
(Takes a little while loading the images)
e-mail :

Sequel-11 to the Summary and Evaluation (of the documents XVI -- XXVIII concerning the promorphology of complex-shaped two-dimensional crystals).
This document (Part XXIX Sequel-11) further elaborates on the analogy between crystals and organisms.
Biological Morphogenesis and Organic Lattices
Introduction.
Shape, Symmetry and Promorph of an organism are the result and expression of some underlying global vectorial matrix, which we have called the Organic Lattice of the organism. But Shape, Symmetry and Promorph as we see them in adult organisms are perhaps not directly caused by such a lattice, but only indirectly. I mean, that the mentioned lattice only emerged in early embryonic development, providing a vectorial matrix for the differentiation and organized migration of cells in the developing embryo. And, then, one might speculate, as soon as concrete vectorial patterning is materialized and more or less complete, the organic lattice degenerates, because now the actual structures that have developed under its (vectorial) guidance, take over the function of localization medium. However, this need not necessary be so. It could be that the organic lattice persists during the whole life of the organism. But, be it as it may, if we try to find such a lattice, we must first of all look for it in the embryological development of (multicellular) organisms.
Let us, to begin with, show a developmental sequence that can be observed in the morphogenesis of the larva of a zygopteran dragonfly (in Dutch : waterjuffer) of the genus Agrion, i.e. the development from the egg-stage up to the fully-grown larva (which then crawls out of the water and transforms into an adult dragonfly).
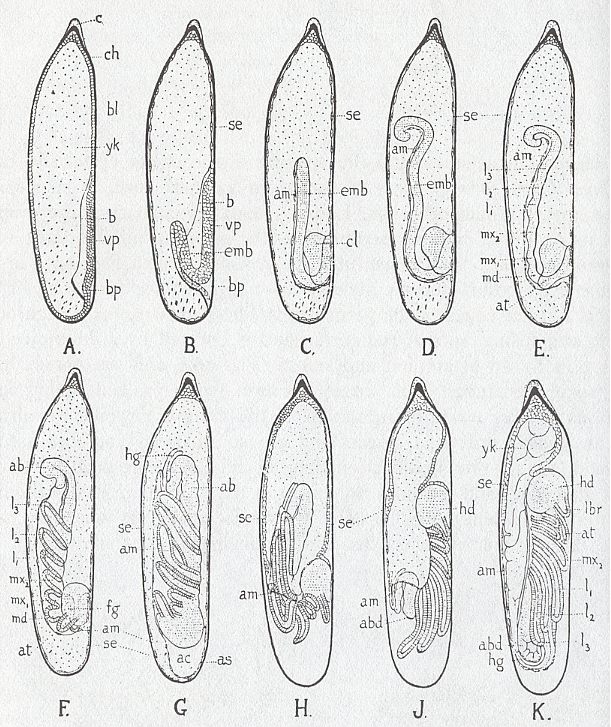
Figure above : Stages in the embryonic development of Agrion (Insecta, Odonata, Zygoptera). Lateral view, ventral surface to the right.
A, Formation of ventral plate. B--D, Invagination of embryo in yolk. E--G, Formation of appendages. H--K, Rupture of amnion and reversion of embryo. (To see all lettering denoting substructures, move horizontal scroll bar if there is any). ab, abdomen. ac, amniotic cavity. am, amnion. as, union of amnion and serosa. at, antenna. b, lateral border of ventral plate. bl, blastoderm. bp, blastopore. c, cap or pedicel. ch, chorion. cl, cephalic lobe. emb, embryo. fg, stomodaeum. hd, head. hg, proctodaeum. l1--l3, legs. lbr, labrum. md, mandible. mx1, first maxilla. mx2, labium. se, serosa. vp, ventral plate. yk, yolk.
(In RICHARDS, O.W. & DAVIES, R.G., Imms' General Textbook of Entomology, Tenth Edition, 1977, Volume I. From TILLYARD, Biology of Dragonflies, after BRANDT.)
Without looking to the details, we see that from a more or less isotropic egg mass a definite directional, and thus anisotropic, structure (the larva) develops.
With respect to evaluating -- for the benefit of (i.e. for testing) our theory of organic lattices -- facts and hypotheses as they are present in developmental biology, we totally rely on the writings of authorities in the field. So the following discussion consists mainly of quotes.
It is important to realize that all cases of embryological development show a bewildering complexity of process, transformation and organization, which are far from being totally understood, as to their 'mechanisms'. But in setting up a theory of organic lattices (or of a comparable theory) it is a fair and sound strategy that we only then resort to (the assumption of) different ontological domains ( like that of the Implicate Order of BOHM, the fields of SHELDRAKE -- although the latter maintains that his fields are immanent --, or the entelechy of DRIESCH ) when they seem unavoidable. And this means that the organic lattice must be considered in biochemical or physiological terms. It is important to provisionally refuse to let appreciation of the wholeness of the organism become an excuse for avoiding rigorous causal analysis. There must be, at least to begin with, an explicit insistence that the unity of the embryo is not a mystical absolute, but can be understood by attention to the organization of distinct processes. This means that the holistic properties as seem to be present in the generation of (snow) crystals, and which could be, mutatis mutandis, transferred to the development of an organism, must, for the time being, remain in the background of the discussions concerning the organic lattice, while the great form-potential of non-equilibrium crystallization must come to the foreground of those discussions. Nevertheless the whole plays an important role in organisms : One of the functions assigned to the organic lattice is that of a medium of localization. The part that a cell plays in constituting the whole is a function of its spatial position within that whole.
