
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) Having, in the previous documents, considered the superfamilies Tipulidea, Chironomidea, Dixidea, and Culicidea, of the Infraorder Tipulomorpha, we will now discuss the last three superfamilies of this infraorder, namely the Psychodidea, Orphnephilidea, Rhaetomyiidea, and Pachyneuridea.
As usual, we begin the enumeration of evolutionarily important features by following ROHDENDORF, 1964.
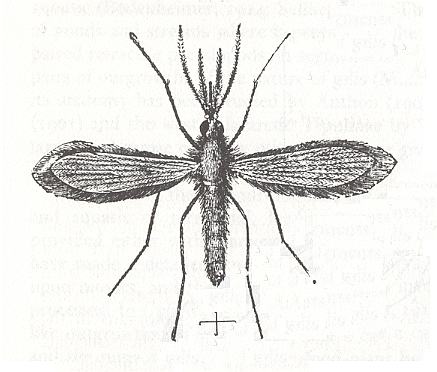
Figure 1 : Phlebotomus sp. (Phlebotomidae), female, Sudan, about x23.
(From RICHARDS & DAVIES, Imms' General Textbook of Entomology, 1977.)
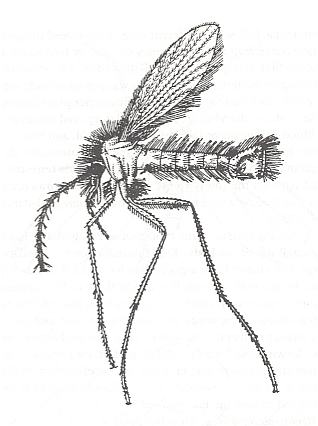
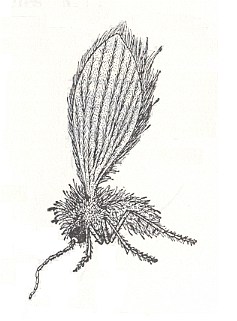
Figure 2 : Phlebotomus papatasii SCOPOLI (Phlebotomidae), male, general view.
(From ROHDENDORF, 1964, after PERFILJEV, 1937.)
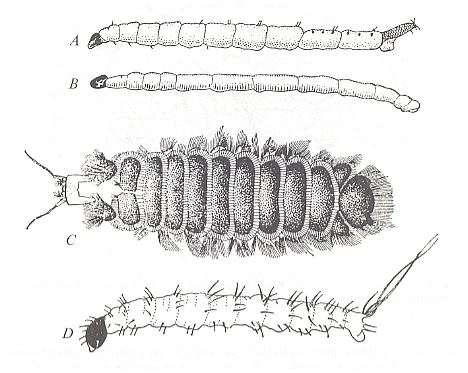
Figure 3 : Psychodidea, larvae
A -- Psychoda alternata SAY (Psychodidae), side view.
B -- Trichomyia urbica CURTIS (Psychodidae), side view.
C -- Sycorax silacea CURTIS (Psychodidae), top view.
D -- Phlebotomus papatasii SCOPOLI (Phlebotomidae), side view.
(From ROHDENDORF, 1964, after HENNIG, 1948-1952.)
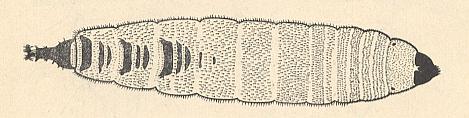
Figure 4 : Psychoda phalaenoides L. (Psychodidae).
(From KRIVOSJEINA, 1969, after SATCHELL, 1947)
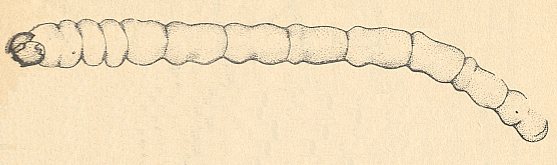
Figure 5 : Trichomyia sp. (Psychodidae).
