
Part IV : Sequel to the special theory of the determinative structure (Nomos) of the material world : Determination of stable patterns
Sequel to the theory of the true and ultimate determinants (if-then constants) of the material world, together constituting causal chains, causal webs, and the determinative connections in low-level and high-level material processes, again with respect to the formation of stable, enduring, repeatable patterns in the material world (space-time world, Explicate Order) : Chreodes and organisms.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) Chreode landscapes in the Implicate Order and the formation of organisms.
Introduction.
After having -- in the previous document -- discussed the generation of branched snow-crystals in terms of chreodes and If-Then constants -- from which snow crystals we can generalize our findings to all branched crystals -- it is natural now to present a discussion of the generation of organisms also in terms of chreodes and If-Then constants.
However, in contrast to the generation of crystals, the generation of organisms is utterly complex, and -- what is worse -- only partly understood. Especially the aspect of, I would say, sophisticated global organization, as it seems to take place in the individual organic development of every multicellular organism, is, I might say, not understood at all. And such a global organization or orchestration of the processes participating in individual development in which the organism-as-a-whole seems to be one of the causal factors or maybe even the sole causal factor leading to its generation, cannot simply be just an effect directly flowing from the participating materials at hand. It must have itself been (gradually) developed historically, that is, orchestrated individual development as such must itself be the product of a long evolution taking perhaps millions of years. We will concentrate on this product -- organized individual organic development -- as it is now, and see how it can be described generally in terms of chreodes and If-Then constants. But because of the enormous complexity, diversity, and incomprehensibility of organic individual development -- morphogenesis -- we cannot give a definitive account. Our 'exposition' will necessarily be incomplete, general, and speculative. And like our exposition of branched crystals (previous document) its significance is philosophical rather than practical.
Preliminary exposition of organic morphogenesis in terms of chreodes and If-Then constants.
While a crystal of a given chemical substance develops as soon as certain more or less external physical conditions ( [1] the substance's local concentration being within a certain range, [2] presence of large enough crystal seeds or other appropriate substrates upon which crystallization can start, and [3] local temperature and pressure being within a certain range ) happen to prevail, and which conditions can prevail relatively easily and repeatedly, the generation of an (individual) organism needs a very specific and 'fully worked out' detailed initial condition which includes not only physical conditions like temperature and pressure but also detailed chemical, spatial, and biological conditions. The overall necessary initial condition for an organism to be generated somewhere is, consequently, very narrow, that is, very specific, and therefore not likely to prevail easily here or there without any previous specific 'elaboration' leading precisely to it, like it does so prevail in the case of crystallization. Organisms will not come into being in the way, say, snow crystals do.
For an organism to come into being at some location (on the Earth's surface) information must materially be present at that location, that is, for every species of organism the appropriate specific information must be present at the site where an individual of such a species is to be generated. In the case of the generation of a crystal such information must, it is true, also be present, here in the form of a fragment of the proper crystal lattice or at least in the form of a substrate which contains some of such information, but this needed information is relatively simple and consequently appears easily on the scene as soon as the general physical conditions are within a certain range. In the case of the generation of an organism, on the other hand, the necessary information that must be present is too complex and too 'worked out' to (be able to) just spontaneously appear at some given location simply out of mere general physical conditions prevailing there. There must already be something there (that is, materially be there [coming from, or brought there by, another organism] ), containing or embodying this information, something that is not simply generated out of here-and-now prevailing not very specific physical conditions. And only when this information is actually present it can generate the necessary initial condition of a dynamical system that is going to produce an organism. And moreover, still other, also specific, conditions, namely a specific chemical and structural machinery, must prevail in order that the information contained in that what is materially present can actually be read, that is, can actually be applied. And only then the mentioned necessary initial condition will be generated. All this is, in the case of multicellular organisms, realized by the presence of a fertilized egg-cell. The information we spoke about is then contained in the DNA of that egg-cell. And this egg-cell also contains the mentioned chemical and structural machinery that can read and apply that information. When this is done the necessary initial condition of the dynamical system that is going to produce the organism is indeed created. And because DNA only codes for either RNA (which serves to transport information from the cell nucleus to certain sites in the cell's cytoplasm) or -- via RNA -- specific amino acid sequences (and thus for specific proteins), DNA does not contain information that says how to actually construct specific supramolecular structures (organisms or macroscopic parts of them). It only supplies the proper materials at the proper places and at the right points in time. In fact DNA does not even code for most of such materials but mainly for the enzymes that catalyze specific biochemical reactions. And these reactions in turn create the mentioned materials, that is, materials with their own molecular shapes and structures. What has thus so far been generated is not an organism or one or more of its macroscopical parts but only the necessary initial condition of a highly specific dynamical system. And it is this dynamical system that will, after having passed through a large number of stages or states, finally generate the organism or some specific macroscopical body part or feature of it.
At several stages of individual development certain dynamical subsystems will be activated. These subsystems generate the mentioned specific parts of the developing organism. Also these subsystems need specific initial conditions for them to get started. These conditions are also the result of the application of information contained in the DNA. But while in the fertilized egg-cell all information contained in the DNA is available, that is, can be set free (for RNA to be synthesized and therefore information to be copied) in order to get read and applied (through the transported segment of RNA), most subsequent cells (generated by the repeated division of the egg-cell) are already specialized which here means that only a specific part of the information contained in their DNA -- we say a part of the information contained in the genome -- can be read and applied. So after repeated division of the initial fertilized egg-cell we get several groups of such genetically specialized cells. And all the cells in such a group are together going to constitute (through the application of the readable part of their genome) the one initial condition of a dynamical subsystem that will eventually generate a specific macroscopical body part or feature of the developing organism. Precisely what part of the genome, in what cells, at what time, and for how long, becomes (exclusively) readable, is, as we assume, determined by the states of the overall dynamical system that started off from the fertilized egg-cell.
In order to link up with our exposition (had in previous document) of the generation of branches in crystals we will, at least to begin with, concentrate not on the generation of the individual organism as a whole, but on the generation of its macroscopic morphological parts (like, for instance, the leaves or flowers of a plant, the wings or legs of an insect, the kidneys or liver of a vertebrate animal, etc.). And, as has been said, the overall dynamical system determines what parts of the genome of what cells become available. So when the overall morphogenesis of the individual organism is well under way special initial conditions for the subsystems mentioned will be created at the proper places within the organism's developing body and in the proper order. And the subsystems will then generate the macroscopic parts of the developing organism.
In terms of chreodes and If-Then constants all this, i.e. the generation of a macroscopic body part, can be described as follows :
At a certain stage of the overall dynamical system a group of cells becomes specialized which means that only a definite part of the genome -- i.e. a definite part of the DNA -- of these cells can be read and applied, which leads to the creation of a special initial condition or first state of a particular dynamical subsystem. This first state consists of a number of simultaneous subconditions. Each such a subcondition, itself residing in the Explicate Order, triggers -- by being injected -- the activation and application of some If-Then constant in the Implicate Order. The application of that If-Then constant results in the content of its Then term to be projected into the Explicate Order. And this projected content is itself a subcondition of the next state of the dynamical subsystem, and in turn triggers the activation and application of another If-Then constant, and so on, eventually resulting in a sequence of consecutive activations of If-Then constants resulting in a consecutive sequence of projected contents. In this way each other subcondition of the initial condition of the subsystem leads to its own sequence of consecutive activations of If-Then constants resulting in a consecutive sequence of projected contents. All this together constitutes the succession of states of the dynamical subsystem which will generate the corresponding macroscopical part of the developing organism.
Described in terms of chreodes we can say that the mentioned initial condition of the dynamical subsystem is so accurately generated and so protected against possible perturbations that it is either precisely the entry of a one-entry chreode (see Figure 14 of the previous document ) or one of the entries of a many-entry chreode (see Figure 15 of the previous document ). We can assume that in the long biological history of the planet the generation of the corresponding macroscopical body part of the individual past organisms of the same species has taken place so often accordingly (that is, in virtue of the large number of these individual past organisms) that indeed in the Implicate Order a corresponding chreode has been 'carved out', and thus that the dynamical subsystem -- in the Explicate Order -- generating such a macroscopical body part is stable. We have concluded this on the basis of the observation that corresponding body parts in individuals of the same biological species (that is, if we go from one individual to the next) are in a marked degree morphologicaly similar to each other, if not, morphologically identical. Think for instance of the morphology of the leaves in different individuals of the same species of tree (and indeed of the leaves of one and the same individual tree), or of the morphology of the flowers in different individuals of the same species of plant (and indeed of the flowers of one and the same individual plant). The same goes for corresponding skeletal parts in individuals of the same species of vertebrate, or the wing venation in individuals of the same species of insect.
In all this we must hold on to the rules that are in force towards the Implicate and Explicate Orders and the If-Then constants as the elementary determinative units :
Prolonged working out of the structure of the organic chreode.
The above elaboration as to the presumable structure of the organic chreode is far from complete. Think about the phenomena of regulation and regeneration which have been found to be inseparably connected with organic morphogenesis. While regeneration means the restoration of the final product or end state of morphogenesis after that final product has been damaged, regulation guarantees the generation of the final end state even after there has taken place a severe perturbation or damage somewhere along the dynamical system's trajectory long before the end state is reached by that trajectory. Especially, every case of regulation implies that the overall dynamical system generating the organism partly follows a different dynamical course, different, that is, from its 'normal' course, but nevertheless eventually leading to the same end state that would have been reached without the perturbation event having happened. But also many cases of regeneration bring with them alternative dynamical courses, as when a fragment of a flatworm (obtained by cutting out a longer or shorter segment of the full grown body of a living worm) again grows up until the state of the full grown body has been reached.
The initial conditions (themselves a result of damage or perturbation) of the alternative dynamical routes, and these routes themselves, have not been accounted for by the two images of organic chreodes discussed (Figure 14 and 15) in the previous document. These images are (with some text) reproduced here as respectively Figure 1 and Figure 2 :
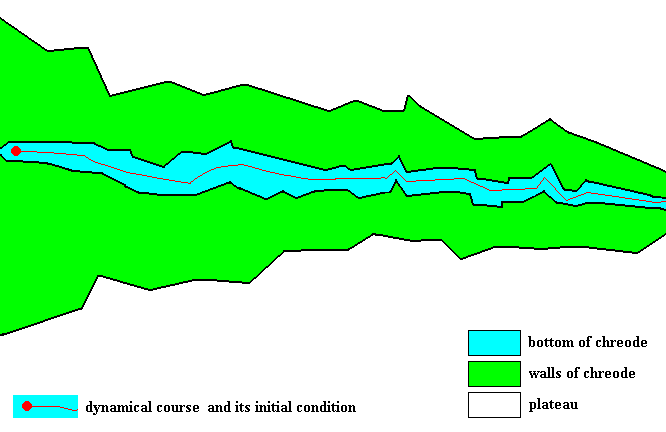
Figure 1 : ( = Figure 14 of previous document). One-entry chreode.
A chreode, drawn as an altitude map, which canalizes only one particular one-off dynamical course resulting in the generation of the corresponding particular one-off morphological structure, for example an arm of a dendritic snow crystal. The blue area is the bottom of the chreode and this bottom gradually slopes down from left to right. This type of chreode contains only one possible dynamical course to follow. We further assume that there is only one initial condition (that is, not several possible such conditions) that directly (and right from the beginning of the bottom course) sets the system in motion resulting in its following the one possible dynamical course of the chreode. Therefore we call such a chreode a one-entry chreode.
The green areas at both sides of the bottom are the lateral slopes of the chreode. If the system is perturbed in such a way that it deviates from its course along the bottom and then finds itself on one of these slopes, the system will thereafter quickly return to its orginal course along the bottom. This bottom is narrow while the slopes are relatively steep, so a transverse section through the 'valley' is V-shaped. At the bottom of the chreode we have drawn (red line) the canalized course (from left to right) taken by the succession of dynamical system states as this succession is determined by the If-Then constants that make up the chreode.
This Figure should not suggest that the Implicate Order contains only one such chreode. That would contradict the the fact of the enormous morphological diversity as we see it in the Explicate Order (think of organism and crystals). There are a great many 'species' of this brand of chreode, each determining or guiding the development of one particular material structure, different from that which is determined by another such 'species'.
