "more animally than animally" and therefore not really mozaic-like [ because it does not result in a normal tuft ]. Such extraordinary animal developing tendencies are, in normal development, kept in check by influences of the vegetal pole of the egg. This is, among other things, demonstrated by what happens in [ experimental ] blastomer combinations.More recently, there has been found at least one substance that is present in an embryo as a gradient, determining spatially distributed differentiation, in the present case differentiation along the main body axis of the embryo of a fruit fly (developing into a larva) (next Figure). With respect to this, let us cite BALL, P., Designing the Molecular World, 1994, p.311/2 :
If one adds cell ring an 1 together with the micromers (mi), then, remarkable enough, a normal pluteus larva develops. So, for the development of a complete larva surely not all cytoplasm types -- as in a true mozaic germ [...] -- are needed. Thus, in the an 1 / mi combination all prospective endoderm is absent, and nevertheless a pluteus gut of normal size develops. Only an equilibrium between animal and vegetal developing tendencies is required. Many experiments with isolates and combinations prove this. The above Figure (e) shows the effect of cell ring veg 2 (vegetal cell ring 2). Here, it is true, much gut endoderm develops, corresponding to the prospective meaning [ prospective function ] (above Figure, (a)--(c)), but in addition to it also an ectoderm strip provided with short vibratory hairs, and some skeletal material, that in normal development is only delivered by the micromers [...]. If to veg 2, mi is added, then the ectoderm with its vibratory hairs totally disappears [ because then we have an extreme vegetal whole ].
On the basis of all these experimental data one can draw the conclusion that in the sea-urchin embryo two gradient systems at the same time, form each other's antagonists and each other's synergists (above Figure, (a)).
From cell ring an 1 to the mi-zone, a decreasing animal gradient proceeds, while in the same direction a vegetal gradient gradually increases. In every part of the embryo differentiation takes place as a reaction to the resultant that applies on the spot, of the activities determined by both these gradients [ i.e. gradients, present in an embryo, together determine a matrix or pattern of triggers. What is triggered are certain definite cell reactions consisting of cytoplasmatic changes that in turn activate or de-activate certain gene segments. ]. These determining properties are, however, everything but stable. For example, if one inserts micromers between an 1 and an 2 (above Figure, (f)), then there they form a second vegetal activity maximum, that is going to compete with the gradient systems of the receptor-embryo. There where in the ectoderm the transplantate is, endoderm is being formed, that, following the mi, invaginates as secondary gut primordium (above Figure, (g)). The tuft of vibratory hairs does not appear at the original animal pole anymore, but at the newly determined most expressed animal position.
In gradient systems every cell region influences the direction of differentiation of all other zones of the embryo [ as transplantates have shown ] , as it is itself controlled by the whole. It is in this respect, that such a [ gradient ] system differs essentially from the induction system of vertebrates [...]. As activating systems, inductors determine the blastemas being dependent upon them, and this induction generally requires a direct cellular contact, while in a gradient system the determining equilibrium proportions are determined in a supracellular way [ i.e. at a level above that of cells ] and can be adjusted everytime by means of 'remote control' [ also shown by transplantations ].
To the question as to the material nature of the gradients no definite answer can yet be given. However, many experiments support the supposition that each one of both the two gradient systems is characterized by special metabolic activities, expressing themselves, among other things, by the reduction of pigments or by the reaction upon enzyme-inhibiting compounds. Recently one has succeeded in isolating an animalizing as well as an vegetalizing substance from egg cells. Probably it here concerns proteins of different natures, influencing morphogenesis.
The embryos evolve stripes (See Figure below), each segment representing a part of the body that will thenceforth follow a different developmental pathway -- to become, say, the upper thorax or the head. The appearance of these segments in fruit fly embryos is thought to be related to the concentration of a protein called bicoid, which increases gradually from one end of the developing egg to the other. The concentration of the bicoid protein provides a signal that "switches on" genes at different points along the body axis, dividing the embryo into a sequence of segments which will develop into different parts of the body. This process of differentiation of regions in an initially uniform embryo is called morphogenesis -- litterally, the genesis of shape -- and the bicoid protein is said to act as a morphogen, which is to say, a determinant of body plan. Exactly how the striped pattern arises from a simple gradient in bicoid concentration is, however, a story that is still being unraveled.