(After KRIVOSJEINA, 1969)
Immediate historical documents [fossils] relating to this superfamily are insignificant, although more numerous than those of Culicidea. Reliable fossil remains of Psychodidea exclusively belong to the Tertiary fauna, while more ancient finds are uncertain (Mesopsychoda dasyptera Br., an insufficiently known form from the Lias [lower Jurassic] of Ust-Balei). All tertiary forms are found in Baltic amber and belong to three recent families : Psychodidae (not less than 30 species), Nemopalpidae (one species), and Phlebotomidae (one species). Apart from these forms of which the assignment to one or another definite family is without problem, some forms are known with uncertain systematic status. All these data are significantly outdated, and the given numbers only have relative significance. Thus, the discovery of species of the subfamilies Trichomyiinae and Sycoracinae is very important, which today are relict groups poor in species : The fact of them being found in the upper Eocene fauna clearly demonstrates their relict nature, pointing to their substantially greater abundance in the past.
The phylogenetic relationships of the Psychodidea is still little clarified. Their connections with the most ancient Tipulomorphs are evident, even perhaps with the Dictyodipteromorpha [found in the upper Triassic -- the age of the fossil site is perhaps a little bit younger, lower Jurassic], and with very primitive forms, of which the winged phase had not yet lost the complete set of mouthparts and having also not worked out perfectly costalized wings or ones that carry a phragm. An interesting fact is the development of a cover consisting of macrotrichia [macroscopic hairs] on the body and wings of the adult stage, by which they are close to the Culicidea. However, it is very probable that this feature is just a trait retained from very early ancestors, and so absolutely not bearing witness to some immediate kinship between these groups [Psychodidea, Culicidea].
The relationships between the groups within the superfamily Psychodidea are still little understood. While the mutual separateness and essentially different pathways of the historical development of the Psychodidae and Phlebotomidae are more or less clarified, things are totally different with respect to the third family, which has an evident relict nature and is only little studied. The insufficient expositions about the organization and history of the Psychodidea is particularly a pity, when one realizes the presence of very primitive features of the larvae, which set this superfamily totally isolated among the other Tipulomorphs. Such is the presence in the majority of the larval forms of rudimentary anterior spiracles which points to a high age of this group : The transition to the water, as was indicated earlier, was the primary solution of the conflict, which [solution] determined the formation of the first representatives of the series of Tipulomorpha. The retention of the anterior spiracles indicates the branching off of the Psychodidea from the most ancient Tipulomorphs, which only populated the surface layers of the water, more precisely, the boundary zones of water basins, that is, the wet soil of the banks.
The basic features of organization and of the essence of the conflicts in the historical development of the Psychodidea can at present be clarified only in the most general terms (see Figure 2 and Figure 3 ). Living, on the one hand, in water, or in variously polluted accumulations of wet materials and, on the other, in fast flowing water-basins, in short, the development of an aquatic way of life of the larvae (but the feeding habits of the larvae remain quite unknown!), together with the development in the adult form (imago) of running legs, of broad hairy wings of the ancient lifting type, and the reduction of the mouthparts, which can only take in liquid food -- these are the features of the family of moth-flies, Psychodidae. The basic conflict -- insufficient [amount of] food -- was, in the history of the moth-flies, as it seems, solved by the refinement of feeding of the larvae colonizing a humid environment and became consumers of certain abundant organic materials, probably vegetable organisms involved in processes of decay. The abundance of nutritive substances at the time of larval development made it possible for the winged phase to go over to vegetable food (and in several cases evidently also to aphagia).
Something different took place in the history of sand-flies, Phlebotomidae. Originally living in a humid habitat and the insufficiency of food had created a conflict, and that was solved by the development of the peculiar 'sapro-microphagia', that is, feeding on different decaying materials which were present in small quantities (corpses of insects, excrements of small animals), or perhaps development of bacteriophagia, and so on. Clearly, this solution of the conflict could not be the complete solution of it and did not stimulate the development of aphagia or phytophagia of the winged form, which remained predacious and possessed piercing mouthparts. Later the habit of sucking blood from vertebrates developed, and correlated with it, refinement of the flight abilities. All this turned out to be a substantial acquisition in the history of the sand-flies, which enabled this group of Diptera to become widely distributed insects. At the same time, and in the most direct connection with this process, a terrestrial, partly almost xerophilic (drought-loving), way of life of the larvae developed, which [larvae] had acquired the ability to live in the soil of burrows inhabited by vertebrates, and feeding on different decaying animal substances.