The next Figure depicts a chreode which determines not one, but a whole bundle of possible dynamic courses (each consisting of a continuous succession of dynamical states, and each state consisting of a number of simultaneous [sub]conditions (not drawn), each one of them in turn triggering the activation and application of a series of successive If-Then constants [not drawn] ). Only the possible pathways followed by the succession of dynamical states are drawn, and from them only seven. Either the one or the other course is followed, depending on the initial condition.

Figure 2 : ( = Figure 15 of previous document). Many-entry chreode.
A chreode, drawn -- like in the previous Figure -- as an altitude map, a chreode which does not canalize just a single one-off dynamical course (like the one-entry chreode of Figure 14) but a bundle of possible and similar ones (all going from left to right), of which seven are drawn and which are canalized one at a time, that is, one or the other, depending on the prevailing initial condition (red disc). The morphological products of these courses are, it is true, different but simular enough that all of them belong to one and the same morphological type. Because there is room for more than one possible dynamical course, that is, for several different possible courses, the chreode bottom is relatively broad (broader than the one depicted in Figure 14), and its transverse section is more or less U-shaped (like a valley carved out by a glacier instead of just a river or stream). There are, consequently, more than one possible initial conditions (in the present example numbered 1, 2, 3, 4, 5, 6 and 7), such that each one of them directly (and right from the beginning of the bottom course selected by it) sets in motion the main possible dynamical course (of the chreode) of which it is the initial condition. Therefore we call this type of chreode a many-entry chreode.
So this kind of chreode determines not a particular one-off morphology, but a whole morphological type. Such a type consists of many one-off morphologies together constituting the type. So when such a chreode is followed a few times the morphological results can be more or less different, but they still belong to one and the same morphological type.
Like in the previous Figure the blue area is the bottom of the chreode and this bottom gradually slopes down from left to right. The green areas at both sides of the bottom are the lateral slopes of the chreode. If the system is perturbed in such a way that it deviates from one of its courses along the bottom and then finds itself on one of these slopes, the system will thereafter quickly return to its orginal course along the bottom.
Also this Figure should not suggest that the Implicate Order contains only one such chreode. That would contradict the the fact of the enormous morphological diversity as we see it in the Explicate Order (think of organism and crystals). Also here there are a great many 'species' of this brand of chreode, each determining or guiding the development of one or another material structure belonging to some definite morphological type, different from that which is determined by another such 'species'.
Indeed, any alternative dynamical route -- implied by every case of regulation and many cases of regeneration -- must lie outside the chreode as this chreode was given in the above Figures. Only in cases of small perturbations (which are expected to take place often during the morphogenesis of an organism) the system remains within the chreode : it climbs up one of the valley walls, it is true, but only for a short distance and then directly returns to the original dynamical course. In order to legitimately call certain observed phenomena in morphogenesis regulation or regeneration, the alternative routes associated with them lie, as has been said, outside the chreode. And because we can expect that in biological morphogenesis not only the 'normal' dynamical course (lying at the bottom of the chreode) but also these possible alternative dynamical courses (lying totally outside the chreode) are stable, that is, insensitive to not too strong a perturbation, also these possible alternative dynamical courses must be chreodes themselves, that is, must lie at the bottom of another chreode (a chreode different from the 'normal' chreode). But how has this extra chreode originated? This question is, it must be admitted, hard to answer. Earlier we have said that a chreode is formed as soon as the same sequence of consecutive system states repeatedly has been passed in the Explicate Order. In the Implicate Order this means that the same bundle of If-Then constants is repeatedly activated and applied to the Explicate Order. But such repetitions are not very probable to occur in the case of the mentioned alternative dynamical courses because the corresponding perturbations that inflict severe damage are, if they occur at all, not expected to be exactly repeated, and some of them did certainly not happen at all in the biological history of the Earth (or of the Universe for that matter), for instance the many cases of experimentally having removed or damaged an organ or tissue of a developing organism in the embryologist's laboratory. So how have these alternative dynamical courses become stable courses, that is, have become chreodes? Our anwer can only be speculative. Let's try.
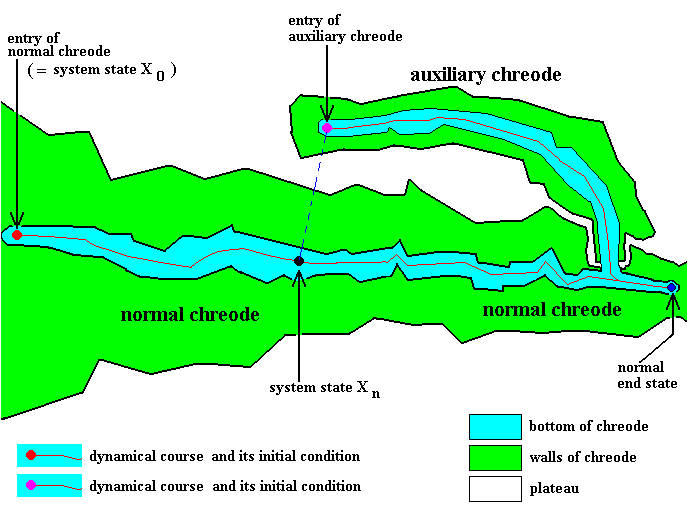
Figure 3 : One-entry normal chreode possessing a one-entry auxiliary chreode, reflecting the given developing organism's capacity to regulate its morphogenetic course.
Apart from small detours, dynamical courses all go from left to right (red track), that is, we must read the diagram from left to right.
Small red disc (at the far left) : Entry ( = initial condition) of normal chreode.
Small black disc : The normal chreode guides the normal dynamical course along its bottom. This dynamical course consists of a great many successive system states, of which the black disc is one of them, namely system state Xn .
Small purple disc : Entry ( = initial condition) of auxiliary chreode.
Small blue disc (at the far right) : Stable end-state of dynamical course (whether this course is deflected or not).
This determinative structure (that is, the particular one depicted in the Figure) embodies the capacity of the developing organism (for which the structure stands) to regulate its course when, at some stage of normal development (say at system state Xn), it is perturbed such that it results in serious damage inflicted upon it.
When a particular serious damage is indeed inflicted upon the developing organism at system state Xn , this system state is destroyed. It has then become the necessary and sufficient condition for the system to jump to the entry (that is, initial condition) of the auxiliary chreode (purple small disc) and then to follow the stable alternative dynamical course (red track starting from this entry) until it debouches into the normal dynamical course again. In this way the normal end-state is still generated in spite of the damage inflicted upon some intermediate system state.
The determinative structure as given by the figure (partly) represents the whatness of the given organism insofar its morphogenesis and the possible potentials of the latter are concerned.
Of course, damage inflicted upon a developing organism can, from case to case, result in many different necessary and sufficient conditions for the system to jump either (depending om the structure of the given chreodic landscape) to the entry of one or another auxiliary chreode (that is, in this case there are supposed to be several different auxiliary chreodes connected with the given normal chreode), or to one or the other entry of the same many-entry auxiliary chreode. In addition it could also be (still depending on the structure of the given chreodic landscape) that the auxiliary chreode has only one and the same dynamical route along its bottom but that, nevertheless, several different initial conditions give courses that all converge on that same dynamical route. This last case is depicted in the next Figure.
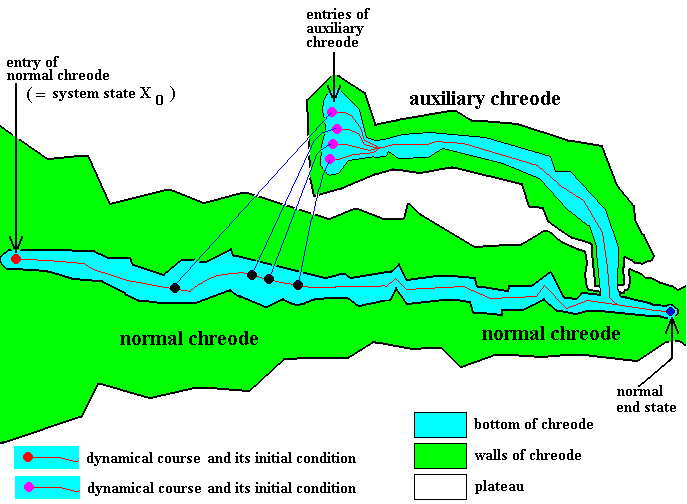
Figure 4 : Case of a one-entry normal chreode having one many-entry auxiliary chreode, also reflecting the capacity of some given developing organism to regulate its morphogenetic course.
The many-entry auxiliary chreode has several direct initial conditions, but they all lead to the same one dynamical course going along its bottom.
Apart from small detours, dynamical courses all go from left to right (red track), that is, we must read the diagram from left to right.
Many different possible damages that could be (the one or the other) inflicted upon the developing organism create many different (one or the other) necessary and sufficient conditions (small black discs) for the system to temporarily leave the normal dynamical course or path and jump to one or another entry (small purple disc) of an auxiliary chreode and then temporarily follow the alternative dynamical path, but nevertheless eventually ending up at the same end-state as that to which the normal path leads.
As is to be expected (from observations of many cases of morphogenesis in diverse organismic species) a normal chreode could be such that it has not just one but a great many auxiliary chreodes. Indeed, there are many different sorts of damage that trigger correspondingly different alternative dynamical courses to follow (that is, activate different auxiliary chreodes being possessed by the same normal chreode, or follow different dynamical courses that are contained within one and the same auxiliary chreode).
The determinative structure as given by the figure (partly) represents the whatness of the given organism insofar its morphogenesis and the possible potentials of the latter are concerned.
From all this it is clear how we should describe different cases of morphogenesis in which different degrees or ways of regulation are found (by embryological observations and experiments) to be possible, describe that is, completely in terms of chreodes (and therefore ultimately in terms of If-Then constants).
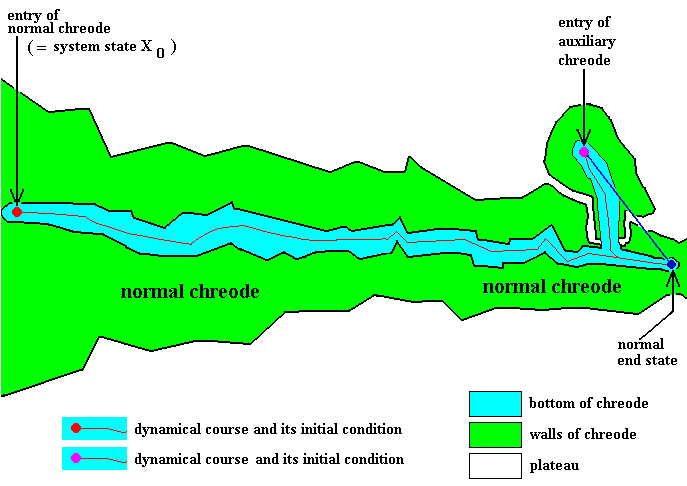
Figure 5 : Regeneration. Case of a one-entry normal chreode having a small auxiliary chreode representing the capacity of the developing organism to regenerate parts that were lost after morphogenesis was completed.
Of course the inflicted damage of the fully developed organism could be so much that only a small part of the body is left. Most organisms cannot in such a case regenerate the lost body parts, but flat-worms can. We will then have to do with the morphogenesis following a true alternative course of the dynamics -- like we have in regulation -- in the end producing the whole organism again. In the present Figure we imagine the normal end-state (small blue disc at the right) of the dynamical system (that generates the given organism) being more or less severely damaged, resulting in body parts being lost. When indeed such a serious damage is inflicted upon the end-state of the developing organism, this state is destroyed. It has then become the necessary and sufficient condition for the system to jump to the entry (that is, initial condition) of the auxiliary chreode (purple small disc) and then following either a short course directly back to the end-state, or following a longer and stable alternative dynamical course until it debouches into the normal dynamical course again. In this way the normal end-state is generated again and the lost parts regenerated.
Of course there can be many such auxiliary chreodes possessed by a given normal chreode, or one which possesses one or several many-entry auxiliary chreodes to make possible all kinds of regeneration that might be needed.
The determinative structure as given by the figure (partly) represents the whatness of the given organism insofar its morphogenesis and the possible potentials of the latter are concerned.