Our concept of organic lattice has, until now, been developed in such a way that the result has, more or less, shifted away from that of a genuine crystal lattice. It has become more akin to that of some sort of field. And this could bring us nearer to the fields postulated by NEEDHAM, or WEISS in the 1930s, or to the morphogenetic fields of SHELDRAKE, postulated in the 1980s. Adherence to the latter fields, i.e. a more or less identification of the organic lattice with a field of SHELDRAKE, can only be successful when for the latter a strict immanence in physical matter is insisted upon, and when, consequently, an ontological equivalence between these fields and physical matter is upheld. Only then such a field is comparable with a (para)crystalline lattice residing in an organism.
Let us quote HARAWAY, D., Crystals, Fabrics, and Fields, Metaphors of Organicism in Twentieth-Century Developmental Biology, 1976, p.124, on the idea of immanent (and thus biochemical) morphogenetic fields, developed in the 1930s by Joseph NEEDHAM and others [comments of mine between square brackets].
Needham believed that fields were distinguished from simple geographic regions of the embryo by three criteria : any given point within the field force had to possess a given quality, a given direction, and a given intensity. Fields were judged in terms of instability and successive equilibrium positions [ like we see it, in fact, also in crystallization, where the particles move into stable positions ensuring a lowest-energy configuration]. Waddington reasoned that "a field is a system of order such that the position taken up by unstable entities [ I assume that here not the entities themselves, but their position within a system of them, is initially unstable.] in one portion of the system bears a definite relation to the position taken up by unstable entities in other positions". Behavior of cells was, within certain boundaries, a function of position within the whole. Many of the organizational forces of fields were "to some extent on a suprachemical level" (p.71, 1937, in a paper concerning the chemical aspects of morphogenetic fields). Needham saw the work of D'Arcy Thompson (1917) [especially the Chapter On the Theory of Transformations, or the Comparison of Related Forms, using deformable coordinate grids], as a significant but chemically unspecified root of legitimate field thinking. Needham hoped that his work on the evocator [playing a role in the phenomenon of induction in embryological development], coupled with a sophisticated understanding of proteins and paracrystalline structures, would give a firm biochemical content to morphogenetic fields. One goal was to account for the multiplicity of biological fields in contrast to the relatively few kinds of physical fields [ The condition of "multiplicity of biological fields", i.e. their diversity of kinds, is, I think, well fulfilled by the concept of organic lattice].And with fields and the like we are in fact adhering to one or another brand of organicism when interpreting complex systems. The latter can be interpreted according to reductionism, holism, vitalism, mechanism, or organicism.
Our ensuing expositions will try to comply with this version of organicism as best they can.
Thoughts about some underlying paracrystallinity in the developing embryo.
HARAWAY, D., in her already cited book Crystals, Fabrics, and Fields, Metaphors of Organicism in Twentieth-Century Developmental Biology, 1976, discusses the working-out of the paradigm of organicism within embryology.
Organicism is an approach to the study of biology which views organic form as its central object and the growth of organic form as its central problem. So organic form is not seen as something evident, but as a problem. Organicism is neither "vitalism" nor "mechanism", which have long been accepted as the rival commitments of biologists. It may be viewed as an alternative to both. It emphasizes levels of organization in their own right. It does not invoke special metaphysical entities (as in vitalism) to account for morphogenesis with its phenomena of regulation and regeneration.
In her book HARAWAY discusses a number of outstanding embryologists, working in the first half of the twentieth century, the work and ideas of which happen to be very relevant to a discussion of our theory of organic lattices. Therefore we will quote many passages from this book (which themselves often contain quotes from the works of the mentioned embryologists). Among these embryologists was Ross G. HARRISON, who, based on his experimental research, speculated about the global mechanisms of organic morphogenesis in terms of some version of a crystal analogy. A repeat pattern of a structured protoplasm is the form the crystal analogy finally took for HARRISON.
In discussing HARRISON's last major experimental paper (HARRISON, ASTBURY, and RUDALL, 1940), HARAWAY, p.81--82, relates the folowing [comments of mine between bold square brackets] :
The paper is a gold mine of forward-looking speculation and attempts at experimental demonstration. The work involved X-ray diffraction photographs of several living embryonic tissues from chick and newt : neural plate, neural tube, ear ectoderm, notochord, and yolk. It was hoped the photographs would support the molecular orientation theory of tissue polarization.HARRISON advovated a molecular basis for the fundamental principle of vital organization. CHILD, on the other hand, thought of functional (metabolic) properties as the basis of axial differentiation and chemical gradients. HARRISON criticized CHILD, noting that "such gradients may well be an expression of the polarity rather than its cause" (HARAWAY, P.89).For some time past, evidence has been accumulating, mainly from transplantation experiments, that certain embryonic systems are at first isotropic [i.e. no preferred directions] with respect to their future differentiation and only gradually become oriented and polarized. This secondary condition has no immediate outward expression, and since the systems in question are known to be equipotential, the changes observed must be based upon some finer structure of protoplasm, which is probably of a molecular order of magnitude. [p.339]In the case of tissue that was still isotropic, any molecular elements responsible for polarization [not necessarily optical, but definitely structural] would not yet be oriented. Molecular rearrangement at the time of polarization might be detectable by X-ray crystallography, but technical difficulties prevented a clear answer. The photos did not show any definite orientation but gave only a ring pattern characteristic of a disoriented protein. The results were not taken as a refutation of the hypothesis, because of the difficulty of detecting a postulated, oriented, noncrystalline array of globular molecules in living tissue [ I assume that "noncrystalline" in the present context must be understood as paracrystalline, i.e. a state where the molecules are oriented in some common direction, but do not form a genuine crystal lattice, because of the absence of rigorously repeated equal distances of motifs in each of the three space directions, i.e. the absence of translational symmetry, which is essential for a structure to represent a genuine lattice. ]. It was extremely difficult to study protein structure even with true crystals. Detection of a "paracrystalline" state, that is, a condition in which the elements had freedom of movement along one or more axes, was exceedingly unlikely. It was a significant victory simply to get X-ray pictures from living material. The basic conception underlying the trial remains potentially valid, and perhaps a contemporary repeat of the work would be successful.