It is necessary to note yet another important feature in the history of the Psychodidea, which consists in the acquisition by these Diptera of small body size. Almost with complete certainty can we maintain the secondary [i.e. derivative] nature of the small sizes of the Psychodidea : to this the rich and complex venation of the wings bear witness, which [venation] became a typical rudiment of the supporting skeleton of the once large wings of their ancestors. See next Figure (consisting of the subfigures 34-37).
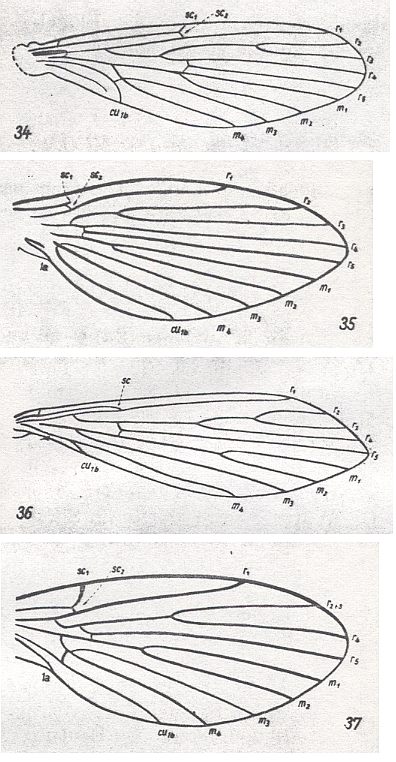
Figure 5a : Wings of recent Psychodidea.
34 -- Nemopalpus zelandiae ALEX.
35 -- Ulomyia fuliginosa MEIG.
36 -- Phlebotomus papatasii SCOP.
37 -- Trichomyia urbica CURT.
(From HENNIG 1954, after ALEXANDER, 1927)
To insects of such small sizes, as the Psychodidea, which, in addition to it, possess a rich cover of macrotrichia, such a complex wing-venation is functionally inconsistent. It is a clear remnant of ancestral features. The little biological significance of flight in the well-running moth-flies, the broadening of their wings, and the transformation of them into peculiar partly cover organs [i.e. the wings folding back over the abdomen, and in this way covering it] confirms the expressed position about the reduction of the absolute dimensions of the body in the historical development of this group of Diptera. This reduction in body size turned out to be one of the solutions of the conflicts in the development of the Psychodidea, which enabled them to colonize every kind of 'micro-station', develop microphagia, and thus this reduction became a very useful acquisition.
In the following we add some data concerning evolutionarily important ecological features of the Psychodidea obtained from OLDROYD, 1964 :
The Psychodidea are not so heavily committed to the water as may other Nematocera. The main body of the Psychodidea consists of an aquatic half -- the family Psychodidae -- and a half that is apparently terrestrial -- the family Phlebotomidae.
Let us first consider the aquatic half, the Psychodidae.
They are the moth-flies or owl-midges, commonly to be seen on the inside of the windows of houses, and abundant at any place where organic matter is present. The adult flies have pointed, leaf-shaped wings, covered with hairs, and so look rather like small moths, especially as they often hold the wings together like the roof of a house, as many moths do. These moth-flies have aquatic larvae, which live mostly in puddly, shallow, semi-fluid conditions, with a high organic content and often deficient in oxygen. The larva has two pairs of spiracles, those on the thorax raised up on short pegs, and the posterior pair at the tip of a rigid siphon, which ends in four lobes with water-repellent hairs. Such larvae mostly crawl about on or near the surface of the mud, and some of them have spines, or feathered processes along the body, which help in this environment. Others live rather on the edges of mud, in the bottoms of ditches, and in crevices near by. It is characteristic of Psychodid larvae, a primitive family, that they are really only semi-aquatic, and only a few of them venture into deep water. The genera Ulomyia, Maruina and and the species Pericoma californica live in torrents and rapids, with special sucker-like organs with which they cling to the rocks. Even these Psychodid larvae are often not below the water-line, but hang on where they are constantly soaked in spray.