During normal morphogenesis specific body parts are being generated by dynamical subsystems. This is more or less analogous to the case of the development of a branch or branches on a developing crystal (as we see it in many snow crystals). But while in crystals this development of branches is caused by a change in local external conditions, in a developing organism it is caused by internal and intrinsic local changes : A certain state of the overall dynamical system generating the organism brings with it that a particular segment of the genome of a number of already developed cells becomes readable and is indeed read. The special proteins then synthesized in such cells cause the group of these cells to become the initial condition or entry of a separate chreode of which the succession of states leads to the formation of a specific body part. See next Figure.
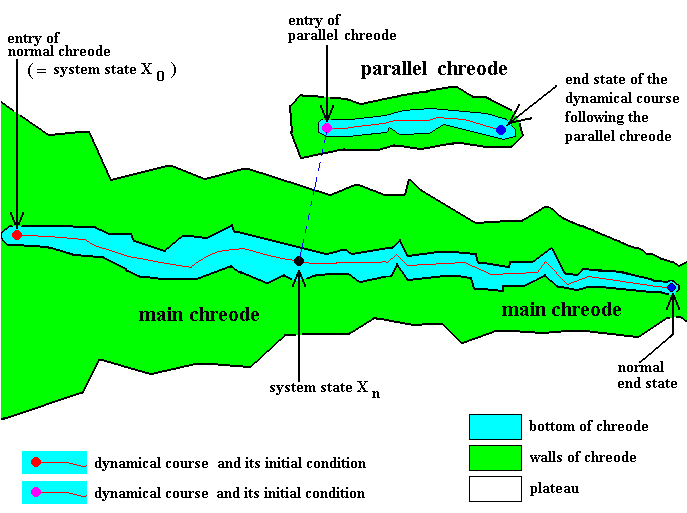
Figure 6 : The capacity of the developing organism to generate a specific body part is here expressed in terms of chreodes.
A certain state (here given as system state Xn ) of the main dynamical system brings with it the formation of a special initial condition for activating a parallel chreode. The dynamical course of the dynamical subsystem associated with this parallel chreode is then followed, leading to the generation of a specific body part.
At the same time the course of the main dynamical system is followed also.
The initial condition corrsponding to the entry of the parallel chreode belongs to the overall (and simultaneous) system state, that is, to the corresponding state of the morphogenetic main dynamical system, in the present case to the system state Xn , because that entry is only a local subcondition of the corresponding overall system state. They are simultaneous, and therefore they also belong to the same corresponding World State. Also the ensuing states that appear along the trajectory in the parallel chreode are simultaneous with the corresponding further states of the main dynamical system. So in the case of the generation of body parts (such as leaves, legs, wings, internal organs, etc.) we have not to do with alternative dynamical courses or routes, but with the following of simultaneous dynamical subcourses guided by separate parallel chreodes, which chreodes definitively belong to the normal development of the given organism, and which will be activated during the course of the main dynamical system. The parallel chreodes are not alternative chreodes, they are not activated instead of some portion of the main chreode but together with it.
A main chreode is always accompanied by a 'swarm' of separate parallel chreodes : The dynamical course of at least every type of organ of the developing organism is guided by a corresponding parallel chreode. All these chreodes, that is, the main chreode and all the accompanying parallel chreodes, belong to one and the same chreodic landscape characterising the normal morphogenesis of a given organism. But in the end also all alternative chreodes making possible regulation and regeneration belong to this chreodic landscape.
BIOLOGICAL EVOLUTION
In the present and foregoing documents we have set up a theory about a possible metaphysical background -- more precisely, an ontological background -- of the world of observable things and processes. And, until now, this theory seems to do well. Elementary, 'atomic' states of affairs, that is, elementary conditions, as they happen to prevail at some place or places and at some time or times in the Explicate Order are 'injected' into the Implicate Order and reside there as 'noëtic' (that is, thought-like) contents (formal contents). They directly and automaticly form the ' If-parts' (or If-components) of corresponding If-Then constants, constants which as such reside in the Implicate Order and make up the determinative structure of the Explicate Order. Successive activation of such If-then constants (the consequent of the one forming the condition for the next, etc.) is then seen in the Explicate Order as a causal chain of events.
Until now the developed theory of ours has adhered to the philosophy of reductionism. This philosophy holds that because, as assumed by it, any macroscopic event or material structure as it can be seen in the Explicate Order is built up from, or composed of, elementary basic units (such as atoms or molecules), it can be fully explained by those units and their interactions : If we have knowledge of the nature and the number of the basic units which participate in such a macroscopic event or material structure, this knowledge is sufficient to undestand the latter completely. That is, any more or less complex structure can, insofar as explaining it, be reduced to general basic constituents and their interactions. As far as we consider purely physical and chemical events this might be true, and maybe it also holds for all cases of organic individual development (morphogenesis), but for many, if not all, events and processes making up the evolutionary history of organisms (at least with respect to those organisms that stand above bacteria and the like) such a reductionism can no longer be adhered to. This is especially true with respect to the many 'sophisticated' adaptations that have been evolved in almost every plant or animal species. Such adaptations sometimes look like 'smart solutions' of certain biological problems as they were posed to many, especially animal species when their external conditions have been changed, that is, when a change in their biotic or abiotic environment has taken place. These adaptations not only concern morphological or physiological 'aswers' to changing conditions but also features of behavior. And they are often 'smart' and 'prescient' not only in animals possessing an advanced nervous system like the higher vertebrates (such as mammals) but also already, and even to a high degree, in, say, insects (especially in the Order Hymenoptera -- wasps, bees and ants [not only in the social species of Hymenoptera but also in species of them that live solitary] ). In the majority of cases such adaptations seem to be such that they cannot be explained purely in a reductionistic way, and here I do not mean that they 'cannot yet be explained' by such methods, but that it seems that they cannot be so explained even in principle.
Although the current view is that all evolutionary changes can in principle be completely explained by the natural selection of the most fitted genetic mutants of a given species or population, (mutants, that is to say, selected from a multitude of genetically different individuals, where the mentioned most fitted genetic mutant is one of the results of the many different random mutations having taken place in different individuals of a given population in the course of time), it becomes more and more clear that this cannot be the only mechanism by which evolution works. When we follow just a single changed character or feature as it is present in the individuals of some given group of animals, it is often relatively easy to explain the evolutionary change which led to the current version of that character by (appealing to) natural selection of some mutant carrying that present version which is, with respect to an earlier version, considered to represent an improved version of that character. When such a process is repeated it will finally result in a highly developed state or version of the original character, for example the evolutionary pathway leading from mere light sensitive spots to genuine eyes with lens, retina and iris.
However, as soon as we consider the whole organism, that is, not solely the evolutionary development of the visual capacity of that organism, but all its other evolutionarily developing capacities and features as well and at the same time, and in a changing biotic and abiotic environment, we will see that all simplicity is lost : a character of an organism does not and cannot evolve on its own but only in concert with, and in dependency of, many other, if not, all other characters of that organism. Now we cannot explain things anymore with the simple model of random mutation plus natural selection, although in some way it will often play a part. A 'smart' adaptation (morphological, physiological, or behavioral) cannot evolve by just a lucky consecutive series of relevant advantageous mutations that happened to take place at the orgamnism's genome (hereditary apparatus -- DNA) and are then being consecutively picked out by natural selection, (cannot so evolve that is) totally without conflicting effects on other parts of the body of the animal or on other aspects of its behavior. When we nevertheless keep on to proceed in this way -- explaining things exclusively by the random-mutation-selection model -- we will get buried under a mass of speculative ad hoc and often conflicting assumptions that have to be made, or else must limit ourselves to the history of a single character which latter is wholly artificial and contrary to biological fact. Moreover, it has been found that there is not a universal one-to-one correspondence -- valid always and everywhere -- between a gene and a character (be the latter morphological, physiological or behavioral). Often, in individual development (morphogenesis), for the expression of a new character or feature (or an improved version of it -- improved with respect to a certain environment) not only genetics are important but also the external context of the given organism. So indeed, not always simply : same gene same character. Further, the greatest part of the genome of an organism is not used and thus not read by the cellular machinery. It is a remnant of the past. So most mutations will not even come to be expressed. And worse, most genetic mutations are unfavorable if not lethal. It is clear, therefore, that, in addition to random mutation plus selection, there must be other factors at work for at least all cases of 'smart' adaptations as we see them in all animal species. These other factors, however, cannot, as it turned out, be found in the world of elementary step by step chemical and physical changes. That is, although we see such changes at work, or can see them at least in principle, they cannot all by themselves account for the evolutionary emergence of new forms at least insofar as such forms represent 'smart' adaptations to certain biological circumstances (as we can see such adaptations for instance in the behavior of solitary wasps, a behavior guaranteeing the succesful development of their offspring, but in fact as we see such adaptations everywhere in the animal kingdom). It seems that the chemical and physical machinery working according to natural laws -- which cannot be changed -- must in some way be harnessed (that is, over-formed) in order that it eventually results in the formation of such adaptations in organic evolution. How then is this machinery so harnessed?
It is here where we let our postulated Implicate Order come in. But we can do so only when we allow for certain concessions to be made with respect to the general ideas concerning it as these ideas were, until now, given in this and in the foregoing documents.
The space-time world, that is, the material world we actually observe (directly or indirectly), and which we have called the Explicate Order, floats, as it were, on, or is an expression of, an underlying world which is not extended along space and time dimensions : the Implicate Order, that is, a world in which things and events are enfolded. We have already 'seen' that 'atomic' (that is, elementary or basic) formal contents of physical or chemical conditions that actually exist (but then extended along space and time dimensions) in the Explicate Order will be enfolded (that is, injected) into the Implicate Order -- while not disappearing from the Explicate Order -- where they then form the If-components of If-Then constants. The consequents of these constants can then be unfolded (that is, projected) into the Explicate Order (without disappearing from the Implicate Order). They then unfold along space and time dimensions.
However, if we wish to account for the above mentioned biological events as they are observed in the Explicate Order ('smart' adaptations and other events in organic evolution) in terms of the properties of the Implicate Order we must assume that not only the formal content of atomic, that is basic or elementary, physical or chemical conditions as they occur (as extended along space and time dimensions) in the Explicate Order can be injected into the Implicate Order (and there residing as mere formal contents, not extended along space and time dimensions), but also certain other non-atomic, that is not elementary, contents. Which contents?
Well, we shall assume that all higher-level contents that are, as they occur in the Explicate Order in an explicit space-time framework, relatively stable, such as the content of an organic species or of a crystal species, can and even will, also solely as formal contents, be injected into the Implicate Order. In this way -- and this is one of the above mentioned concessions we have to make -- the content of the Implicate Order (that is, everything that is in this Order) is in some respects dependent upon the Explicate Order : Every organic species, for example, is not something which has always existed, so its formal content can only be actually injected into the Implicate Order when such a species has appeared in the Explicate Order. If we were to hold that, in adition to all actual, also all possible (stable) contents, and thus also the contents of all possible organic species, are already present in the Implicate Order, we would fall back into one or another teleological theory (in which everything is considered to be already 'forseen' by Nature and thus in some way pre-existent). All versions of such a theory have been refuted on reasonable grounds. So we will hold that at least the organic world is actually evolving, resulting in the formation of new patterns, new not only in the Explicate Order but also in the Implicate Order. But because we need the (concept of the) Implicate Order to account for the mentioned biological evolutionary processes as we see them in the Explicate Order, we must assume the taking place of processes not only in the Explicate Order but also the 'taking place' of analogues of them in the Implicate Order, and this in addition to the mere 'following' by it of If-Then constants. The 'processes' as they take place in the Implicate Order must consist in 'interaction' of, or 'reactions' between, not material entities (such as between material organic individuals, or between such an individual and its physical and biological environment), but between their noëtic counterparts, that is between the corresponding pure (i.e. formal) contents of these entities, which formal contents will then look in the Implicate Order like mathematical entities in which indeed the space and time dimensions are enfolded into, or represented by, mere parameters or specified variables contained in these mathematical entities. Or we could see them as analogues of thoughts (that's why we called them noëtic counterparts) or logical entities for that matter. Such a noëtic reaction between two or more different formal contents, as it can take place in the Implicate Order, can, as we assume, result in noëtic reaction products analoguous to, for example, what happens in chemical reactions. And of course there will be cases in which some given formal contents do not react with each other at all, as is also often the case in chemistry. A noëtic reaction product, which itself is just a formal content, will then be unfolded, that is, projected (into the Explicate Order), only at that place or those places in the Explicate Order and only at those times in that same Order where there are no factors that totally forbid such a projection.