A second important method was applied to embryological problems in the 1949 paper : optical examination with a polarizing microscope. Orientation in biological tissue had been studied before with polarized light (Schmidt 1924). Birefringent material was seen in Harrison's material, especially at cell membranes. Harrison noted that their work was preliminary and should be followed up systematically. "Especially should the cell boundaries be examined thoroughly, for it is there, perhaps more than anywhere else in the cell, that we may expect to find the seat of directive forces" (Harrison, Astbury, and Rudall 1940, p.355). A glance at current journals in cell and developmental biology reveals the appropriateness of the admonition to study cell membrane systems in relation to form problems.
Harrison speculated that primary polarization of [the] egg arose from alignment of the protein molecules oriented as a function of their dipole character. The opposite chemical properties of the two poles would result in different chemical reactions and electrophoretic transport of charged molecules to different sectors of the egg. Field properties would be set up by forces arising from the chemical structure of the cell, and material gradients could be one expression of the situation. Substances produced under direction of the genes would be localized in the structured cytoplasmic milieu, resulting in different chemical reactions taking place in various regions. New local centers of activity with a potential to differentiate into subfields were thus established. Side chains of the oriented protein molecules were envisioned as the locus of specific chemical activity (an intimation of proteins functioning simultaneously as enzymes and structural units), and relative reaction velocities would account for greater and greater local diversity of action. As the egg continued to divide, the above processes accounted for accompanying local diversification of cells. But each cell of an organism would retain a characteristic cytoplasmic asymmetry within which gene-directed metabolic processes occurred. The idea that each cell contained the full genetic endowment was broadened to include the possession also of a basic cytoplasmic structure. Within this framework, differentiation, dedifferentiation, and determination took place. There was no need to postulate neoblasts, any more than there was a need to search for the irreversibly determined, differentiated cell. No elements of the cell -- genetic, metabolic, or cytoplasmic -- were conceived as either inactive or unstructured.HARRISON is one of those that laid the foundation for an organicist molecular biology. HARAWAY, p.92, reports :
Significant cell movements such as gastrulation and neurulation were explained by cell-shape changes due to the action of crystallization forces resulting from changes in molecular configurations during chemical reactions.(NOTE of HARAWAY : The recent demonstration that microtubules and microfilaments involved in cell-shape changes are necessary for the folding of the neural plate to occur in Xenopus embryos is ample confirmation of Harrison's speculations (see Karfunkel 1971).)
"Differentiations are in a sense, then, the byproducts of cytoplasmic activity and are accompanied by movements involving change of form" (1937, p.373). At no point in the scheme does a single cause determine a single effect. Systems of oriented processes differentiate into subsystems coordinated by their mutual functioning. Many lines of Harrison's bold theory remain unconfirmed, but his approach still constitutes perhaps the richest contribution to a coherent theory of development. The concept of fields resulting from structured activity is a typical organicist resolution of the field--particle polarity. The resolution rests, essentially, on going beyond structure--function dichotomies.
in the sense, I think, of desired grain s i z e, i.e. the size of the 'unit cell' ( = repeating unit) of the paracrystalline lattice.]. "The existence of the equipotential system necessitates, in fact, the assumption of some sort of molecular hypothesis for the representation of adult form in the germ . . . . In other words, it is the intimate protoplasmic structure that underlies symmetry" (1921a, pp.87--88).
The fundamental organizing insight in Harrison's approach to development was his conception of the role of protein molecules. Proteins were important because they gave him a hold on a source of asymmetry of the desired "grain" [
He began thinking in molecular terms very early (at least by 1921) and built molecular models in his Osborn laboratory constantly. He was influenced by H. Przibram [wrongly spelled "Prizbram" by HARAWAY ], who referred symmetry and polarity relations of organisms to the arrangement of particles in a space lattice. However, Przibram had little appreciation of an organicist approach, and Harrison's originality resided in his use of the crystal analogy within such a perspective, with a full "realization how vastly more complicated these relations are in organisms than in crystals."Indeed, PRZIBRAM thought of more or less genuine crystal lattices constituting an organism, which would, however turn organisms into crystals, which they obviously aren't. In our theory of organic lattices the lattice only involves certain chemical constituents of the organism, i.e. the lattice is not (identical to) the organism, but resides in the organism. So the 'classical' crystal analogy does not hold. Our theory only postulates a paracrystalline background and introduces, as a paramount aspect, the non-equilibrium crystallization of the material of the organic lattice, resulting in a detailed morphological and vectorial matrix of at least the early embryo.
The fifth lecture of the so-call Silliman Lectures, held by HARRISON in 1949, was called " The symmetry of organisms". The essential problem, however, was not true symmetrical organization but residual asymmetry of organisms. HARAWAY (p.98--99) reports the following :
In the lecture Harrison reflected on the discovery of optical isomers by Pasteur in 1848. The discovery of molecular asymmetry had been a principal turning point in the relation of chemistry to biology. Optical isomerism provided Harrison with the analogy for his own version of the organism--crystal relationship. But it was clear from work on coiling in snails that molecules with the same asymmetry could produce organisms with opposite, mirror-image symmetry. Therefore it was too simple to postulate a one-to-one relationship between molecular pattern and protoplasmic structure. The clue to the dilemma lay in the suspected presence of a special type of crystal organization in organisms : liquid crystals. Such labile structures may change their orientation. "They are, in other words, para-crystalline and are often fixed in one or two dimensions instead of all three." In this context Harrison interpreted his work on the axes of symmetry in limb, ear, and gill systems of the newt. Molecular models showing successive substitution of four groups bound to a central carbon atom, initially very suggestive, were inadequate to the complexity of the organism. Rather, "the model showing progressive orientation of asymmetric molecules will explain the facts better". It further gave a means of understanding twinning across reflecting planes, a perplexing phenomenon encountered continually in the course of limb and ear experiments. "Twin crystals are very common and in cases of asymmetric forms, they frequently are the mirror image of each other . . . . This phenomena [sic] then speaks very strongly for the crystalline or paracrystalline nature of the living tissue making up the ear vesicle" (Silliman Lecture 5, pp.3, 8, 7).It is clear that our organic lattices (and of course also those implied in the organicist version of the crystal analogy as held by HARRISON) do not suffer the mentioned deficiencies. Moreover, non-equilibrium crystal growth can account for differential growth.