Psychodidean flies belonging to the family Phlebotomidae are the true 'sand-flies', though this name is often used, especially in conversation, for the biting midges of the family Ceratopogonidae.
Phlebotomid sand-flies are found in hot countries, that is, throughout the tropics of the world, and in sub-tropical and warm-temperate regions, as far north as Manchuria and Paris, and southwards to the Cape of Good Hope, into the northern part of Australia and over all South America except the cold southern tip. They have not as yet penetrated to the Hawaiian islands. About 350 species are known, of which only the adult females suck blood.
Sand-flies are tiny, fragile flies with their wings more elongate but less hairy than those of the moth-flies and with a longer proboscis. Their larvae are hidden away in crevices in the soil, sometimes 20-30 cm down, in cracks of masonry, between stones, and so on. It is a paradox of this group that the adult sand-flies, as this name suggests, are associated principally with hot and arid places, but the larvae in fact are dependent on moisture to a degree that is unusual in an apparently terrestrial animal. However humid the air, the sand-fly larvae and pupae die very quickly unless liquid water is present, and on contact with their skin.
Sand-flies are now known to be commoner in tropical forest than had previously been realized. Many hide in crevices of the great buttress roots of the high forest trees.
We have seen that the Psychodidae, the moth-flies, are all aquatic as larvae. Apparently the larvae and pupae of Phlebotomus (a genus of sand-flies) are also really aquatic, but they have colonized a microclimate that exists in crevices, where condensation of atmospheric moisture provides the droplets of liquid water that they require. Even in the hottest climate this water persists. The food of the larvae is organic : decaying vegetable matter, dead insects, and particularly the dung of small animals. The perhaps limited food in the microclimate of the larval crevice is in this family supplemented by the adult's sucking the blood of some involuntary host.
The legless larvae of sand-flies are distinguished by having long, isolated hairs, with a few especially long ones posteriorly.
All the representatives of the superfamily Psychodidea have wings that belong to the
Primitive lifting (psychodoid) wing type.
Description (taken from ROHDENDORF, 1951, with changes) of the type as such :
Representatives of the type
Wings of the primitive lifting (psychodoid) type are characteristic of species of two families of the superfamily Psychodidea, namely the moth-flies, Psychodidae, and the sand-flies, Phlebotomidae. The Nemopalpidae, also belonging to the superfamily Psychodidea, is a relict group, and the wings of its representatives probably also belong to this type.
Here we give some Figures (a number of them already given earlier) depicting wings of this type.
Wings of recent Psychodidea, belonging to the primitive lifting type.
34 -- Nemopalpus zelandiae ALEX.
35 -- Ulomyia fuliginosa MEIG.
36 -- Phlebotomus papatasii SCOP.
37 -- Trichomyia urbica CURT.
(From HENNIG 1954, after ALEXANDER, 1927)
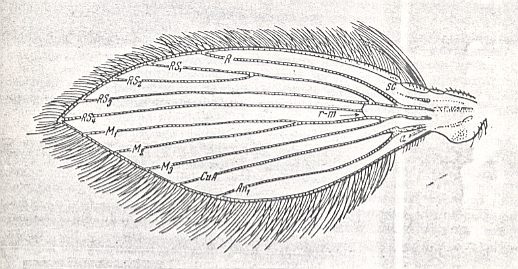
Wing of the primitive lifting type.
Psychoda sp. (Psychodidae).