Figure above : In the early stages of development, fruit fly larvae develop striped patterns. The regions of different shade represent parts of the embryo that will subsequently follow different developmental pathways. A gradual variation in the concentration of a morphogen called the bicoid protein from one end of the embryo to the other provides the chemical signal that stimulates this patterning.
(After BALL, 1994)
Gradients can be influenced by chemical substances. Say HADORN & WEHNER, p.191/2 :
If one treats eggs or cleavage stadia of sea-urchins with lithium acids (Li+) [ emphasis added ] , then they develop markedly vegetally. Large parts of tissue that would [ otherwise ] have formed animal organs [ in contradistinction to vegetal organs ], now develop into endoderm. In this way a monstrum develops mainly consisting of gut parts. Rhodanide ions (SCN - ) show precisely the opposite effect (opposite with respect to lithium acids). They inhibit the vegetal gradient, resulting in the egg developing extremely animally. This can be pushed so far that whole eggs only form ectodermal vesicles with stereocilia (as in the isolate of (d) in the Figure given earlier ).
Li+ and SCN - also act antagonistically on the segmentation of the roof of the primitive gut in amphibians [...]. Li+ suppresses the development of the chorda to the benefit of abnormal large myotomes [ muscle parts ], while SCN - results in a giant chorda at the expense of the myotomes. Corresponding antagonistic effects of Li+ and SCN - is demonstrated with respect to the development of eyes and head in octopusses [ or squids, or cuttle-fishes, which is not clear from the text, which uses a general term. ]. Thus material gradients, as they are determining with respect to the whole organization of the embryo of echinoderms [ among which the sea-urchin ], can be supposed to play a role in the differentiation of blastemas [ patterned differentiation ] in many other animal groups. Anyway, also the regional differentiation in the neuro-ectoderm [ anterior-central-posterior ] of the vertebrates can be considered an interaction of mutually overlapping, antagonistic material gradients of trunk and head inductors [...].
Position-information (revisited).
About position-information, as a way of explaining the patterned spatial distribution of organic material, especially of organ primordia, we already spoke on several occasions, and quoted (earlier, in the previous document ) from HADORN & WEHNER, p.192.
Let us repeat this quote.
In many blastemas [a group or patch of not yet differentiated cells] characteristic patterns emerge in which each one of different cells shows a determined direction of differentiation. Thus, for example, on the integument [outer skin] of insects, bristles, hairs or scales stand off at regular spatial intervals in a definite array of distribution. How is it that certain individual cells at certain positions, are determined to undergo a very special development, which is entirely different from the direction of differentiation of the surrounding neighboring cells? [ The patterns of bristles on insects are not necessarily periodic -- allowing an explanation involving repellence -- but nevertheless show genuine and definite patterns.]It is clear that the concept of position-information is closely related to the concept of concentration-gradient systems. In fact the former is the more general notion. A concentration gradient defines the location of a cell by means of a concentration profile (of several different substances). So this is a special case of position-information.
A possible supposition, backed up by experimental results, is that in homogeneous blastemas first certain centers of inhomogeneity are formed, that constitute a kind of blueprint. The individual cells, then will, depending on their position, react to a 'positional information' coming from this blueprint, resulting in different differentiation. In its broadest meaning one can perhaps interpret the development of cells in the sea-urchin embryo as the result of position-information too. In this case, however, it is necessarily implied that the whole egg system co-determines the determining properties of a certain location.