As we have said, the pure content (formal content) of an existing (in the Explicate Order) organic species, can and will be injected into the Implicate Order (without thereby disappearing from the Explicate Order) and reside there as a purely noëtic entity. It does not reside in the Implicate Order at a particular place in it or at a particular point in time. As a noëtic entity (i.e. a purely formal content on its own) it is spaceless and timeless. However, the content of such an organic species as it exists (in the form of one or more populations of living individuals) physically in the Explicate Order necessarily includes the content of its precise ecological niche, because every organic species is inseparately connected with its ecological niche and forms with it one integrated whole. Indeed, the content of this niche is only conceptually, that is, only in the context of scientific method, separable from the content of the corresponding organic species. And not only the content of the actual ecological niche in which the species now lives is inseparably connected with the content of this species : This same content of that species is also so connected with its possible, that is, its potential, other ecological niches. By the expression "an ecological niche that is, or can be, occupied by a given organic species" we mean the total of precisely those specific features of that species' environment to which (specific features) it is, actually or potentially, exactly geared. These features consist of temperatures tolerated by living individuals of that given organic species together with other physical and chemical conditions. But above all they consist of existing food resourses present in the environment or habitat in which the representatives of the mentioned organic species live, and also (they consist) in the presence or absence of predators and other biological factors relevant to that species.
And because in this way the content of actual and possible ecological niches of a given organic species is clearly delimited and thus definite, it is a genuine content, and because it is connected with the content of one or another organic species which as such is a relatively stable content, the content of its ecological niche (actual or potential) is also relatively stable and can, and will, like the content of the organic species itself, be injected into the Implicate Order. So we can have, say, two, injected (formal) contents, (1) the (formal) content of the organic species and (2) that of one of its potential ecological niches. And these two contents, as they reside in the Implicate Order as noëtic entities, can noëtically react with each other yielding a noëtic product, which might unfold, that is, be projected, into the Explicate Order. Or we can have the injected (formal) contents of two different organic species which, as also residing noëtically in the Implicate Order, then noëtically react with one another resulting in some noëtic reaction product which might be projected into the Explicate Order. Or, as yet again another possibility, they will not noëtically react with each other as the case may be.
Example
Let us, to make things clear, give a crude, that is, a more or less simplified, example concerning the evolutionary emergence of bloodsucking mosquitos (from non-biting flies) and the consequent emergence of malaria in humans :
There were and are many different species of the suborder Nematocera of the two-winged insects (Diptera). The set of diptera constituting this suborder distinguishes itself by the fact that its members always have their antennae as long multisegmented thread-like organs, while the rest of the Diptera (Brachycera orthorapha and cyclorapha) has short antennae in which many segments are fused together such that the antennae now consist of only a few (mostly three) segments forming in this way well-integrated antennae. Initially, that is, at some time far back into the geological past, the suborder Nematocera did not contain bloodsuckers. We will now concentrate on one of these initial species. Its larvae are aquatic and feed on bacteria and organic particles that abound in fresh water. The adult (winged) insect developing from such a larva is aerial and does not take food anymore or just some nectar from flowers to fuel its flight muscles. After mating eggs will be laid in or on the water and the cycle begins again. The food reserves (proteins) built up in the developing larva are just enough to produce a batch of eggs in the female adult fly. She can lay just one such batch of eggs, and no more. However, in the fresh waters where the larvae live many predators abound, so the majority of the eggs will not result in adult insects. The continuance of the species could be in danger. One way out of this dilemma is for the female to be able to produce and lay yet more eggs so that enough of them will survive all the way up to adult insects to maintain the species. But then the larva must stay longer in the water in order to build up corresponding larger food reserves (proteins). And to stay longer in the water increases the risk of being eaten, or of dying as a result of possible total evaporation of the water basin before larval development has been completed. So perhaps it would, alternatively, be favorable for a secured continuance of the species that the female herself will feed for a while on protein-containing food (instead of just feeding on nectar) in order to be able to produce and lay a second and perhaps third batch of eggs.
Thus, the a c t u a l ecological niche for the described species of Diptera was :
But now the new situation gives a yet a further possibility : The combination of biting blood-sucking mosquitos and the presence of warm-blooded animals is a possible ecological niche for an internal microscopic parasite. Although such a microscopic parasite can live within the blood of a vertebrate it cannot disperse itself over more or less large areas. But having now blood-sucking insects (mosquitos) who visit one vertebrate individual after another, the microscopic parasite has a means of dispersing itself. It only has to adapt itself to a temporary life within the mosquito, that is, it must be such as not to cause, or else being resistant to, any physiological or biochemical defensive reaction of the insect against it.
So we can imagine that a noëtic reaction between
(1) the formal content of the newly formed species, that is, the formal content of the originated species of mosquito,
and
(2) the formal content of its (now) actual ecological niche (fresh water for the larva , the presence of warm-blooded vertebrates, and adult insects able to find them and to suck blood from them),
will take place in the Implicate Order and that the resulting noëtic reaction product is the formal content of some microscopic blood parasite that can live in the blood of warm-blooded vertebrates and that also can live, at least temporarily, in the body of the blood-sucking mosquito.
When at the relevant places and times in the Explicate Order there are no factors prevailing that forbid projection, the formed noëtic reaction product will indeed be projected into the Explicate Order, and this projection we will experience as the appearance of a microscopic parasite which develops itself in vertebrate blood and which disperses itself by means of blood-sucking two-winged insects : mosquitos.
This microscopic internal parasite might cause a disease in an infected vertebrate, but this need not to be malaria, because that probably depends on the details of the nature of the mosquito species and of the special chemical and biological nature of the original parasite. That is, when the mosquito species was indeed a species which we now reckon to the genus Anopheles and when the vertebrate animals from which the blood is taken are humans, and all this together with some other conditions being fulfilled, the microscopic parasite will be the one that we now know to cause malaria in humans.
(End of example).
There have been also very long evolutionary sequences involving many steps, each of them for a more or less longer period represented in the Explicate Order by a new organic species.
Expressed in terms of the Implicate Order we can then say that after the initial organic species is connected with a new ecological niche as its actual niche and as such represented in (that is, projected into) the Explicate Order, the organic species' formal content can, in the Implicate Order, noëtically react with the formal content of this new actual ecological niche, resulting in a changed formal content of the organic species, turning it into a new species, which will then be projected into the Explicate Order. Then this new formal content of the new organic species can again noëtically react with the formal content of the same actual ecological niche, resulting in a further change of the formal content of the species (resulting in yet another species), and this, after being projected into the Explicate Order and at the same time also remaining -- as formal content -- in the Implicate Order can noëtically react with the formal content of the same actual ecological niche ....., and so on and so on.
The species or population and its ecological niche.
An ecological niche (actual or potential) of a given animal- (on which, instead of plant-) we here concentrate) species (or, as the case may be, subspecies, or local population) is that part of its natural environment, or that collection of aspects of that environment, to which the specific morphological and physiological constitution of individuals of that species (subspecies, local population) is precisely and specifically geared. Such a niche is thus an (actual or potential) extension of the mentioned constitution of that species. And like the formal content of that species, the content of its ecological niche is precisely defined, and can therefore also be considered as a formal content. It is just as specific as that of the organic species which -- in the Explicate Order (actually or potentially) occupies that niche. As has been said, the most important feature of such a niche is the presence in it of appropriate food in sufficient quantities. This means that the niche is at least for a large part a biochemical extension of the constitution of the species that occupies it (note that individuals live in their environment, but occupy a definite ecological niche in it). The other important feature is the presence or absence of predators and other relevant biological factors.
The actual ecological niche is the one presently occupied by the given organic species. The latter is sufficiently adapted to this niche. Repeated noëtic interaction between the formal content of the species and that of this actual ecological niche could lead to further adaptation and perhaps eventually to over-specialization.
A potential or possible ecological niche of that same given organic species is a niche to which the species is not completely adapted, but the constitution of the species and that of the potential niche are such that the species can relatively easily evolve the necessary supplement of adaptation, without side effects that conflict with the extended adaptation. This necessary supplement of adaptation will be realized by the projection (from the Implicate Order) of a noëtic reaction product having resulted from the noëtic reaction between the formal content of the organic species and that of the potential ecological niche. The fact that all this will happen when in the relevant place (and time) in the Explicate Order such a niche is present determines the latter to be a potential niche for that given organic species. The latter consists -- as do all organic species -- of a collection of more or less spatially dispersed populations of living individuals. And only that part of the species will adapt to the potentially new ecological niche of which the individuals belong to a population that lives in such a place that it is in contact with that potentially new ecological niche. And this new adaptation can, when the population is sufficiently isolated from the rest of the species, represent one of the factors which together are responsible for a new biological species to appear. And because the other part of the species remains -- ex hypothesi -- unchanged we witness the splitting up of the one initial species into two species of which one of them is new. And when the branched-off population happened to constitute a large, that is, substantial, part of the species as a whole as it was then represented by its individuals, there is a chance that the gene pool of the total of left-over populations is different from the gene pool of the total of the populations before the splitting up. And this could mean that then also the left-over of the original species now belongs to a different species, that is, different not only from the above described new (branched-off) species but also from the original species. Two new species have then evolved. However, when the above described left-over populations (successfully) keep on occupying their original ecological niche, we might question their supposed change into a new species despite the difference of their gene pool from the orginal gene pool. If we then do not accept (or believe) that such a change has occurred (or if some changes did take place we are not prepared to interpret the resulting differences [from the original species] as constituting a new species), then not two, but only one new species had evolved.
Anyway, it is clear that it is ecology that drives evolution. And we will say more about it in the sequel.
Generating diversity as a way to continue organic life.
Life has been on Earth already during at least a billion years. In the course of this very long period of the Earth's history many changes of conditions have occurred : climatological changes, changes of the crust's relief (formation and deformation of mountain chains), other geological changes and events such as volcanism, further, changes in the land-sea patterns, possible changes in the chemical composition of the oceans and other water basins, changes caused by the bombardment of meteorites, etc. Many of these changes certainly wiped out many organisms. Nevertheless life is still abundantly present on the Earth. This can only be so if life in general is able to survive all these kinds of environmental changes and catastrophes. What general quality then life must possess in order to be able to survive? Well, according to me the type of life that we encounter here on Earth must be such that it intrinsically seeks to diversify itself constantly. That is to say, it must strive to inhabit or colonize all kinds of environments as they are present on Earth. If then one or another specific (type of) environment is destroyed as a result of one of the mentioned (great) changes or catastrophes, and all life (or just one or more animal or plant species) in that particular environment is wiped out, then other environments are still preserved and with them their life stock. And indeed, life is diversifying itself constantly as newly formed types of environment are eventually populated by adapted organic species, new species. An example was given above with respect to the emergence of blood-sucking mosquitos followed by the emergence of certain internal microscopic parasites. In fact we must be more precise : new environments contain new ecological niches to be occupied. But also within the same (type of) environment new (types of) ecological niches can emerge and they also will soon be occupied by living organisms, which are then new subspecies or even new species. And because of all this, life is very resistant to local annihilations, to universal destruction of a type of ecological niche, or even to the loss of a whole type of environment. Although many organic species will be lost when these things occur, many other organic species more or less closely related to the lost species will continue to live on in the other niches and environments not affected by the mentioned catastrophes. And eventually the new types of niche and environment created by these catastrophes will become populated by organic species specifically adapted to them, however harsh these new niches and environments have come to be. All this is evident on the basis of many observations which show that virtually all types of existing environments, however harsh they might be, have their own organisms specifically adapted to live in them. And this is what we meant (at the end of the previous section) when we said that it is ecology that drives organic evolution.