(From ROHDENDORF, 1951, after HENDEL, with changes)
Size of the wings
Shape of the wings
The wings of this type are of 'middle' length or are elongated. Their width is 2.5 - 4 times shorter than their length. Sometimes the wing is even more than 5 times narrower than long. A sharp narrowing of the wings is observed in the most minute forms (some species of the genus Phlebotomus, for example Ph. dentatus SIN., Ph. minutus ROND.). The relative large Psychodidae possess very broad wings (the genus Pericoma, [ROHDENDORF refers to Figure 24 in his work (see Figure above ), but the depicted wing is, according to his subscript, belonging to the genus Psychoda] ).
Fore- and hind-margin of the wing are convex, more or less similar to each other. The apex of the wing is sharply pointed (the angle is a little smaller than a right angle). The apex is not clearly distinguished [from the rest of the wing]. The basiala is fairly sharply distinguished from the wing-blade and has a peculiar structure. The anal lobe is expressed by the general convexity of the wing's hind margin. The alula and the thoracic scale are absent. The wing-scale is present in the form of a germ ('a beginning') on the basiala.
Skeleton of the wing
The wing-venation consists of many straight longitudinal veins, fan-like distributed over the wing-blade. The branchings of the radial and medial systems do not lie at a single transverse level in the wing, but positioned in the wing-blade at different places. The venational pattern does not show any sign of costalization. In this respect this wing type sharply differs from all other wings. The costal vein runs all along the wing. The subcostal vein is almost totally reduced. The radial (5 branches) and medial (3-4 branches) are of equal thickness, lying at equal distances from each other, uniformly and insignificantly diverging. Almost all radial veins end up at the wing's fore-margin, while the last radial (fifth) branch ends up at the sharp wing-tip (i.e. at its apical corner). Cross-veins are present only in the form of the short radio-medial (rm) and medio-cubital cross-vein (mcu) being placed near the wing-base [The mcu (and also the rm) is well visible in 37 ( Trichomyia ) of the Figure above ]. The basiala is well developed mainly in Psychodidae. In the Phlebotomidae it is reduced. The venation of the basiala is peculiar. This part of the wing itself consists of two parts : a broad proximal one, and a narrowed distal one. The proximal part, which is the part of the basiala closest to the thorax, carries a germ of the wing-scale, and includes the distinguished basal parts of the costal, radial, and anal veins. The narrow distal part of the basiala includes a large number of individual elements, which consist of (1) the lower part [Russian : nyz] of the cut-off piece of the costal vein (see Figure above ), (2) the vestigial subcostal vein, (3) the basal part of the radial vein, and (4) the two basal parts of, respectively, the cubital and anal vein [ in the Figure just referred to, we can see that, in addition to these four mentioned elements, the distal part of the basiala contains the common trunk of the medial system]. The general structure of the basiala of the psychodoid wing can be characterized as being a peculiar elastic joint which makes it possible for the wing to swing, letting, during the wing-beat, the basiala stand still. The wing-membrane is not especially investigated.
Covering of the wing
In addition to being covered by short delicate hairs (microtrichia), the wings of this type are covered by long and flattened spinelets, or better, scales, distributed along almost all veins of the wing. Especially long spinelets-scales sit on the margin of the wing, where the spinelets of the hind-margin are markedly longer than those of the fore-margin (Here the germ of costalization of the wings only expresses itself by this one feature ! ). A dense covering of spinelets-scales is present on almost the whole wing, and is especially dense in the representatives of the family Psychodidae. Sensoria are not especially investigated and are visible in the basiala only.