COHEN, J. and STEWART, I., The Collapse of Chaos, 1994, write about position-information (positional information) the following, at p.75 of the 1995 edition :
A lot is known about how genes produce proteins, but there's much more to making an organism than just throwing together a heap of different proteins. In particular, they have to get to the right place at the right time. The developmental biologist Lewis Wolpert has developed a theory of "positional information" to bridge the gap between chemistry and form. The idea is that certain key chemicals can be distributed in the developing organism so that, for instance, the concentration is higher at one end than at the other. Particular cells in the organism can then use that chemical to work out where they are, and then look up their DNA instructions to find out what to do. In this way the same primal cells can develop into different kinds -- muscle, kidney, nerve. It is as if the organism provides its cells with a map (the differences in chemical concentration) and a book (DNA). You look at the map to find out where you are, and then at the book to find out what to do. This is a very flexible system, in principle similar to the way audiences in big sports stadiums can create pictures by each holding up a colored card. To get it right, you have to know where you are sitting and what card to hold up. The main problem with Wolpert's theory is that in some respects it is too flexible. It allows more variation than actually occurs. You can make any picture out of colored cards.The critique, expressed in the last part of the quote is, according to me, a rash one. In this quote we read :
" Particular cells in the organism can then use that chemical to work out where they are, and then look up their DNA instructions to find out what to do."
Well, it is clear that "look up their DNA instructions to find out what to do" is directly implied by "to work out where they are". To a cell, being at a certain place in the embryo, is having its cytoplasm to be in a certain state, which directly causes the activation of certain genes (leading to the particular differentiation of that cell). So because of this implication the system is certainly not too flexible.
By the way, in the above quote we saw how closely related the conceps of positional information and gradient system are.
A gradient system, or, more generally, a position-information system, is, indeed, a kind of map, i.e. it is a blueprint or predesign of the final spatial organization of an organism, or of at least the main features of such an organization. And again it is clear that an organic lattice, grown into a crystal as a result of non-equilibrium crystallization, can fulfil such a function, while having the advantage of originating spontaneously (with parameters set by the DNA code) from something initially not spatially organized in the relevant respect, and (having the advantage) of being robust. Moreover, it can provide a much more detailed morphology of the predesign than any gradient system, in my opinion, can accomplish. The details of this predesign-morphology must directly represent chemical and/or other differences, which are -- or, later in development, trigger -- definite states of the cytoplasm, which in turn trigger the activation of certain genes, resulting in the cell's differentiation.
Also in plants one assumes, in many cases, the presence of gradients with critical concentration values. The distribution of epidermal pores in leaves proceeds according to mutual inhibition, and results in a 'lattice' of pores (but here, because -- I assume -- of many intervening processes) of an irregular kind ( NULTSCH, W., Allgemeine Botanik, 1974, Dutch edition, Part 2, p.173/4). See next Figure.
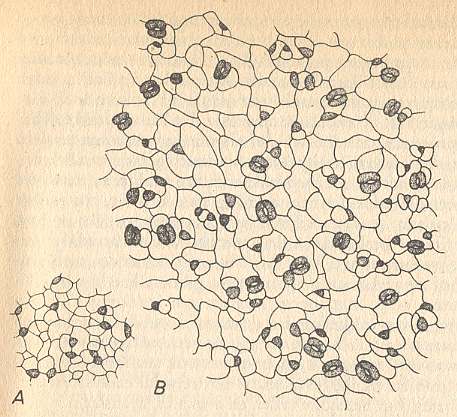
Figure above : Epidermis of Alliaria petiolata.
A : Very young leaf on which, after cessation of the embryonal division activity, epidermal pore primordia are being initiated at certain distances from each other.
B : Older leaf. Here the epidermal pores that were initially laid down, have come, as a result of continued surface growth, to lie so far apart that the areas of inhibition do not all-sidedly touch each other anymore. New pore primordia are being initiated in the open spaces that have so originated.
(After NULTSCH, Allgemeine Botanik, 1974, from BÜNNING and SAGROMSKY.)
Also the patterns, in this case regular, of spines (or spine tufts) on cactusses, especially in Opuntia, could possibly be based on such a mechanism. See next Figure.

Figure above : Opuntia microdasys.
(After PERL, P., editors of Time-Life and FERORELLI, E. (photography), Dutch printing, 1989.)
Especially in the case of higher plants with their typically branched appearance, the assumption of an underlying non-equilibrium -- and consequently its tendency to branching -- organic crystal is especially significant.
