
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) Following ROHDENDORF, 1964, pp. 36, we discuss the infraorder Blephariceromorpha. We do this wholly in terms of the family Blephariceridae which is the only family known in this infraorder. This is the last infraorder of the Nematocera to be dealt with, that is, when we're done with it we have discussed all the infraorders of Nematocera as recognized by ROHDENDORF in his 1964 book.
Extent, evolution, and system
About 161 species of the single family Blephariceridae are known, which family subdivides into 5 subfamilies :
Edwardsininae with 1 genus (about 20 species),
Blepharicerinae (8 genera and more than 70 species),
Paltostomatinae (6 genera and more than 40 species),
Hapalotrichinae (3 genera and about 10 species),
and, finally, the Apistomyiinae (3 genera and about 20 species).
No doubt, the given numbers only have relative significance [that is, they only show the relative weights of the different subfamilies]. In fact there will be not less than twice this [total] number of species : This is evident by the fact that the majority of species have a very narrow geographical distribution, species that are associated with mountain streams. The Blephariceridae are distributed all over the world, reaching their greatest diversity in the Oriental and Neotropic regions. The individual subfamilies are characterized by a specific geographic distribution :
The Edwardsininae populate the Australian and the south of the Neotropical regions,
the Blepharicerinae live in the Holarctic and Oriental regions,
the Paltostomatinae live in the Neotropic, the Ethiopian (southern Africa), the Australian, and partly in the Oriental regions,
the Hapalotrichinae mainly live in the Palaearctic region (one species is known from Japan),
and, finally, the Apistomyiinae mainly live in the Oriental regions (one species is known from Australia, and one from southern Europe).
Up to date there are not known any paleontological documents of this group, therefore we must base our expositions of their history solely on the study of recent forms.
These peculiar insects are most closely related to the Tipulomorpha and Deuterophlebiomorpha : Out of the recent groups the Chironomidea as a whole are nearest to them. Examining the relationships of the Blephariceridae more precisely, one should recognize their deap-seated differences from all other recent Diptera. Similarity with them is in essence expressed by various purely convergent features, determined by the aquatic way of life of the larvae. The isolation of the Blephariceromorpha from the other diptera, the presence of certain ancient features (free large upper jaws [mandibles] in the winged insects, legs of the thin type, homonomous joints of the antennae, multisegmented abdomen) forces us to suppose the great age of the separation of this group from the common ancestors with the Tipulomorpha, which [separation] took place, probably, not later than the period of the lower Jurassic.
Chief features
The way of life and the [individual] development of these peculiar diptera let them be close to the strongly expressed relict group Deuterophlebiomorpha (dealt with in the previous document). The larvae of these insects [Blephariceridae and Deuterophlebiidae] live (sometimes communally ! [Presumably ROHDENDORF here means that these blepharocerid larvae can be found together with deuterophlebiid larvae in the same locality] ) in the fast-flowing water of mountain streams, crawling about on, and firmly attaching to, underwater rocks. The identity [or at least similarity] of conditions of a swift mountain stream, in which live the developmental phases of these two groups which are not genealogically connected but fairly far apart organizationally [that is, their morphological structure], determined the development of many similarities in the structure of the larvae and pupae, mainly of a protective nature. See next Figure.
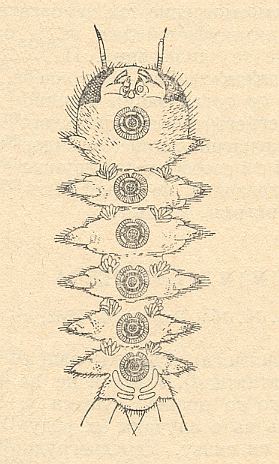
Figure 1 : Larva of Blepharicera fasciata WESTWOOD, ventral view.
Compare with the larva of Deuterophlebiidae Figure 1 of previous document .
(After LINDNER, 1930, from ROHDENDORF, 1964)
Therefore, comparing the Blephariceridae with the Deuterophlebiidae is very interesting. It helps to understand the essence of the conflicts which determine the tendencies in the historical development of these two groups of diptera.
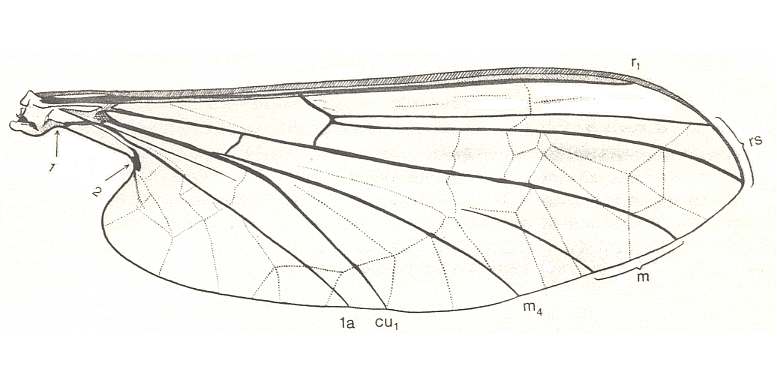
Figure 2 : Wing of Liponeura bilobata LOEW ( Blephariceridae). For better quality of this picture click twice on the image.