Functional characteristic
The representatives of the primitive lifting (psychodoid) type have a very weak flight, in which the basic effect is the overcoming of the forces of gravity, that is, generating lifting power. A more active flight is worked out in the sand-flies (Phlebotomidae), undoubtedly connected with the development of blood-sucking by the females. Minute sizes determined the character of flight in the sand-flies which flight comes close to that of those insects whose wings belong to the type named "feather-wingedness" (ROHDENDORF, 1949), and [which flight] is characterized by its slowness, by extremely low load, and by the ease of the flying insects to be carried by even weak air currents. Accurate data about the character of flight of moth-flies and sandflies are absent. There are indications that sandflies do not fly about farther than 150 m from the place of emergence of the adult, but it seems to me that these observations do not exactly anwer our question. It is very likely that we here have to do with the flying insect passively being carried away over different distances, which is not dependent on the quality of its flight. In all, the flight of insects with psychodoid wings does not have equal significance. In some cases (Psychodidae) flight is connected primarily to the sexual function and has little significance for collecting food. In other cases, in sand-flies (Phlebotomidae), flight is a necessary aspect of their life-activity, guaranteeing blood-sucking.
History of the type and its transformations
Reliable fossil material only exists in Baltic amber (Tertiary, upper Eocene) and belongs to genera of the family Psychodidae. Apart from this there exists an indication from the literature about the discovery of an alleged species of Psychodidae from lower jurassic deposits of the Angara region [the Angara is a river] (namely from the Ust-balei finding-place [ Irkutsk ] : Mesopsychoda dasyptera B., R. etc). However, the incompleteness of the description and the totally imperfect drawing of the fossil do not allow judgement as to the correctness of the identification which can be assessed as being doubtful. Investigation of recent wings of the primitive lifting (psychodoid) type outlines the evolutionary pathways of this type. The source of its formation undoubtedly were forms of the tipuloid type [ See in Part II : the last of the sections dealing with the superfamily Tipulidea (last fourth of document) ], namely its first forms which had not yet developed costalized wings. The formation of the type of psychodoid wings went its way on the basis of the general direction of the evolution of the superfamily Psychodidea, which had distinguished itself first of all by the decrease of body size. The recent state of the psychodoid type indicates a clear subdivision of it into two separate subtypes. In one group [subtype], the sandflies (Phlebotomidae), the wings did not undergo a significant increase in size. Apparently, their wing-size did not increase as compared with those of their ancestral, ancient, tipuloids. The wings merely became pointed [at the apex], acquired a dense covering of spinelets, and lost the ability to fold the wings back over the abdomen [when the insect is not flying], that is, paleopteria was developed. The absolute size of the insects decreased strongly, and the ability to suck blood appeared. These features turned out to be the chief determining factor in the formation of the present subtype. Another evolutionary pathway was realized in the moth-flies (Psychodidae), which had their [relative] wing-size significantly increased. Their wings became peculiar broad, pointed blades densely covered by spinelets-scales. The moth-fly began to hold [when resting] its huge wings like the roof of a house, that is, fold them back along the abdomen, having it covered by them, as do many moths, cicadas, and other insects. The wings of moth-flies certainly possess a very large relative surface area and a low load per unit surface area, that is, features characteristic of broad-wingedness, a type of flight-apparatus of insects which is most abundantly repesented in the insect order Lepidoptera [butterflies and moths]. The evolution leading to such wings went in the direction of an increase of lifting power of them by decreasing their load, realized by the increase of wing-size, without a corresponding strengthening of the muscular apparatus.
Apart from connections with the tipuloid type, the psychodoid wings are more or less similar to those of the culicoid type [See Part III , where we deal with the superfamily Culicidea (last fifth of document) ]. This similarity consists in the fact that the wings of both types are densely covered by spinelets or scales. However, this similarity is limited by the following feature : the character of the wing-beat. The very structure of the wings points to a totally different pathway of specialization in the blood-sucking mosquitos (Culicidae).
This concludes our exposition of the superfamily Psychodidea.
The superfamily Orphnephilidea (also known under the name Thaumaleidea) including about 70 species of four genera of the single family Orphnephilidae is not sufficiently known by me (that is, to ROHDENDORF). The data given in the existing literature concerning these peculiar Diptera wonderfully illustrates an example of 'non-correspondence' of the organizational features of the larval and adult phases, having found expression in the very different assessment of the systematic position of these Diptera, which on one occasion were considered to be a subfamily of the Chironomidae, on another as a separate family, and, finally, on yet another occasion as an individual superfamily, which shows almost certainly connections with the Brachycera [true flies]. The Orphnephilidae, totally unknown as fossils, are distributed on almost all continents of the Earth and are clearly mountain-dwellers.