Harrison concluded his important observations with a simple but far-reaching statement.The significance of all this detail, which I fear may have bored you, is that it gives crucial evidence that these two quite different systems, the ear and the forelimb, are made up of units arranged in a repeat pattern which becomes definitely more and more oriented. In this respect they are similar to the egg . . . . In fact, that kind of organization seems to pervade all living matter and will have to be reckoned with in any theory of the organism. [pp.10-11]In the last lecture, "Development and growth in complex systems", the limitations of classical [italics mine, JB] crystal analogy, rather than the power of the organicist version [of the crystal analogy], were explored. Crystals grow by accretion [not so perhaps in the case of liquid crystals], and addition to each axis is proportional to axial relations of the lattice [ This is not entirely correct : growth rates normally depend not only on the geometry of the lattice but also on the atomic aspect (corresponding with a given crystallographic direction) as it is presented to the nutrient environment. Further, we know from snow crystals that the habitus of crystals can change, when certain conditions change. ].
"Growth of organisms is more complex, since they grow internally and are made up of many different components with different rates of increase" (Silliman Lecture 6, p.1).
Further development of the theory of organic lattices and their relation to embryology.
We have developed a crystal analogy resulting in the concept of organic lattice. But it surely is a hard problem to enrich this analogy with concrete content.
If an unambiguous point or location of reference, and three axes going through that point, can be assumed to exist in the developing embryo, then the organic lattice has the nature of a coordinate grid and can, on the basis of that, serve as a localization medium, in the sense that cells 'know' where they are within the embryo. The detailed macroscopic morphology of such a grid or lattice, as a result of non-equilibrium crystallization, can also serve this purpose. When a cell knows where it is, it can open the appropriate section of its genome, resulting in the production of special proteins, in turn resulting in the cell's differentiation.
The detailed structure of the organic branched crystals also serves to be a form-matrix for the (penultimate or ultimate) morphology of the given organism. But the assumption of the generation of this detailed structure, as the expression of the crystal's form-potential, is based on insights taken from the mode of growth of solid (or more or less softened, swollen) crystals. And this growth is by way of apposition, which means the accretion of particles from without, i.e. from the nutrient environment, onto the surface of the growing crystal. And this seems to pose two difficulties for the crystal analogy to hold.
First, the initially formed structures (in true crystal growth) do not change anymore as soon as they are laid down. They remain as such within the growing crystal. Changes of morphology only occur by additional external deposition of new elements. This is definitiely not the case in organisms.
And this latter process brings us to the second problem (for the crystal analogy to hold). Organisms do not grow by apposition, but by intussusception : They take up food into their interior and then, after having it transformed (assimilated), adds it to its bulk mass from within.
So although crystal growth, and especially (controlled) non-equilibrium growth, is able to produce organic shapes, it doesn't produce them in the way organisms do. But despite of all this, the crystal analogy must have some legitimacy, as the work of HARRISON and others have shown, and, also, we cannot believe that life, which is so dependent on form, hasn't incorporated (appropriated) the great form-potential of non-equilibrium growth into its processes. And, moreover, without active involvement of one or another lattice, we have the problem of regenerative ability and vectoriality in embryological development (and also in the development of singly living cells like those of many Radiolarians).
But, in fact, we implicitly had already solved the problem of crystal growth versus organic growth in our version of the crystal analogy : We already established earlier, that while a true crystal IS its lattice (provided with chemical motifs), an organism is not : An organism HAS a lattice (also provided with chemical motifs), and this lattice was assumed to function more or less independently (thanks to assumed compensating reactions) with respect to the remaining chemical 'machinery'. So in this context it is only natural to expect that this lattice grows by apposition (of material from the immediate vicinity of the embryo), just like a true crystal does, while the rest of the organism grows by intussusception and assimilation.
The second difficulty that was stated above, namely the fact that this appositional way of growth implies that initially laid-down structures do not change anymore during the course of further growth of the crystal, had also already been solved, by assuming that the lattice can be deformed, without ceasing to be a lattice (accomplished by the action of assumed compensatory reactions). Indeed we see in embryological development the phenomena of extension (growth), followed by deformation. So the whole embryological form-development consists in a lattice (provided with chemical motifs) that grows by apposition (and where the elements are recruted from the embryo's immediate environment -- nutrient medium of growing crystal), while it is at the same time deformed by growing processes of the embryo (which are of an intussusceptional nature). In this way we must see that the lattice and the symmetry of the embryo, as they are in themselves, are not transformed by deformation, and that, accordingly, the localization medium is unaffected, despite the fact that at the same time the lattice is transformed with respect to straight three-dimensional space, even such that, from this viewpoint, it is not a true lattice anymore, because strict translational symmetry is lost. So the embryo and its organic lattice is geometrically an independent entity, it is a non-euclidean spatial object embedded within an euclidean space. The lattice is chemically independent by assumed compensating reactions.