(After HENNIG, 1968)
The thorax is moderately enlarged, convex, bearing witness to the clear predominance of dorso-ventral muscles, lifting the wings. The head is large, with antennae of moderate length, very large eyes which often are complexly built, and with mouthparts that are structurally totally primitive (free teeth-bearing upper jaws!). Sometimes in certain groups (Edwardsininae and others) the mouthparts show reduction. Apparently in these diptera [that is, the Blephariceridae] the development of aphagia takes place only in certain groups, where it is a totally new acquisition. Many forms are still active predators, catching small insects, and visiting flowers. The structure of the central nerve-system, of the gut, and of the excretory and sexual systems, is, like the features of reproduction, unknown. The protective features, so clearly expressed in the larvae and pupae, are not clear in the adult : they are paleopterous [and thus do not protect their abdomen with their wings when at rest]. The legs are of a clearly expressed thin type.
Conflicts and determining tendencies in the historical development.
The process of acquisition of protective features of the larvae living on the surface of the bottom of water basins and feeding on aquatic plant-overgrowths [that is, browsing on algae growing on the underwater rocks and bottom surface of pools] was undoubtedly the chief conflict in the history of the blepharicerids. The necessity to slowly move about on the surface of the substrate (gnaw off overgrowths!) determined the working-out of protective features (firm integument and armament) and at the same time forced these forms to settle in more deficient (as regards fauna and flora) conditions of the fast cold mountain stream which is less populated by other animals. The conflict nature of the tendencies -- feeding on overgrowths, together with deficient digestion and therefore the need to consume a large amount of food (to which the relatively structural simplicity of the mouthparts and the low mobility of the insect bear witness) on the one hand, and vulnerability to predators when living in the open and slowly moving about, on the other, is the cause that had determined the development of this group of diptera. At earlier times refinement of feeding was, apparently, of no great significance in the historical development of the blepharicerids. Only more or less recently in their history a conflict began to arise between the feeding conditions of the larva and that of the winged form, probably in connection with the transition of this group of insects into conditions of mountainous terrain at sites of rocky slopes with poorly represented flora and fauna. The common ancient entomophagia [feeding on other insects] and the acquired nectarophagia of many winged blepharicerids begins, in the phylogenetically youngest groups, to be replaced by aphagia during the process of desimaginization. The refinement of feeding of the larva [which would then be expected] is probable but not yet proven. Comparing the blepharicerids with the deuterophlebiids is very interesting (see Figure 1 ). The coalescence of head and thorax in the larvae into one single little mobile complex provided with a sucker sharply distinguishes the first from the second. Evaluating the totally different more active and selective ability of consuming food in the [larvae of the] deuterophlebiids as to be a great step forward as compared with the less active devouring of the substrate (algal overgrowth on underwater rocks) in the blepharicerids, is certainly correct. In comparing the winged forms of the representatives of the mentioned families we must note the little development of regressive processes in the blepharicerids which are [in their winged phase] active, relatively long-living, insects of which the majority still eats, and in contrast must note the regressive features of the winged phase of the deuterophlebiids which are desimaginized aphagous insects totally lacking mouthparts.
Thus, the working-out of protective features in the larvae, and their transition to conditions of fast-flowing water, were the determining tendencies in the historical development of the Blephariceromorpha. In contrast to the Deuterophlebiidae there was no deficiency of food and thus no conflict was developed in their history : As a rule, the larvae did not intensify their feeding, they remained consumers of algal overgrowths. The winged phase continued feeding and only later on in its history developed aphagia [in some groups]. Of both groups [Blephariceridae, Deuterophlebiidae], as also of certain Tipulomorpha (Psychodidea), it is characteristic that the larvae live on the surface of the substrate, and thus no worm-like shape of the body [so characteristic of dipterous larvae] was developed. The winged phase of the Blepharicerids had changed little, and in its development the guaranteeing of sexual activity and reproduction were the determining tendencies, to which the most peculiar eyes in both sexes and the features of the flight-apparatus bear witness.
Conclusions
The determining tendencies in the development of the Blephariceromorpha had consisted in the development of an extreme stenobiosis (withdrawl into conditions of fast-flowing water of mountain streams). Refinement of feeding basically did not play a decisive role in their history : The majority of the winged [Blepharicerid] insects had preserved the ancient nature of feeding. Also the need for improved flight was significant in connection with feeding and sexual activity, which [need for improved flight] turned out to be the determining tendency responsible for the recent condition of these insects having mechanically refined wings, legs of the thin type, and refined eyes.
This concludes our exposition of the infraorder Blephariceromorpha (based on RODHENDORF, 1964). And having now considered all groups of the nematocerous diptera, this also concludes our systematic treatment of these diptera, and before we continue with the rest of the order Diptera, that is, with the infraorders Asilomorpha (containing most of the Brachycera-orthorrapha), Myiomorpha (containing most of the Brachycera-cyclorrapha), and the remaining small infraorders closely related to these, we will first study the historical developments, i.e. the various transformations, of the wing-venation within the Nematocera, and consider, on the basis of this, the possibility of the polyphyletic origin of the order Diptera.
In accordance with all this, the next document will deal with the origin of the order Diptera.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XIV.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V
Back to Evolutionary Part VIII