We continue with some data about this group taken from OLDROYD, 1964.
Thaumaleidae, with the alternative, almost unpronounceable name of Orphnephilidae, are tiny midges about 3 mm long, with their eyes close together in both sexes, and not, as is usual, in the males only. Although about fifty species exist they are enigmatic insects, remote and little known. They occur in high latitudes and in mountainous areas : in Europe chiefly in Lapland, Norway, and Scotland, and in the mountains down to Italy. They also occur in the United States and Canada, and in New Zealand and Tasmania.
At one time Thaumaleidae were thought to be land-midges akin to the Mycetophilidae ( = Fungivoridae, fungus-gnats), but after almost a century the larvae were discovered in cold mountain streams where a shallow layer of water runs over rocks. So shallow a layer, in fact, that the larva is not completely submerged, but creeps about in a mere film of water, browsing on vegetable detritus on the stones. According to Vaillant the larva normally touches the rock only at its two ends, and can glide quickly away on the surface film when disturbed. The larva is superficially like those of the biting midges of the family Ceratopogonidae, with a well-formed head, a thoracic proleg, and hooks which serve as a proleg posteriorly. The pupa is found near by in mud or wet moss.
The correct position of this family among the Nematocera is a matter for argument among systematists, who reach different conclusions by considering adult and larval characters. Biologically this family is interesting as an experiment towards aquatic life, one which has probably been made a number of times in the evolution of flies. The larvae are amphipneustic, that is, they have open spiracles on the thorax and at the tip of the abdomen. In this, in having abdominal plates, and in their habitat, they are remarkably like some Psychodids, particularly those living in not fast-flowing water where thin trickles of water flow over stones. Their present distribution, broadly interrupted in all the warmer areas of the globe, suggests an ancient group in retreat, clinging to its specialized habitat. So they are best considered as an evolutionary experiment that did not succeed.
The superfamily Rhaetomyiidea until recently [1964] was only known from the upper Triassic fauna of central Asia. However, in 1962 a peculiar recent form from Australia, namely the genus Perissomma COLLESS, represented by two species from the south of that continent, was described. The genus is considered to belong to the sharply isolated family Perissommatidae which family almost certainly is a relict of this ancient superfamily, which [superfamily] had reached a highly specialized state in Mesozoic times.
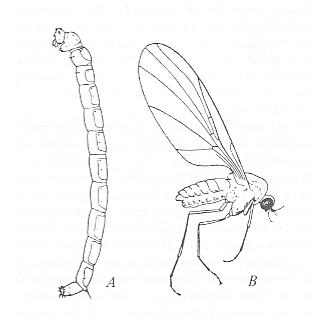
The Superfamily Pachyneuridea consists of a single family, the Pachyneuridae, represented by 5 species. The flies of this superfamily probably look more or less like fungus-gnats. We have no drawings or photographs depicting the general view of the adult form, but we have one depicting the larvae :

The Pachyneuridae were assigned by ROHDENDORF, 1946, initially to the 'Nematocera oligoneura', which group of Diptera (consisting of fungus-gnats, gall-midges, and the like) is more or less equivalent to the infraorder Bibionomorpha as (later) established by ROHDENDORF, 1964. In the latter work, however, he assigned them to the infraorder Tipulomorpha, without giving arguments for the legitimacy of this transition. Apart from explicitly mentioning the Pachyneuridae as representing a superfamily of the Tipulomorpha, he does not separately deal with them while he does so with the other superfamilies of that infraorder.