Anagenesis
However, in addition to (qualitative) diversification of life-forms there is, in organic evolution, yet another trend : increase of (the level of) complexity of the morphology, physiology and behavior of living organisms. When one given species of organism has, what one calls, a higher level of complexity of organization than another closely related (and therefore comparable) species of organism, this does not mean that the former is, thanks to its higher complexity, more successful than the other, less complex, organism in the sense that its continuance as a species is better guaranteed, namely by producing more offspring that reaches maturity. This is evident by the fact that many very simple organisms, that is, organisms with a low level of organizational complexity, are still living today, and often even in abundance. For an organism to be more complex than another is just for it to be different. As such it is just a case of diversification having taken place. Nevertheless it is a special case of diversification : We see that in evolution there is a general trend, although not with respect to all organic species, towards complexification. We can call the evolutionary process of complexification of organisms (the process that leads from 'lower' organisms to 'higher' organisms) anagenesis. It is clearly visible in the evolution of the Vertebrates. Here it leads from fishes to amphibians to reptiles and birds and finally to mammals. The mammals are evidently the 'highest' vertebrates (while fish still exists, even abundantly, also with respect to the wealth of species). But it is just as clearly visible in Insects, where the higher and most recent orders are the beetles, the two-winged insects (mosquitos, gnats, and flies) and especially the hymenoptera (wasps, bees, and ants), which (orders) 'stand above' the lower and more ancient orders like the mayflies, the dragonflies, the orthoptera (locusts, crickets, grasshoppers, cockroaches) and the orders of primarily wingless insects. And also within such an order the most recent forms are the most complex. In the order hymenoptera this is very conspicuous especially with respect to the diverse instincts which are very complex in the higher (and more recent) members of the order (higher solitary wasps and bees, and, of course, social wasps and bees, and ants).
Anagenesis, as it is scattered all over organic evolution, really is an unsolved mystery as soon as we realize that it does not as a rule represent an improvement of life of the organisms having undergone anagenesis. The phenomenon as such cannot be wholly explained by random mutation of genes plus natural selection. It also cannot be so explained by the colonization of new ecological niches. Therefore we must locate the very tendency in organisms of anagenetic evolution directly within the Implicate Order, that is, in the noëtic domain. Accordingly we assume that the noëtic counterpart of anagenesis -- noëtic anagenesis -- takes place in the Implicate Order and is even the rule there insofar formal contents of organic species are concerned.
Let us speculate how.
First of all every formal content of an organic species -- as it is, as a noëtic entity, present in the Implicate Order (injected into it from the Explicate Order) -- will, among other things, r e p e a t e d l y react with the formal content of its actual ecological niche, and this leads to a progressively better adaptation to that niche, or it leads even to overspecialization which is, when projected into the Explicate Order, an evolutionary dead end. Such noëtic reactions, however, do not necessarily result in the anagenesis of the content of given organic species, they just result in a different formal content (and thereby increasing diversity), not necessarily a more complex one (Note that we assume that when noëtic 'reactants' form a noëtic 'reaction product' the original reactants are not lost, because in the noëtic world there is no such thing as consumption).
Genuine anagenesis (that is, evolutionary development of some given species with (relatively) low-level organization -- morphological, physiological, or behavioral -- into one with high-level organization) is in fact a process of integration of the content of the species, that is, the content as it is present in every individual of that species, with the content of (apects of) its extended phenotype. The latter is the precise ecological of the given species. Although the content of this niche in fact is part of the complete content of the species when taken in the broad sense, it is the 'extended content' of the species when taken in the narrow sense, because this content extends beyond the boundary of the individuals, that is, beyond the boundaries of their physical bodies, making up the species. So at least part of this extended contend is external (but still intrinsic) to the content of the phenotype-proper of the species. Well, we assume that the process of anagenesis consists in 'internalizing' the extended portion (or part of it) of the species's total content. And we can be assured that most of the species's extended content consists in the preparation to, and the realization of, reproduction of the individuals of the species (production of offspring), that is, in the continuance of that particular species. For example in vertabrates have fish. A very external mode of reproduction is widely distributed among them : The female liberates eggs into the surrounding seawater. They are then fertilized by the corresponding males, and then left alone. Only a small percentage of these eggs will have the chance of developing into fish. On the other hand we see, namely in most mammals, an extreme form of internalization of the reproduction process : not only the young are taken care of (protected from dangers, and so on) like we see it already in birds, but also the development of the embryo in the egg takes place inside the body of the mother. With social animals this is even taken a step further : The young do not leave the herd even when they have fully grown up. In human society, but also already in social insects such as collectives of ants, the internalization process has gone so far as to bring in, into the habitat of the collective, things like food reserves and appropriate physical living conditions like temperature and perhaps humidity. These were examples of anagenesis. And indeed they do not necessarily represents improvements for sustaining life and for reproduction, because their alternative counterparts, that is, related species that have not undergone these internalizations, still live today and often in great abundance : Fish, for example, are still everywhere in the oceans [and in fresh waters], and are represented by many different species. Also most insect species are not social and do not even care for their young at all [as some non-social insects do] ) but are still abundantly present in a great variety of terrestrial environments. In cases of anagenesis there will be other aspects (orther then connected with reproduction) that are internalized, but, as has been said, the largest part of the external content of a species is connected with its reproductional activities. In insects, for example, there are many species in which the adult does not even feed anymore, it only takes part in reproduction and dissemination (spead the species worldwide or at least over large areas).
The 'drawing in' of some (part of) extended (external but still intrinsic) content into the content of the species-proper, that is, the full integration of the extended content with the content of the species, cannot, of course, as such take place in the Explicate Order. It must take place in the Implicate Order. There it is a noëtical integration of two formal contents. We could say that the extended formal content is noëtically absorbed by the formal content of the corresponding species converting the former to a part of the internal content. We could also express this as a nëtic assimilation of the initially extended formal content with the central formal content of the species. And although in the Implicate Order this happens with respect to the central formal content of all organic species (as they occur, or had occurred, in the Explicate Order) the result will project into the Explicate Order only there and then where it belongs and when no factors are present that forbid such a projection. And, of course, that is the reason why there are still so many organismic species around that have not evolutionarily developed a relatively higher level of organization.
In fact every adaptation of some given organic species to a certain aspect of its environment (biotic or abiotic) is a form of internalization of that environmental aspect into the content or form of that organic species which had acquired the adaptation. The environmental aspect is, so to say, reflected onto that content. Indeed, when observing individuals of such a species we can, as it were, 'read off' aspects of its environment, more precisely, aspects of its proper ecological niche. Such a process of internalization can be repeated or strenghened, and indeed, when a certain (not precisely defined) limit of intensity of internalization is crossed we have to do with anagenesis.
After some given organic species has internalized some particular aspect of its actual (or of one of its potential) ecological niche, it has secured new possibilities, that is, the domain of its potential ecological niches is expanded along a certain direction, a direction prompted by the specifics of the mentioned internalization. And precisely the added part of the domain (as a result of the expansion) can in turn be internalized, again resulting in a further expansion, and thus enlargement, of the domain of potential niches, an enlargement having proceeded in the same direction. As this goes on, the degree of internalization will steadily grow resulting in the anagenesis of the initial organic species (which has, possibly through an intermediate series of new species, progressed into an entirely different species). Often such a process will come to an end at that point where further internalization (along that direction) turns out to be impossible. If this happens early, the species has, it is true, changed but we cannot say that it has aquired an organization of a higher level than it had before. So then no anagenesis had taken place. But in most cases, however, the process of directed successive internalizations is allowed to go on and will then result in such a high degree of acquired internalization that we can justifiably speak of anagenesis.
All the single internalizations take place in the Implicate Order, that is, they take place in the form of noëtic reactions : The formal content of an existing (in the Explicate Order) organic species is present (as a result of injection) in the Implicate Order and resides there as a noëtic entity. Also the actual ecological niche of the species -- as its actual phenotypic extension -- is, as a noëtic entity, present in the Implicate Order, and, in the same way also all its potential ecological niches. In the Implicate Order some one aspect of the actual or of a potential ecological niche is internalized into the formal content of the species, and then the result of this internalization is projected there and then into the Explicate Order where it properly belongs and where no forbidding factors are present. And it is then that the domain of possible ecological niches of the species is expanded (or might be expanded) as a result of the species's newly acuired content. The newly added part of that domain is now injected into the Implicate Order together with the new central content of the species. This newly added part of the domain of possible ecological niches, as this part is now present -- as a noëtic entity -- in the Implicate Order, can now in its turn be internalized into the mentioned (new) central content of the species (also -- as a noëtic entity -- present in the Implicate Order) resulting in the next version of the central formal content of the species which is then projected into the Explicate Order, possibly resulting in yet another expansion of the domain of possible ecological niches of the organic species.
We locate all internalization processes, including most, if not, all processes leading to subtle adaptations of an organismic species to one or another aspect of its environment, in the Implicate Order, because it has become evident that they cannot be the result of just the mechanism of successive appearance of relevant (!) genetic mutations which are then singled out by natural selection. Certainly this mechanism plays a part in some way, but it cannot make up the whole process. It, that is, the mentioned mechanism, is what we actually seem to see (after having accepted some theories), and it therefore offers itself as a possible scientific, that is, reductionistic description and thus explanation of observed facts of 'internalizations' as seen in the higher forms of reproduction and in observed facts of subtle adaptations. But what we thus see in the space-time world -- that is, in the Explicate Order -- is just the result of the projection, and thus unfoldment along the space and time dimensions, of the products of noëtic reactions having taken place in the Implicate Order.
Rationality of organic evolution.
Of course what precisely we should understand by the "noëtic condition" in which 'things' are supposed to find themselves in the Implicate Order, and (what we should understand by) the "noëtic reactions" supposedly to take place in this Order cannot be precisely determined. They are part of our hypothetical extension of common Reality, an extension which is supposed to make up an order in which things exist as purely formal contents comparable to mathematical entities or thoughts. Indeed 'thoughts' form the best analogy to use when describing the content and nature of the Implicate Order. When we consider evolutionary accomplishments (as we see these in the morphogenesis of individual organisms and as we see them also in the many subtle adaptations and 'internalizations') we are struck by the structural 'rationality' of these accomplishments. Over the centuries this rationality has beguiled many researchers and philosophers into assuming a teleological nature of organic Reality. Today such theories meet with no response anymore, and rightly so, because they introduce a kind of providence into Nature and thus lead to religion instead of scientific explanation. Our theory also rejects teleology at the level of natural processes. It holds that the 'rationality' as we seem to see it in biological processes is not really genuine rationality but the mere following of natural laws. It is, however, a sign pointing to genuine rationality being present in some deeper ontological domain. And this genuine rationality is the very fundament of the things and events as we actually observe them. It determines how certain results, for example subtle biological adaptations, are accomplished by natural processes which do not depart from obeying natural laws. Precisely because any space-time process can only proceed according to such natural laws (each one consisting of a chain of successively triggerable If-Then constants as described earlier) it will almost always have to follow a long and convoluted path to finally realize a given particular structure such as a subtle biological adaptation. Even if we could unearth this long path it would, as it is in itself, not constitute a full explanation (or an even an explanation at all) because in addition to the knowledge of how precisely a certain biological adaptation is (chemically and physically) formed a full explanation also demands the answer to the question why precisely this particular adaptation has appeared in a given organic species. And then, indeed, high-level features (in contrast to just atoms and molecules) must be considered as fundamental entities, that is, as basic 'reactants' in certain processes. Such features are for example, as indicated above, the (formal) content of a given organic species, the (formal) content of its actual ecological niche, or the (formal) content of one or another of its potential ecological niches. And it is these considerations (high-level features figuring as fundamental units in a certain context) that lead us to the postulation of a noëtic domain steering as it were the physical and chemical processes as we observe them (to be so physical and chemical) in our space-time world.
Substance and anagenesis.
In First Part of Website we have explained what we should understand by substance in the metaphysical sense. A substance in this sense is a self-contained individual material entity which is an intrinsic unity, that is, it is what it is totally in virtue of itself. This, for example, in contrast to artifacts -- like, say, automobiles -- which are what they are in virtue of something else. A substance is a product of some dynamical system (or is such a system), and the dynamical law governing the system was called the essence of the substance. This essence is the fundamental central intrinsic content of the substance. And all determinations of some given substance (like its shape, its color, its chemical composition) which directly and necessarily flow from its essence are intrinsic determinations of it and as such belong to the intrinsic (but not central) content of that given substance. All other determinations of such a substance are not intrinsic, but extrinsic to it : they are (co)determined by factors outside the substance's content. This implies that the essence of a substance is that what it specifically is, and not what it individually is. It is a species, and is represented by all individuals belonging to this species.