So we must admit -- but this was clear from the outset -- that non-equilibrium crystallization is not, and cannot be, the sole 'method' of form-generation (morphogenesis) in organisms. It proceeds alongside with deformation, resulting from intussusceptional assimilational growth and changes of cell shapes, while cells differentiate, generally according to their location within the embryo, where this location is recognizable, i.e. defined, by the features of the existing landscape which is the result of non-equilibrium growth of the organic lattice. While internal deformation does not (as assumed) change the organic lattice-as-it-is-in-itself, it does change, even significantly, the morphology of the embryo. External deformation, i.e. change of posture of the embryo as a whole, not only does not (as assumed) change its organic lattice, but also does not change the embryo's morphology and structure.
It could further be assumed that the organic crystal, as it resides within the organism, and also within the developing embryo, is a mixed crystal, which, when becoming unstable -- with respect to holding more or less different motifs at its lattice points (as is the case in mixed crystals), demixes, resulting in chemically different zones or areas.
Position-information and differentiation in a developing embryo.
HADORN, E., & WEHNER, R., in Allgemeine Zoologie, Dutch edition, 1977, p.192, write the following (translated into English) :
In many blastemas [a group or patch of not yet differentiated cells] characteristic patterns emerge in which each one of different cells shows a determined direction of differentiation. Thus, for example, on the integument [outer skin] of insects, bristles, hairs or scales stand off at regular spatial intervals in a definite array of distribution. How is it that certain individual cells at certain positions, are determined to undergo a very special development, which is entirely different from the direction of differentiation of the surrounding neighboring cells? [ The patterns of bristles on insects are not necessarily periodic -- allowing an explanation involving repellence -- but nevertheless show genuine and definite patterns.]In this more or less recent theory one speaks in terms of blueprints or similar terms that obviously relate to a pre-pattern accounting for a pattern. As such it is certainly not an elegant scheme of explanation, but the basic assertion is interesting : many specific cell differentiations depend on the location of such a cell within the relevant part of the embryo. And we can generalize this, to cell differentiation direction being -- in many cases -- dependent on the location of the cell within (just) the embryo. But of course the cell must then 'know' its position. It must be affected by location-specific agents. One way of accomplishing this could be -- as one assumes -- by way of chemical concentration gradients. The substance of such a gradient is called a morphogen. As far as I know, only two such putative morphogens are actually detected in animal embryos : retinoic acid in the chick limb bud (SHELDRAKE, The Presence of the Past, 1988, p.331 (note 28)), and the bicoid protein in fruit fly embryos (BALL, Designing the Molecular World, 1994, p.312). Some critical concentration values then determine how a cell responds, by activating a certain part of its genome, resulting in its subsequent differentiation.
A possible supposition, backed up by experimental results, is that in homogeneous blastemas first certain centers of inhomogeneity are formed, that constitute a kind of blueprint. The individual cells, then will, depending on their position, react to a 'positional information' coming from this blueprint, resulting in different differentiation. In its broadest meaning one can perhaps interpret the development of cells in the sea-urchin embryo as the result of position-information too. In this case, however, it is necessarily implied that the whole egg system co-determines the determining properties of a certain location.

Figure above : A sector of a snow crystal.
This particular structure is part of a snow crystal, and snow crystals generally have a six-fold symmetry. One can, however, imagine that the depicted structure is the result of (non-equilibrium) growth of a lattice+motifs that has intrinsic bilateral symmetry (D1) only. And then this structure is no longer just a part of a crystal but a whole crystal.
(Adapted from LIBBRECHT, The Little Book of Snowflakes, 2004.)
Regulation in the embryo, and regeneration of parts in animals and plants.
Our model of embryological and postembryological form-generation -- morphological blueprint resulting from non-equilibrium crystallization, plus deformations of the (organic) lattice underlying the blueprint, and plus location-dependent cell differentiation, where "location" refers to the blueprint landscape -- can also roughly account for regulation and regeneration :
If a certain part of some young embryo is removed (or if a certain part of a plant is removed, or, finally, if a particular part of a certain type of animal is removed), then a number of cells and the corresponding part of the organic lattice get lost (to the (developing) organism). But, when the same (non-equilibrium) conditions are still present, the (organic) crystal will restore its morphology again, and undifferentiated cells grow and divide along with the extension of the regenerating organic lattice. But then these cells will eventually going to react to their position, and will differentiate accordingly, i.e. they will repeat the original differentiation of the lost cells : The embryo regulates its developmental pathway, or (respectively) the organism regenerates lost parts.
Epigenetic versus preformistic development.
Organic morphogenesis is epigenetic, not preformistic, i.e. (in epigenesis) new structures are being developed that were not there before.
But the notion of epigenetic development is not so clear-cut as it sounds. Of course in a fertilized egg we will not find some miniature adult organism, which only has to increase in size. But a preformed adult organism can be present in a more hidden fashion, namely as the definite and total set of genetic instructions, as is assumed by many workers in the field. And after having become more or less convinced that genes alone are not sufficient, we could add the set of dynamical laws and initial conditions corresponding to a set of certain dynamical systems of which the parameters are set by genes and their products. In this way the form of the adult organism is pre-formed, but in a more subtle and hidden sense.
It is very hard to demand that entirely and absolutely new features appear in embryological development that were in no sense already implicitly present in the early embryo or even already in the fertilized egg. This would probably entail the assumption of fundamentally new basic principles of being, in addition to those that were already present. And this connects to the theory of Layers of Being, where the Organic Layer differs from the Inorganic (physical) Layer by the appearance of some categorical NOVUM. In earlier documents of the present Series we have discussed such a layered world. But, as we have said, in developing our theory of organic lattices we try to avoid such assumptions as long as possible. So we start from assuming that the developing egg does not contain an actually existing pre-formed end stage, but only a potential one. And the latter means that the fertilized egg is not supposed to contain nothing that is relevant to the ensuing developmental sequence, and its final result. Indeed, our blueprint, that is a result of the non-equilibrium crystallization of the material of the organic lattice (co-evolving with the extension and ensuing differentiation of cells, and undergoing deformations), emerges from a -- in many respects -- more or less isotropic initial state. But the material, or pre-material, of this state is already present, and has the ability to organically crystallize when conditions are suitable. And this resulting blueprint of the morphology of the organism's final stage is indeed only a blueprint and nothing more. And the cells have the potential to differentiate, in virtue of their genome, of which either this or that part can be opened (i.e. can be made active) depending (generally) on the cell's position within the embryo, resulting in the production of specific proteins, causing a specific type of metabolism corresponding to a certain definite differentiation of the cell concerned.