Continuing with data on Pachyneuridea :
We now give the additional (few) data we can get at the moment about the Pachyneuridea from two sources, namely again from ROHDENDORF, 1946, Evoljutsia krila i filogenes dlinnoysich dwykrilich (Oligoneura [ Diptera, Nematocera], (The evolution of the wing and the phylogeny of Oligoneura (Diptera, Nematocera)), and from KRIVOSJEINA, 1969, Ontogenez i evoljutsia dwykrilich nasjekomich, 1969 (Individual development and evolution of the two-winged insects)
Let us then give these data :
The known 3 or 4 species of the family Pachyneuridae, are perhaps most interesting with respect to structural features of the wings which, in this family, are the least specialized as compared to the wings in the other Oligoneura : In the wing any soft and elastic membrane at the wing-base (basiala) is absent, which wing-base does not possess adaptations to enable 'propeller-flight' [that is, to enable the wing to 'imitate' the movement -- and especially the effect -- of a rotating propeller (airscrew) ]. The 'handles' [i.e. the basal sections] of almost all wing-veins are well developed. They are not reduced. The specialization of the wing only consists in the reduction of the basal section of the median vein [ See next Figures].
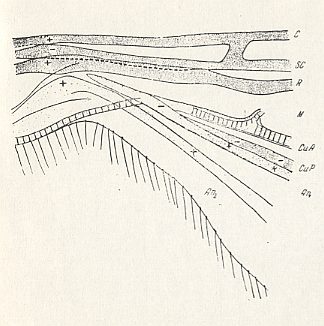
Figure 9 : Pachyneura fasciata ZETT. (Pachyneuridae).
Wing-base, basiala, diagrammatic.
From top to bottom, (bases of veins) : C = Costa, SC = subcosta, R = Radius, M = media, CuA = anterior cubitus, CuP = posterior cubitus, An1 = first anal vein, and to the left : An2 = second anal vein.
A + sign means that the vein lies on a convex ridge, while a - sign means that the vein runs along a concavity.
See for wing-venation Figure 1 in Part II .
(After ROHDENDORF, 1946)
We can compare this drawing of Rohdendorf of the wing-base of Pachyneura with a diagram -- also drawn by ROHDENDORF, 1946, -- in which the wing-base of Phryne ( Phryneidae [ = Rhyphidae] ) is depicted. The Phryneidae belong to the infraorder Bibionomorpha :
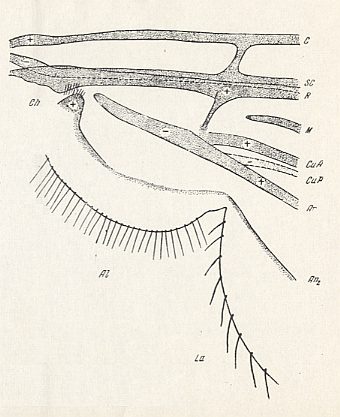
Figure 10 : Phryne sp. ( Phryneidae).
Wing-base, basiala, diagrammatic. Ch = chaetarium. For the meaning of the other symbols, see previous Figure. ( Below CuP lies the vein An1 .)
(After ROHDENDORF, 1946)
The next Figure depicts a wing of one of the representatives of the family Pachyneuridae :
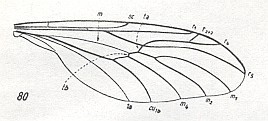
Figure 11 : Wing of Pachyneura fasciata ZETT.
(From HENNIG, 1954, after DUDA, 1930.)
The family Pachyneuridae is known from only a few localities in Europe, the Far East, and North America. Metamorphosis and biology of these insects remain unknown.
In pieces of damp dark wood or in tree-stumps live the larvae of Pachyneura sp. (Pachyneuridae). See Figure 8 . In some cases a great many larvae of Pachyneura sp. are found in woodblocks.
( KRIVOSJEINA, 1969, p. 22).
For the time being this is all we can say about the superfamily Pachineuridea. It is clearly a phylogenetic relict. Its description forms, so we could say, the transition to that of the next Infraorder of Diptera, the Bibionomorpha. See next document.
Will be supplemented if necessary . . .
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part V.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V