For the inorganic world we have found that independently existing atoms and molecules can certainly be considered substances. But our best example is the crystal. Every crystal is a genuine substance in the metaphysical sense. It has an intrinsic shape (or sequence of them appearing during its generation), visible when the crystal grows in uniform and mild conditions of concentration and temperature, further it has an intrinsic symmetry, an intrinsic chemical composition, etc. There are many species of crystals, that differ with respect to these intrinsic qualities.
In the organic world it is individual organisms that are substances. Also here there are many species, each represented by existing individuals.
We now will speculate about the true nature of anagenesis ( = the evolutionary development of evermore higher-level organization and structure in organisms).
We already had speculatively established that anagenesis corresponds to a certain high degree of internalization of certain external but still intrinsic features of the given organic species. Such a high degree of internalization is the result of a longer or shorter chain of successive smaller internalizations in the same qualitative direction. What is the basis of this trend to internalize external (but intrinsic) features into the internal content of the organic species? Well, we speculate that any genuine substance -- as defined above -- because it is, in a way, a self, 'strives' to exist, as much as possible, on itself, which here means that all factors which co-determine what the substance in question intrinsically is, that is, all its intrinsic features, thus also all its external but nevertheless intrinsic features, are residing within that substance. Said, differently, it wants to integrate all its external intrinsic features with its internal intrinsic features.
When we now look to inorganic substances (in the metaphysical sense) such as crystals we see that there are only a few such external but intrinsic features or even none at all. For example when a crystal of a certain species grows at the bottom of a jar or of a natural cavity, or grows in contact with other crystals of a different species already present, it will adapt its shape to its surroundings, but such a resulting shape, although morphologically reflecting these surroundings, is not intrinsic to the given crystal species. So generally we can say that inorganic substances do not show any tendency, or only a slight tendency, to internalize external intrinsic features. The same is probably true of the lowest organisms such as bacteria. Or at least for them there is little to internalize. Things become quite different in more complex organisms. Such an organism, carrying of course its internal intrinsic features, has, in addition to them, many external but nevertheless intrinsic features as well : It lives in an ecological niche to which it is precisely geared. Especially this ecological niche contains many elements which intrinsically belong to the reproductive activities of the species's mature individuals, for example the appropriate substrate on which to lay their eggs, more or less hidden from predators, and when this regular substrate only serves as such a relatively safe site and not at the same time as food, there must be appropriate food around that can be attained by the young (or, in other cases, by their parents). So the more complex organisms indeed primarily (that is, from the start) possess many external intrinsic features, and it is in such organisms where we can expect the presence of the evolutionary tendency to internalize external intrinsic features.
Let us give some examples.
There are insects, such as mosquitos, which have specially adapted mouthparts and also a specially adapted physiology and behavior in order to suck blood from vertebrates. In this way a certain aspect of the animal's ecological niche is internalyzed into the content of the very species, here tranlated into corresponding morphological, physiological, and behavioral features. This is not yet a case of anagenesis but just a single adaptation to a newly acquired extension of the ecological niche of the ancestral non-biting flies. As another example many higher plants have flowers provided with a very special 'color' pattern not visible for us, but for bees. The bees are attracted to these flowers -- from one to the other -- and are guided by the pattern to the source of nectar in the flower resulting in the pollination of those flowers. So these plants have included bees in their pollination mechanism and now possess, as an intrinsic internal feature, a part of the vision capabilities and visual preference of bees in the translated form of a specific color pattern of their flowers. And some plant species, namely some orchids, go even further : their flowers morphologically imitate certain bees (or, in other species, certain wasps) which latter are then attracted to them because they look like potential mates. And because the insects repeatedly let themselves be deceived the plants are pollinated. Also this adaptation is not yet anagenesis but just one step of internalization. Anagenesis will have taken place when a whole chain of such internalizations has resulted from many successive 'incremental' simple internalizations all aligned along one and the same direction. The clearest examples are all the many existing forms of 'care for the offspring' as we see them in higher animals.
Evolution and the Implicate Order.
Although simple adaptations, and, consequently, simple internalizations, might be fully explained by the mechamism of random genetic mutations plus (natural) selection of some of them, complex adaptations cannot. In fact even most alleged simple evolutionary adaptations or (just) evolutionary changes can only be 'explained' when such an adaptation or change and the processes that resulted in them are considered in isolation, as if they were self-contained processes independent of the other biological processes going on in the organism and in its evolution. But it has turned out that it is impossible to consider them as single insulated processes. It is always the whole organism that is involved. And the cases of complex evolutionary adaptations and changes, that is, of complex internalizations, are not at all explicable by invoking the mentioned mechanism, because that would force us to accept a host of unbased assumptions in order, first of all, to have the relevant genetic mutations at our disposal, that is, their appearance at the right places and times, and then also (with such assumptions) to exclude all kinds of counteracting side-effects cogenerated during the evolutionary formation of such a complex adaptation or change.
From this one may conclude that, although the actual processes involved in evolutionary adaptations (and [just] evolutionary changes) totally obey existing fundamental natural (physical and chemical) laws, the resulting historical pattern of (micro) physical events and coupled chemical reactions, is so immensely complex that by no human intelligent effort it can be unearthed as a coherent complete chain of successive events (the one event necessarily flowing from the previous one), and even if one could do so (by a joint effort) one could not apprehend the result, that is, one could still not say that now the matter is fully understood and explained. But in this view the reason that we cannot explain things does not lie in those things (complex evolutionary adaptations and changes) themselves but in the limited capabilities of the brain to encompass vast schemes of logical or mathematical deductions or, even worse, vast schemes of microphysical and chemical reactions. It does not say that such complex reaction schemes do not actually exist, that is, the establishment of the impossibility of reductionistically describing, and thus explaining, complex evolutionary adaptations and changes is as such not a proof that they do not evolve along purely physical and chemical lines (that is, following existing physical and chemical laws).
However, the established impossibility of reductionally describing and thus explaining complex evolutionary adaptations and changes could be interpreted as being a fundamental impossibility (even for 'infinite' brains) and thus as such pointing to the fact that physical and chemical laws cannot, all alone by themselves, completely, that is, exhaustively, account for these evolutionary processes. And this latter view we shall here adopt.
We must therefore assume that primarily the internalization (resulting in evolutionary adaptations and changes) takes place in the Implicate Order. But to fully apprehend this we must start with some general remarks about te nature of the Implicate Order and its relation with the Explicate Order. Two views are possible here :
However, in a thoroughly deterministic setting of Reality -- which we consider the right setting -- forks do not fit. In the causal trajectories of Reality there are no points where Nature has two or more choices. Therefore, the 'harnessing' of the physico-chemical machinery must be described differently. For this we must return to chreodes as described earlier in the present document.
In fact the formal content of a given organic species, as that content resides in the form of a noëtic entity in the Implicate Order, is its chreode. When such a chreode is actually followed by physical and chemical processes in the Explicate Order -- that is, when these processes proceed according to that chreode, which proceeding is equivalent to the projection of the chreode from the Implicate Order into the Explicate Order -- an individual organism is developing from its egg-stage (initial condition) to the adult form. This morphogenetic process, including all post-embryonic generative processes, represents, the whole internal content of the given species, including not only all intrinsic morphological and physiological features together with their pre-stages and all other 'larval' features, but also all the capacities of the organism to behave in such and such a way, that is, all its abilities to present a behavior of a specific intrinsic type. And, as we know, this internal content of the given organic species is extended by external but still intrinsic contents : First of all the content of the actual ecological niche presently occupied by the individuals of the species, and then further also the contents of the several potential ecological niches of that same species. And it is noëtic reactions between the internal formal content and one of the corresponding external contents that will create new possibilities :
In the Implicate Order a given chreode can noëtically react either with the formal content of the actual ecological niche of the species noëtically represented by the chreode, resulting (after projection) in an intensification ( = more advanced specialization) of one or more already existing (in that organic species) adaptations to the species's present environment, or (reacts) with the formal content of one of the species's potential ecological niches, resulting (after projection) in the appearance of one or more new adaptations, that is, adaptations to a more or less new environment. Both possible results appear first of all as just noëtic reaction products in the Implicate Order, namely as new chreodes (alongside the old ones), that is, more or less altered chreodes. If then conditions in the Explicate order are appropriate at the relevant places and times with respect to one or the other new chreode this chreode will project into the Explicate Order. And this projection is then seen either as the evolutionary strenghening of an existing adaptation, or (in other cases, seen as) the formation of one or more new adaptations, that is, adaptations to a new environment, or better, to a new or extended ecological niche.
The decision to bring in chreodes into the present discussion (concerning evolution) is justified by the fact that organic evolution is really the more or less gradual alteration of the process of morphogenesis ( = individual development) in a given initial organic species in the course many years and of many generations. In this way a given evolutionary trajectory or lineage is a succession of generally a large number of gradually differing morphogeneses. Here an actually proceeding of the morphogenesis of an individual of a given species, as we see it in the Explicate Order, is the result of the projection from the Implicate Order into the Explicate Order of the corresponding chreode. This projection here consists in successive projections of the consequents of If-Then constants (as they play a part in the chreode) as a result of successive activation of groups of such constants.
The noëtic reaction of a chreode with the formal content of the actual or some potential ecological niche of the species represented by the chreode results, as has been said, in a new, that is, altered chreode. As a result of this the organic species is, in the one case, adapted still narrower to its original ecological niche (or to a specific aspect of it), or, in the other case, has acquired an adaptation to a new, formerly only potential, ecological niche. And a further result of this is that the horizon of possible ecological niches has widened, that is, the range of possible ecological niches or of aspects of them for the species to occupy has become somewhat larger. And now, after injection of this extended range of possibilities, the new chreode can again noëtically react with one of the formal contents in this injected range, resulting in yet another chreode. This process can, of course go on indefinitely, resulting in a consecutive series of chreodes, while the same series is in the Explicate Order expressed as a consecutive series of qualitatively different morphogeneses (plural!). And this series of morphogeneses (plural!), resulting in a corresponding series of succesive generations of organisms along a particular line of inheritance, constitutes a particular evolutionary trajectory. So now, finally, we have solved -- as far as this is possible anyway -- the problem of 'harnessing', 'overforming', or 'guiding', of the physico-chemical machinery at work in a given evolutionary process by non-physico-chemical agents. And we have done this by again involving the chreode (which we had earlier invoked with respect to the morphogenetic process, that is, the individual development) and showing its ability to change as a result of a noëtic reaction taking place in the Implicate order.
About the nature and general content of the noëtic reactions driving the evolutionary process.
What is the nature of the discussed "noëtic reactions" and their alleged "reaction products"? That is, what in fact are these 'processes'? Earlier we stated that they are somehow 'thought-like'. As a most general description of their nature this might be sufficient. But what about their general content? Are they something like statements, and if so, what precisely are they about?
As far as I know, little is to be found in the relevant literature. In the book The collapse of chaos, written by COHEN & STEWART, 1994, there are some hints, namely there where they describe the evolutionary process (-aspect) of "complicity". The latter is, however, only vaguely expounded and the discussion hopelessly confuses epistemological and ontological points of view (that is, on the one hand, how and whether we can know natural things and processes, and, on the other hand, what the nature of those things and processes themselves might actually be independently of their being known). Nevertheless the book powerfully stimulates further thought about the nature of organisms and their evolution, and indeed might even contain the seeds of our own theory of evolution-directed-by-non-physical-agents, that is, directed by noëtic reactions and their products, (the authors speak about interaction of "spaces-of-the-possible", which they identify with the phase-spaces of dynamical systems theory).
A far more worked-out discussion, which might be used (after re-interpretation) for elucidating and expanding our own theory, we have found in the Russian entomologist B. ROHDENDORF. He has written a lot about the systematics and evolution of insects including fossil insects (of which a lot have been found in the former Sovjet-Union -- an important site where mesozoic insects were found now lies in Kazachstan). The mentioned discussion we have found in ROHDENDORF's book Istoritsezkoje razwitije dwukrilich nasjekomich ( = The historical development of the two-winged insects), 1964, Moscow, written, of course, in Russian (which language I am able to read). There exists an English translation of this book : The historical development of Diptera, (The University of Alberta Press, 1974) but the translation is in some places poor or even wrong, especially in the purely theoretical parts of the book, and even whole paragraphs are forgotten to translate.