On the role of genetics in the generation of form in embryological development.
But isn't it so that the complete form of an organism, including all the stages that lead to it, is completely determined by the total set of genes of the given organism? The most viable concept in this respect is the genetic program. But the genetic program, as the total set of instructions encoded in the DNA of a given organism, generally is the same in every one of its cells. Pattern formation insofar as it involves a pattern of differentiation (of cells), must correspond to a pattern of gene expressions : a certain gene expression is the activation of one ore more stretches of the DNA chain, so we have to do with a pattern of different activations across the collection of cells of the given organism. And of course this pattern will change as a result of all kinds of interactions.
A genetic program, just as a given genetic program, lets an organism be distinguished from another organism : It accounts for the stable and repeatable and intrinsic difference between organisms. One of these differences is a difference in morphology. Genes do not create a certain particular morphology of a given organism. It is expected that genes and their products, together with some environmental factors, represent definite 'parameter settings' [ a parameter is a certain magnitude that remains constant during the particular run of a dynamical system] for certain dynamical pattern-generating systems abiding within the organism. So we can surmise that genes and their products ( Where the set of particular genes has been evolved by natural selection of the phenotypes of mutants, and passed on from one generation of organisms to the next.) set off these form-generating systems into a particular direction. Such an action of setting parameters, as is done by the genes and their products, could also be postulated to play a determining role in the formation of the organic lattice (provided with chemical motifs) and in the non-equilibrium growth of this lattice (where the formation and growth of the latter is itself a running dynamical system), resulting in a complex morphology with internal periodic or quasi-periodic (in the case of a quasi-lattice) structure. And we have seen how many different morphologies can be generated this way in the case of the fast growth of the ice lattice. And, as has been said, the resulting morphological structure can as such represent a system of positional information, effecting patterns of differentiation of cells, which patterns are then part of the true morphology of the (developing) organism.
Organic morphogenesis (embryology).
After having the idea of organic lattices further elaborated, it is time to explicitly integrate this idea with morphogenesis (especially embryology), as the latter is actually observed in (multicellular) organisms. However, this attempted integration is only preliminary, speculative, crude and incomplete. It is only meant for setting the stage for a truly extensive and comprehensive investigation on which I myself am not in a position to embark. Further, because this website is not, or is not intended to contain, a textbook on embryology (also because I am not an authority on this very difficult and extensive subject), the reader is referred to one or another existing textbook on embryology or on general biology, or zoology for that matter, if he doesn't feel sufficiently at home in the subject.
Most of the inspiration, playing a role in what comes next, comes from a textbook of general zoology : Allgemeine Zoologie, by HADORN, E. & WEHNER, R. (Georg Thieme Verlag, Stuttgart, Germany), Dutch edition, 1977 (Aula) : Algemene Zoölogie, Volume I. We will refer to this Dutch edition by "HADORN & WEHNER".
In what is about to come the reader will not find a systematic argument which neatly derives the existence of organic lattices from the data of embryology. It is just a collection of notes about the compatibility (or even the necessity) of our postulated organic lattice with certain observational facts or hypotheses that are to be found in embryology.
Let us begin with two Figures that schematically depict some aspects of embryological development. Later we will refer back to these Figures.
Both Figures are about the gastrulation taking place in the amphibian (frogs and the like) embryo. Gastrulation is an invagination process : A certain area of cells of the blastula (which, as a stage preceding gastrulation, is a hollow cyst, the wall of which consists of cells) starts to move inwards, and this process continues till a large part of the cavity of the blastula is filled with invaginated cell mass. Differentiation, accompanied by further cel movements, then takes place, resulting in the rudiments of a number of organs. In the amphibian blastula one can indicate certain definite cel areas that, after gastrulation, end up in several definite organ rudiments of the further developed embryo. In other words, one can draw a fate map on the amphibian blastula. However, judging from the method, decribed in HADORN & WEHNER, p.149, one can do this only after the fact, which, according to me, means that the cells of the blastula are not yet (visibly) differentiated or pre-differentiated. The fatemap only indicates the scheme of invagination and migration of cells and cell patches. So although it doesn't indicate visible differentiation, it indicates definite migration tendencies of the various cell areas (HADORN & WEHNER, p.153/4). And the bilateral (eudipleural) symmetry of the fatemap clearly indicates that a definite vectoriality relating to that of the organism which is about to be formed, is already present in a stage as early as the blastula.
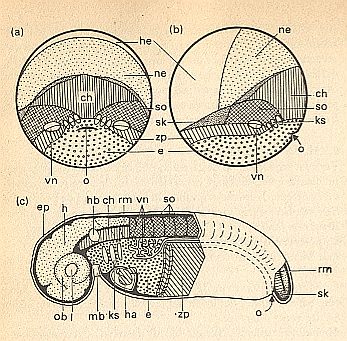
Figure above : (a) and (b) : Fate map ('predesign') of an amphibian embryo ( Triturus) at the beginning of gastrulation.
(a) Hind view, primitive mouth (blastopore) visible as a hollow in median position.
(b) Sinistro-lateral view (i.e. side view from the left).
(c) Organ topography as it is in the developmental stage where the tail bud is visible.
o = primitive mouth (arrow).
Ectodermal germs (primordia) and organs.
Skin ectoderm (he, white) forms epidermis (ep), lens (l), auditory vesicle (hb) and mouth curve (mb).