Metamorphosis
HADORN & WEHNER, p.193 (Still Part I of the Dutch edition) define metamorphosis as follows :
By [ the term ] m e t a m o r p h o s i s we mean the totality of processes, by virtue of which the larva transforms into an adult animal.They then further elaborate :
This change of form can proceed either more or less gradually (for example in lobsters), or require a complete reconstruction of all kinds of organs. For example a radially-symmetric sea-urchin is, in a very complex way, constructed from a bilaterally-symmetric pluteus larva (See (c) in the Figure given earlier ). During metamorphosis the structures, specifically adapted to larval life (organs involving cilia, filtering devices, digestive organs) disappear. They are replaced by new organs that are required by the adult animal (copulation organs and brood-care facilities). At the same time also the animal's behavior changes.Certain insects (Holometabola) undergo a strong metamorphosis as they pass from larva to adult insect. HADORN & WEHNER, p.194, write the following about these insects.
The larvae (maggots, caterpillars, chafer larvae) do not in any way resemble the adult stadia (flies, butterflies, cockchafers). The reorganization takes place during the pupal stage, in which no food is taken up, and which can last days, months or even years. During the embryonal development at first only the larval organs differentiate. Other cell lines remain undifferentiated during the whole of larval life. They lie concentrated in small clusters or in large imaginal discs [...]. After pupation the larval organs are taken apart. The remains of them are are being digested or dissolved by fagocytes. Only now the cells of the imaginal discs begin to differentate : they (i.e. each of them) produce certain parts of the adult insect body. The process of this development is regulated by hormones [...].It is clear that during metamorphosis the organic lattice, if present (as asumed), is formed anew, i.e. the old non-equilibrium crystal is replaced by another one, which is morphologically different and represents the predesign of the adult animal.
Regeneration
By the term r e g e n e r a t i o n is meant the replacement of parts of an individual organism that have been lost. Evidently this is a phenomenon quite different from the primary embryonal development. It is not the epigenesis of structures but their regenesis.
Here we do not discuss physiological regeneration (like the replacement of hairs in mammals, for instance), but only healing regeneration.
Let's quote HADORN & WEHNER, p.197/8 ((still of) Part I of dutch edition 1977) on this subject :
During healing regeneration a replacement of body parts takes place which were lost by accidental injury (or as a result of experimental interventions) or which are lacking as a result of a natural fragmentation during vegetative reproduction [...].It can be argued that one of the factors responsible for the possession of a regeneration potential, as it is the case in animals, and especially in plants, lies in the organic lattice : It is well-known that crystals have the ability of wound healing, i.e. after a piece has been broken off, they regenerate as soon as they are returned to their proper growing conditions.
On a smaller scale in fact regeneration takes place in all cases of injury (wound healing). But regeneration can also take place with respect to large parts of the body. In a large number of arthropods, fish and amphibians, whole limbs are being restored. Lizzards, for example, replace the whole tail.
In many invertebrates not only the whole is capable to restore the part that is lacking, but a part can also regenerate into a complete new individual. Star-fishes not only restore arms that are lost : in many species one single arm can regenerate also the central field and the remaining four arms. Single segments of annelids (for example earthworms) not only develop a new anterior end of the body with mouth, anterior part of gut and brain, but also a new posterior end with rectum and anus. In planaria (flatworms) 1/100, in fresh water polypes even 1/200 part of the body can still develop into a complete new animal. Such small isolated fragments are not able to take up food, because a mouth is lacking. Before any growth can take place, first of all a complete animal must be reconstructed from the available cell material. This is accomplished by fusion into large organ parts and by reorganization of cell groups. And only such a morphallaxis results in a normally proportioned, but reduced individual.
In general one can say : the higher the level of organization of an animal, the weaker its regeneration potential.
This concludes our exposition of morphogenesis in organisms (embryology) insofar as it is relevant to the theory of organic lattices.
In the next two documents we will prepare to systematically trace the existing analogies obtaining between crystals and organisms.
e-mail :
To continue click HERE for further study of the Theory of Layers, Part XXIX Sequel-13.