ROHDENDORF's general theory of the nature of the evolutionary process is undoubtedly rejected by western authors (of whom I know at least one) or his book is not even read by them. I have read the russian edition a long time ago (somewhere in the seventies when I was doing entomology) and have re-read it again these days (2007), now in the context of my, in the meantime acquired, knowledge of very general ideas and theories concerning the generation of form, especially (or at least geared to an understanding of) organic form (as has been laid down in all the five main Parts of the present website).
In his general theory, that is, only in his very general theory, concerning the basic process of organic evolution, ROHDENDORF proceeds more or less along the lines of the dialectical materialism of Marx and Engels (the philosophy of whom goes back to the German philosopher Hegel ). In the Sovjet-Union this was, of course customary, especially in stalinistic times (during which ROHDENDORF wrote many of his works). But we shouldn't be prejudiced against dialectical materialism just because it has been more or less forced onto scientific workers and has indeed resulted in some disasters (in the agricultural domain, for instance). As far as I have seen (by reading not only this particular book but also many other publications of ROHDENDORF ),
ROHDENDORF does not explicitly adhere to the marxistic theory (the very word 'marxistic', and also the names 'Marx', 'Engels' or 'Hegel', are nowhere mentioned in this, or any other book of him known to me), and does not engage in political statements or in unwarranted extrapolations (that is, going beyond mere biology). He has extracted some valuable and relevant elements from this marxistic theory, re-interpreted them biologically, and then has come up with his own version which is only about the evolution of animals, and certainly does not explicitly (and, as far as I could see, not even implicitly) include the evolution of human societies together with their political structures. And because I am opposed to the idea of the alleged exhaustive role (that is, excluding all other possible mechanisms) of the random-genetic-mutations-plus-natural-selecion mechanism in organic evolution, I am in principle open-minded with respect to any seriously-meant 'alternative' theory of evolution provided it does not flatly reject the general findings of DARWIN and his followers, that is, as far as it does not simply deny any role of natural selection and of genes in the process of evolution.
Well, there are a number of such 'alternative' theories of organic evolution (we already mentioned COHEN & STEWART ), and in ROHDENDORF we have found one that could provide valuable elements to our own ideas of organic evolution, which ideas were discussed above (in a still very general fashion).
So let us proceed along ROHDENDORF's lines.
When we examine any given (high-level) organic feature, such as a morphological, physiological or behavioral trait of some given organism insofar as the latter represents some systematic category (that is, some higher taxon, such as a genus, family, superfamily, suborder, etc.), and when we have succeeded to assess it functionally (that is, if we can assign a definite organic function to it), we see that it really is an adaptation to one or another particular aspect of its natural environment. And this means that such an adaptation in fact represents a solution of some previously existing biological problem or inherent conflict (that is, a real, but nevertheless in some sense formal, conflict or 'contradiction' in nature, not in the investigation). Indeed, some of such solutions have -- in the history of organisms -- given rise to whole new groups of organisms. These groups can be new species but also -- depending upon the nature of the adaptation -- new higher groups such as genera, families, suborders, etc. And because we can assume that every solution creates a new conflict, this conflict will, sooner or later, also be solved, creating yet again a new conflict. So according to this dialectical process evolution keeps on going. This is ROHDENDORF's position, and it is certainy worthwile to work it out further and see how it could fit into our own theory of the general process of evolution (a theory which invokes the concept of the Implicate Order as developed above).
The expression 'solution' inadvertently sounds as if we have to do with something 'thought-like', and maybe its nature (that is, the nature of biological solution) really is thought-like, or, as we would prefer to express ourselves, noëtic.
Although ROHDENDORF himself does not think of the mentioned solutions in terms of initially noëtic entities (eventually projecting from the noëtic domain of Being into the space-time world), we will 'locate' these solutions, that is, the initial 'invention' of them, within the Implicate Order. Only on subsequent projection into the Explicate Order they will be visible as morphological, physiological, or behavioral adaptations to particular aspects of the environment of the organism in question.
The nature of the internal [formal] conflicts
We will now paraphrase ROHDENDORF's exposition about the role of internal conflicts and how they drive throught their solutions (which depend on the nature of the environment in which the organism lives) the evolutionary process.
We will do so by following the Russian text (pp. 279) as close as we deem necessary and add comments when called for. By "paraphrase" I mean that although I am able to provide an exact translation, I decline to do so, because I want to have the freedom to adapt the given text where and when I deem necessary to in order to explain the content better or suit it to the needs of its integration into my own theory of evolution.
To what do the internal [formal] conflicts in the historical development of organisms amount? While considering phylogenesis [ = evolutionary development] as a historical process of modification of successive ontogeneses [plural] of organisms [ontogenesis is the complete process -- not only the embryological -- leading from (fertilized) egg-cell to fully mature organism] inseparably connected with their conditions of life, it is natural to see the essence of this process as the modifications (changes) of the qualities of the vital function [ = life processes of an organism]. The realization of organic needs of the animal organism -- processes of exchange [feeding and respiration], of individual development, of multiplication, and of [geographical] distribution (spread), which proceed under the control of the nervous apparatus [and of the organism's genes], all this turns out to the basis, the necessary condition for the very existence of the living being. Only in virtue of the presence of these processes the prolonged existence of organic forms and their historical development is possible. Therefore the necessary conclusion is that the internal conflict of [that is, between] the needs [demands] coming from the [internal] life processes represents just the basis which drives the process of [historical] development, and which turns out to be the true way by which the changes of the [particular] living conditions generate the origination of new forms [that is, the prevailing living condition (environment) determines the nature of the solution of some particular internal [formal] conflict].
What is the essence of the internal conflict of the life processes? An extensive and meticulous analysis of the dialectic essence of the primary life processes as natural phenomena is not the objective of the present investigation, which will be an extensive totally independent theme all by itself. My [that is, Rohdendorf's] task just consists of the g e n e r a l d e t e r m i n a t i o n of the character of the conflicts of the main processes of the vital function and the concrete consideration of a number of single cases of historical development. As can be seen easily, every life process, every [organic] phenomenon, has its features belonging only to it, and, essentially, the characters of the conflicts in each individual case are totally idiosyncratic and not repeatable. This extreme diversity [resulting from the corresponding diversity of internal conflicts] of the pathways of historical development of different groups clearly indicates the importance of particularly the g e n e r a l d e t e r m i n a t i o n s of the characters of the internal conflicts in the life processes. Every detailed and exhaustive investigation necessarily forces one to delve into the analysis of a few partial processes and does not make possible to encompass the whole problem with the necessary width. The above mentioned general determination of the characters of internal [formal] conflicts in the life processes of the organism boils down to the establisment, more precisely said the clarification, of those needs [demands] [of those life processes] which are 'submitted' by the organism to the living conditions for the realization of the necessary processes. The diversity of these demands clearly indicates and clarifies their mutual contradiction [that is, just because these demands can come in so many different forms, conflicts between them will necessarily arise].
The internal conflict in the organism is a very complex system of mutual relations of processes. All these most different [formal] conflicts can be divided into two large groups, namely (1) properly internal conflicts of the organism, that is, the conflict between two different aspects [each of them is another life process, such as feeding] of the general life process as a whole, and (2) internal conflicts prevailing between two different aspects of a particular single life process [such as, for example, feeding].
The first group of conflicts -- comprising conflicts between demands [needs] of different aspects of the one general life process [vital function] are internal with respect to the organism and external with respect to the given aspects of the general vital function. Examples of such conflicts are presented in the next table, that is, in Table I :
Table I
| Terms of [ formal ] conflict | |
| Need for respiration ( in an aquatic or terrestrial environment ) | Need for nourishment ( on land or in the water ) |
| Need for nourishment ( on in water or on land ) | Need for individual development ( in the water or on land ) |
| Need for reproduction | Need for individual development and growth |
| Need for nourishment in definite conditions | Need for reproduction in different conditions |
These examples may easily be multiplied : It is sufficient to consider any concrete instance of the living conditions of animals for the conflicting needs of the different aspects of the [general] vital function to become clear, not to speak of the fact that each example can be illustrated in different forms by the material of different animals.
It is possible that some obscurity may appear concerning the very essence of the representations about the contradictoriness of the processes indicated in this table. While the identification of conflicts between processes connected with different demands imposed to the [external] conditions, for instance between respiration, nourishment, development and reproduction, is not particular difficult, it is a bit more difficult to identify conflicts between reproduction and individual development, but also in this case the depth of the difference of these, each other conditioning, processes directly points to their contradictoriness. Considering, for instance, the conflicts between, on the one hand, the features of the processes of exchange (nourishment, respiration) and [the features of] the processes of individual development on the other, we can easily see them in the diffences of the processes themselves. Indeed, the realization of exchange during the time of development is continuously modified as a result of growth (enlargement) and as a result of changes (that is, of rebuildings) of the organism. The very fact of these differences in the just given example proves the presence of [formal] conflicts.
Then we have the second group of conflicts which latter boil down to the conflicting features of (each of) the particular processes (these latter) together making up the one overall vital process of the organism, that is, their internal [formal] conflicts [So each single process -- as part of the overall vital process of the organism -- has (at least) two (formally) conflicting features]. Examples are presented in the next table, that is, in Table II :
Table II
| Process of vital function | Terms of [formal] conflict | |
| Exchange as a whole | Assimilation -- integration of necessary substances and the building up of living material | Dissimilation -- excretion of waste substances and the decomposition of living material |
| Nutrition | Need for nourishment of the most caloric substances | Need for obtaining water by the organism |
| Nutrition | Need for rapid and abundant absorption of nutritive substance | Need for complete assimilation of food |
| Nutrition | Need for feeding on easily accesssible substances | Need to secure chemical full-valueness of nutritive material |
| Respiration | Need for the greatest intensification of the repiratory process by means of extraction of the maximum mass of oxygen | Need for the most rapid delivery to the tissues of the body of oxygen and removal of carbon dioxide |
| Individual development | Need for most rapid attainment of the fully mature condition | Need for growth and rebuilding of the organism in development |
| Individual development | Need for building up of larval organs and other temporary structures | Need for reduction of temporary structures and building up of organs of the definitive state |
| Individual development | Need for quick growth and development of the earliest minute stages in ontogenesis | Need for intensive feeding and protection of these stages |
| Reproduction | Need for the formation of sex products in the greatest quantity | Need for the formation of the most viable sex products |
| Reproduction | Need for obtaining fertilization of a possible large number of egg-cells | Need for guaranteeing the development of the greatest number of egg-cells |
These examples of the internal [formal] conflicts between individual features of one or another single life process can also easily be multiplied when considering concrete phenomena of historical development. There is no need to analyze more precisely the essence of the contradictoriness of one or another partial process of the general life process. In the table are, with sufficient clarity, indicated the needs which form conflicting sides which together make up an unbreakable unity [for instance Assimilation and Dissimilation as conflicting sides of the process of Exchange, but nevertheless together forming a unity].
[As with the conflicts of the first group] the solution of the conflicts and the substitution of them by new ones realize the process of organic development.
In concluding the discussion of the internal [formal] conflicts of the organism, it is absolutely necessary categorically to underline, t h a t o n l y t h e s e [formal] c o n f l i c t s r e p r e s e n t t h e r e a l f o u n d a t i o n o f [historical] d e v e l o p m e n t. Evaluation of every kind of so-called 'conflicts' between predator and prey, between organism and the conditions of its habitat, between those and those competing organisms, indicates that such mutual relations a r e n o t t h e i m m e d i a t e r e a s o n [cause] o f t h e m o d i f i c a t i o n s o f o r g a n i s m s, t h e y a r e j u s t c o n d i t i o n s w h i c h d e t e r m i n e d o n e o r a n o t h e r s o l u t i o n o f i n t e r n a l c o n f l i c t s. Analysis [in the sense of distinguishing] of a correct understanding of internal conflicts and the influence of external factors on the organism represents an important methodological problem, the elaboration of which turns out to be an independent and rather broad task, upon which I cannot engage.