Neuro-ectoderm (ne, stippled) forms the neural tube and the brain (h), spinal cord (rm) and eye cup (ob).
Mesodermal germs (primordia) and organs.
chorda (ch, vertical hatching), somites (so, crosshatching), lateral plates (zp, oblique hatching), heart (ha), tail bud (sk), pro-kidneys (vn, white).
Endoderm (e, small cirles) forms, at the exterior -- in (a) and (b) -- the vegetal region of the early gastrula, and later forms -- in (c) -- the gut, in which occur gill slits (i.e. elongated apertures, ks).
(After HADORN & WEHNER)
(End of Figure subscript).
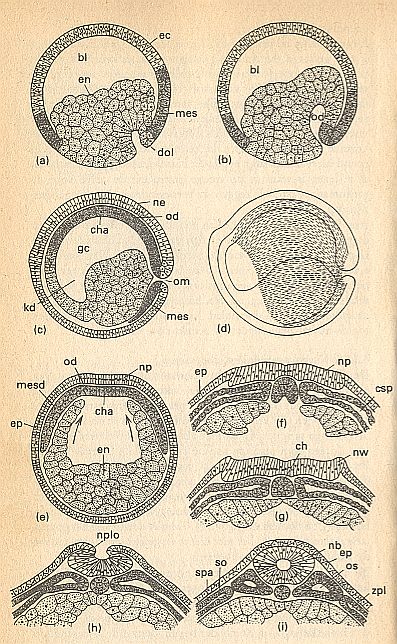
Figure above : Completion of gastrulation and organogenesis in an amphibian germ.
ec = ectoderm (lightly colored), en = endoderm (lightly colored, stippled), mes = mesoderm (dark, stippled).
(a)--(d) Longitudinal sections show the continuing invagination of endoderm and mesoderm :
(a) Early gastrula with a still large blastocoel (blastula cavity, bl). Invagination under the dorsal lip of the primitive mouth (blastopore, dol) begins now.
(b) Midway of gastrulation. At the expense of the blastocoel, a cavity of the primitive gut (od) develops.
(c) Late stage of gastrulation. The mesoderm with the median chorda primordium (cha) lays itself, as the cap of the primitive gut (od), against the neuro-ectoderm (ne). The cavity of primitive gut = gastrocoel (gc) has now reached its maximal size. The anterior part of this cavity is the lumen of the cephalic gut (kd). From the closed primitive mouth (om) ventrally an invagination of the mesoderm (mes) proceeds. The blastocoel is now completely ousted, and the surface of the embryo now entirely consists of ectoderm.
(d) The striped region indicates how much the inturned mesoderm extends across the germ in an early stage of neurulation.
(e)--(i) Transverse sections through the middle of the trunk :
(e) The mesodermal roof or cap (mesd) of the primitive gut now lies between the ectoderm constituting the future neural plate (np) and the gut endoderm (en). It covers the gut cavity, formed by the upwardly growing gut walls (arrows), which have not yet closed the gut cavity (ep = epidermis).
(f) The chorda (notochord) has separated itself, while holding a median position within the roof area of the primitive gut, from the remaining mesoderm. In the mesoderm on both sides of the chorda primordium a coelom slit (csp) develops. The ectoderm of the neural plate (np) is thickened with respect to the epidermal ectoderm (ep).
(g) The rims (edges) of the neural plate now form neural embankments (nw) (becoming the neural folds), the gut cavity below the chorda (ch) is now closed.
(h) The neural folds (nplo) grow toward each other, marking the beginning of the formation of the neural tube.
(i) Neural tube (nb) closed, detached from the epidermis (ep) and sunken in. Mesodermal somites (primitive segments, os) separated from the side-plate mesoderm (zpl). Coelomic walls with external (somatopleura, so) and internal (splanchnopleura, spa) lining.
(After HADORN & WEHNER)
(End of Figure subscript).
During the formation of the blastula from the fertilized egg cell, cell division is dominant, while during gastrulation cell migration (cell localization) is dominant, resulting in the outline of the main body plan.
The next Figure depicts the early stages of development of a sea-urchin egg (And from a developed egg then a bilateral symmetric larva will hatch, which later transforms into the five-fold symmetric adult sea-urchin). Some interesting details of the gastrulation process are indicated :
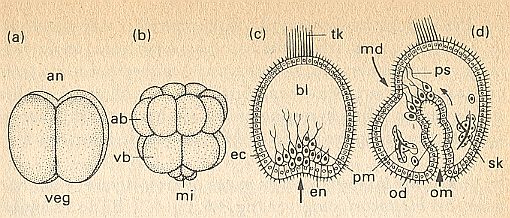
Figure above : Cleavage and gastrulation of a sea-urchin egg (schematically).
(a) 4-celled stage. up, animal pole (an), down, vegetal pole (veg).
(b) 16-celled stage. ab = animal blastomers, vb = vegetal blastomers. mi = micromers.
(c) Beginning of gastrulation. The arrow indicates the direction of invagination of the endoderm (en). ec = ectoderm with cilia and tuft of vibratory hairs (tk), primary mesenchym (pm) with pseudopodia migrates inwardly into the blastocoel (bl).
(d) Late gastrula, just before the mouth breaks through (md). Secondary mesenchym with pseudopodia (ps) in contact with the ectoderm of the mouth edge (md, arrow). The primitive gut (od) lays itself against md (arrow). pm ( = primary mesenchym) secretes skeleton (sk). om = primitive mouth.
(After HADORN & WEHNER)
Organic lattices and organic morphogenesis (embryology).
Above we cited HARAWAY (1976) with respect to the ideas of Ross HARRISON concerning cell movements and folding processes. Let us repeat a part of this citation :
Significant cell movements such as gastrulation and neurulation were explained by cell-shape changes due to the action of crystallization forces resulting from changes in molecular configurations during chemical reactions.The mentioned microtubules (microtubuli) and microfilaments play a role in cell shape, and a change of this shape can entail the formation of specific multicellular structures in the embryo. Let us consult some recent authors. [comments between square brackets].(NOTE of HARAWAY : The recent demonstration that microtubules and microfilaments involved in cell-shape changes are necessary for the folding of the neural plate to occur in Xenopus embryos is ample confirmation of Harrison's speculations (see Karfunkel 1971).)
"Differentiations are in a sense, then, the byproducts of cytoplasmic activity and are accompanied by movements involving change of form"
Cells retain their shape by virtue of an internal 'skeleton' composed of fine tubules (microtubules) whose principal constituent is the protein tubulin [ In fact these tubules are made from a perfect regular checkerboard pattern of units that contain two distinct proteins, alpha- and beta-tubulin (STEWART, I., Nature's Numbers, 1995, p.82)]. Microtubules [. . .] will spontaneously assemble from a tubilin solution provided the ionic composition is correct [. . .], and indeed in living cells they can be shown to be undergoing periodic oscillations between their assembled (polymerized) and disassembled forms. ROSE, S., Lifelines, 1997, p.170.
HADORN & WEHNER (1977), p.153/4, wrote the following about the mechanism of gastrulation and the dynamics of the cell shape, involving microtubuli and microfilaments.
During gastrulation massive cell migrations take place. The transposition of extensive cell regiments into the interior of the germ and a stretching of the ectoderm covering the surface, are completely tuned to each other. Nevertheless this organizational process is not directed by the germ as a whole. Rather it is such that every one of the blastemas individually has autonomous migration tendencies. If one isolates a piece of ectoderm, then stretching takes place just as well as in ectoderm that is situated within a normal tissue relationship during gastrulation, and a fragment from the mesoderm of the upper lip of the primitive mouth spontaneously migrates from whatever arbitrary place on the germ inwards when transplanted.Of course these cells were already differentiated to do so. And this differentiation was clearly determined by the location of these cells within the developing germ when they were still situated in the latter (as above : A fragment from the mesoderm of the upper lip of the primitive mouth . . .). This location is a location on the pre-gastrula, which (location) is as such, however, little specific (as I assume). According to our theory it is made much more specific when it is at the same time a location in the organism's predesign, which is the macroscopically patterned organic lattice.
A condition for the migration of blastemas first of all is an autonomous change of shape of the individual cells. Cells that enter, via the primitive mouth (in amphibians), the blastocoel [cavity of blastula] (See Figure above ), lose their original cubic-prismatic shape. Instead they become elongated or bottle-shaped, in the course of which they remain still connected for some time with the surface of the germ by the 'neck of the bottle'.
For directly, for these form changes, responsible structures, the m i c r o t u b u l i and m i c r o f i l a m e n t s are eligible [...].
Just like in gastrulation, also n e u r u l a t i o n [...] is based on change of cell shape. The anterior part of the cell, that faces the emerging tube-shaped cavity, narrows, while the cell mass facing the periphery widens, and the whole cell elongates. In the case that this activity directly originates from the cells themselves, this must lead to the formation of the neural tube by folding of the neural plate (See Figure above ). Electronmicroscopic observations show that at the beginning of the transformation microfilaments and microtubuli are present below the inner cell wall that is in the process of narrowing, which orient themselves along the longitudinal direction of the stretching cell. Whether these structures only function as cytoskeleton or whether one should also attribute to them a dynamic function (for instance contraction of the filaments), is not yet a settled matter.[ We now can read in GOODWIN, B., How the Leopard Changed Its Spots, 1994, 2001, p.92 : " The cytoplasm is not just a blob of soft jelly with a lot of things dissolved in it. It actually has within it a complex and intricate structure in the form of a network of protein polymers [involved in microtubules and the like] that make up the cytoskeleton [...]. These polymers are constantly being made and broken down again, so the whole structure is extremely dynamic. It is, in fact, quite chaotic in its activities. The mechanical state of the cytoplasm -- its rigidity or softness -- depends largely on the state of the cytoskeletal polymers." ]
Anyway, an important datum is that after treatment with colchicine, a substance that fragments the microtubuli [...], the formation of a tube does not take place. The cells do not assume their typical shape anymore, and already elongated cells become subspherical again. Also microfilaments can reversibly be broken up into pieces, namely by means of cytochalasine [...] yielded from fungi, that, like colchicine inhibits the morphogenesis of cell connections [cell regiments] into tubular organ primordia.
Such findings only lift up a small tip of the veil that conceals the whole mechanism of gastrulation. They are supplemented by observations on sea-urchin eggs (See Figure above ). From the primary mesenchym cells -- as they initially lie above the inwardly migrating primitive gut, and which, by powerful pulsations, are freeing themselves from the endodermal tissue connection -- pseudopodia grow. These reach out into the blastocoel, and, by trial and error, make contact with certain parts of the inner side of the ectoderm. As the gastrulation proceeds, this 'wire contact' -- between (1) the anterior end of the primitive gut and (2) the place where the ectoderm invaginates ( This place lies asymmetrically with respect to the axis along which the invagination takes place) and [where subsequently] the oral aperture breaks through -- is significantly shortened. It is not demonstrated that at the same time the endoderm is also being 'towed along' by contraction of the pseudopodia. However, if one treats the developing germ with colchicine, a rotationally symmetric gastrula develops with an abnormal gut primordium, abnormal in the sense that the developing gut runs in the direction of the animal-vegetal axis. From this, one could conclude that the microtubuli fulfil a morphogenetic function (HADORN & WEHNER, p.154/5).
In the next document we continue to investigate the compatibility and necessity of organic-lattice structure in morphogenesis.
e-mail :
To continue click HERE for further study of the Theory of Layers, Part XXIX Sequel-12.