It is paramount to realize that a denial of the significance of the internal conflicts in the organism and then attribute to the external factors the import of the i m m e d i a t e driving forces of organic development shows itself as the well-known form of mechanicism, which, under the name of the "struggle for materialism" in fact replaces dialectics [ I assume that the reason for this replacement is the denial of the internal nature of the conflicts, that is, their fundamental nature].
REMARK ( Intermezzo of the writer of the present website).
Indeed, a formal conflict cannot as such reside in any viable and fully functioning individual organism. Such an 'organism' would not be able to exist, neither could its formal content as such, that is, as a contradiction, be present in the noëtic domain. Nevertheless, the formal conflicts as discussed here are inherent in the formal essence of Life as such, meaning that they are present in the Implicate Order. However, they were not present there before Life had emerged in the Explicate Order. The emergence of Life in the Explicate Order has, through injections, enriched the Implicate Order. And as Life progressed and diversified new forms of formal biological conflicts were added to the noëtic content of the Implicate Order. But such a conflict cannot be a single noëtic entity, because then it would be a contradiction in terms which as such cannot exist anywhere. Indeed, each term of such a conflict is connected, not with the other term of the conflict, but with the formal contents of corresponding organic species (not necessarily insects). Indeed, each single term of a given conflict is a specific need of some type of organism, while each corresponding opposite term is a specific need of some other type of organism. So the conflict's contradictory content being divided and as such dispersed across the contents of different organic species prevents the mutual annihilation of its opposite terms, and so prevents the annihilation of the formal conflict itself. So when, in the Explicate Order, organisms of a given species have moved into a different but potentially appropriate environment (or when the original environment has changed into one of the species's potential environments), the formal content of that species (as it is injected into the Implicate Order) and the formal content of that new environment (also as it is injected in the Implicate Order) will noëtically react with each other. "Noëtically react" here means that some relevant formal biological conflict (residing in the Implicate Order in the way just expounded) becomes the (noëtic) c o n t e x t of the reaction. The conflict is then (noëtically) resolved and the nature of the (formal) solution (i.e. one solution or another) is determined by the formal content of one of the noëtic reactants, namely the formal content of the mentioned new environment (the one or the other). The in-this-way-generated formal solution of the conflict is then projected into the Explicate Order at the appropriate places and times. And what we then actually observe, or deduce from observations, is the evolutionary development, having taken place in all or in a number of individuals of a given organic species, of a certain adaptation, or set of adaptations, to certain aspects of the new environment.
We can agree with ROHDENDORF that the 'conflicts' as we think to see them in the relationships between predator and prey, organism and the conditions of its habitat, etc., are, it is true, conflicts, but, he rightly holds, only in a derived way : They refer back to the real conflicts, that is, the fundamental conflicts which lie at the bottom of them and which all lie inside the organisms (as shown above with respect to the organism's life processes).
However, here, the expression "inside the organisms" should be understood not as "inside the organism" (singular), that is, "inside the body" in the sense of "inside the body's physiology", but (should be understood as) "inside Life as such". Any such biological conflict is inherent in the very fabric of Life-as-a-whole, but in such a way that its opposing terms are each connected with different organisms. For further elaboration of this issue, see below.
These -- real -- primary conflicts (such as between aspects of the main life processes of the organism) thus lie at the foundation of the secondary -- we could say high-level -- conflicts (as we see them between predator and prey, etc.).
In terms of the integration of ROHDENDORF's theory of internal conflicts and their solutions into our own evolutionary theory which invokes the Implicate Order and thus noëtic reactions and noëtic reaction products, we would like to emphasize the following :
The noëtic reaction is not between the two members of the internal [formal] conflict, but between the formal content of the organismic species in question and the formal content of the given (potentially suitable) environment in which individuals of the species find themselves. This particular environment determines the nature of the solution of the internal conflict. If those organisms had found themselves in another (also potentially suitable) environment, then another solution of the internal conflict would have been realized. A genuine internal conflict only has a formal nature, that is to say that it is only going to play a role when the mentioned noëtic reaction takes place (in the Implicate Order).
The many examples and discussions yet to come will increasingly clarify the matter still further and continually help to test and to elaborate further our theory.
(End of Remark)
(continuing paraphrasing ROHDENDORF's text)
The pathways of the historical development of two-winged insects [Diptera] discussed in the main part of the book indicated the exclusive import of a dialectical analysis of the processes of development as an unbroken chain of conflicting processes, their solution, and their [consequent] replacement by new conflicts. [In the theoretical part of the book] a further exposition of the features of the internal conflicts between the needs of the organism described the character of those very conflicts and their terms [sides] and provided a conception concerning the general traits of the conflicts in organic development.
The next necessary stage of the whole concluding part of the investigation [that is, of the book] boils down to a meticulous description of concrete conflicts in the historical development of one or another animal group [this latter accounts for the mentioned concreteness of examples to come], first of all, of course, the two-winged insects, which are most familiar to the author.
Essentially, the examples, that will be given below, of concrete conflicts turned out to represent concise independent sketches of individual episodes from the history of the given phyla or tribes of animals. In these examples, set up according to a definite plan, first of all is indicated the character of the conflict -- this or another aspect of the vital function upon which [aspect] the given process had seized. Further the terms of the conflict were analyzed -- those or other needs of the vital function between which [needs] originated conflicting relations. Finally, for every example solutions to the conflict are indicated, which [solutions] were realized in these or those [prevailing] [external] conditions of life. And with all this the concrete animal groups were indicated which had originated as a result of the given solution of the conflict.
As will become clear in the examples, special significance is attributed to the fate of one of the two terms of each conflict, namely that term which along the course of the solution of the conflict turns out to be the lesser term, which will undergo modification as a result of the 'victory in the struggle' of the conflicts [each conflict representing such a struggle] of the other term, namely the chief term of the [given] conflict. In essence, the changes [taking place] in the lesser term also turned out to present themselves as those characteristic traits, which characterize a new group [of animals] which emerged as a result of the given concrete solution of the conflict. Further down, after the discussion of the concrete examples, we shall more precisely deal with these relationships between terms.
EXAMPLE 1.
| Terms of [ formal ] conflict | |
| A. Function of Locomotion. |
B. Function of Feeding. |
| Need for a solid substrate for locomotion of the larva provided by three pairs of walking legs. | Need for nutrition of [and in] a moist semiliquid substrate forming comparatively large masses. |
[ As has been said, the nature of the solution of the conflict -- i.e. which one of the possible solutions will actually be materialized -- depends on the nature of the external conditions, that is, on the nature (climate, biotic composition, etc.) of the environment in which the given individual organisms, representing some biological taxon, find themselves.]
Possible set of external conditions.
In conditions of low degree of accessibility [short supply, or just non-appropriateness] of surface layers of the nutritive substrate ( [ this non-appropriateness being so by reason of ] desiccation of the substrate by the action of dry air, or [by reason of] poor nutritional value of the [surface layer of] substrate, and, finally, [by reason of] of the necessity of cover [of the larva] ), the larvae [evolutionarily] developed the ability to live submerged in the deep mass of the substrate and [so] reaching moist semiliquid layers [of it]. [In these external conditions] the chief term of the conflict turns out to be B, while term A became modified and is the lesser term. The larvae lost their walking legs and acquired a worm-like body. This turned out to be the chief prerequisite to the formation of the first representatives of the order of two-winged insects.
Alternative set of external conditions.
In different [external] conditions (moist environment, the absence of the necessity to have cover in the deep mass of substrate), the larvae fed themselves with surface layers of the substrate : here the chief term turns out to be A. The larvae retained walking legs and [evolutionarily] developed the ability to eat relatively solid dry substrates. This solution did not lead to the emergence of apodous larvae. As a result there arose the 'sister group' to the Diptera, [namely] not yet known in more detail representatives of the mecopteroid insects [that is, scorpionfly-like common ancestors of the orders Lepidoptera (butterflies), Panorpida (scorpionflies), Trichoptera (caddisflies), and Diptera (two-winged insects) ].
[ In these alternative external conditions -- moist environment, absence of the necessity for the larva to have cover -- the surface layer of the nutritive substrate, is, it is true, dry, but not too dry, and therefore not degraded in nutritive quality. So there is no need for the larva to be adapted to feed on semiliquid material, that is, the lesser term is now B and it therefore changes.
[ For reasons of clarity we must add that in different places and times on the earth's surface the external conditions are normally different, so both solutions can, generally, be realized even with respect to the same initial animal species. They are realized then in the corresponding different places and times.]
EXAMPLE 2.
| Terms of [ formal ] conflict | |
| A. Function of Individual Development. |
B. Function of Locomotion. |
| Need for visiting [by the adult] locally disposed substrates to deliver offspring, that is, substrates in which the larvae [should] live. | Possibility to retain the performance of [just] slow flight [and that means : no need to invest in developing strong flight muscles and special wings] by using two [functionally more or less independent] pairs of wings possessing insignificant lifting powers. |
Possible set of external conditions.
Conditions of low degree of appropriateness of surface layers of the nutritive substrate ( [ as a result of] its desiccation by the action of dry air, or [as a result of] poor nutritional value of the substrate, and, finally, [as a result of] of the necessity of cover [of the larva] ) determined only the one solution consisting in the development of strong flight by means of the anterior pair of wings [the hind wings have changed into, as it seems, gyroscopic organs, the halteres], that is, the development of the two-winged condition. A turned out to be the chief term, and the new flight capability was developed, that is, B changed, which is therefore the lesser term. The formation of two-winged insects was the result of this solution.
An alternative solution [triggered by alternative external conditions] of the conflict did not materialize in history, as would then B be the chief term and the insects would have retained feeble flight by means of the ancestral four wings, and then finding of [only] locally disposed nutritive substrates for the larvae would have been difficult and such insects necessarily would have died out (that may possibly have happened in the history of the group, but we do not have any paleontological evidence concerning it).
EXAMPLE 3.
| Terms of [ formal ] conflict | |
| A. Function of Individual Development. |
B. Function of Exchange. |
| Need for feeding by means of accessing the most moistened layers of the nutritive substrate of the larva. | The need for respiration by means of contact with atmospheric air of all spiracles of the [body] segments. |
Possible set of external conditions.
In conditions of increased moisture (for instance free water) A turned out to be the chief term and the larvae developed the ability to live in the deep and dampest parts of the substrate. This determined the modification of the lesser term of the conflict -- the ability of having the tracheal system in contact with the atmospheric air. There had been developed metapneustic respiration by means of the enlarged and complicated most posterior pair of spiracles, while the other spiracles atrophied. As a result of such a solution of the conflict the first representatives of the ancient Tipulomorphs appeared.
[ The infraorder Tipulomorpha contains Diptera such as crane flies (daddy-longlegs and the like), blood-sucking mosquitos, non-biting midges, blood-sucking midges, blackflies, and others.]
For the next exchange of conflicts see Example 4.
Alternative set of external conditions.
In conditions of moderate moistness (for instance in terrestrial biotops) B turned out to be the chief term, namely that the larva of the two-winged insect retained respiration with all segmental spiracles. This determined the modification of the lesser term of the conflict, namely the need for feeding of and access to the most moistened parts of the substrate. A change of food [that is, the adaptation to different nutritive materials] was [evolutionarily] worked out, [that is,] its improvement by passing over to more nourishing substrates [but] significantly more rare and [only] locally disposed but [as had been said] by contrast very rich in nutritive substances (such as rotting wood which is infested by mycelia, bark of trees, fruiting bodies of the higher fungi). As a result of this solution of the conflict the first representatives of the ancient Bibionomorphs arose.
[ The infraorder Bibionomorpha contains Diptera such as fungus gnats, ground midges, gall midges, and others.]
For the next exchange of conflicts see Example 5.
The Implicate Order.
When they, that is, the larvae, venture into an environment with increased moisture (or when their original more or less dry environment becomes wetter), the formal content of the species (as it is injected into the Implicate Order) will noëtically react with the formal content (also as it is injected into the Implicate Order) of the new environment (that is, the environment with increased moisture), and the appropriate c o n t e x t of this reaction will be the conflict between the terms A and B. The outcome of the reaction is a solution of the formal conflict and will consist in the metapneustic condition of the tracheal system. In fact it will consist in a new formal content of the species. Upon projection of this new formal content into the Explicate Order we observe the evolutionary development of metapneustic larvae that now inhabit the mentioned moist environment. And these insects are the first representatives of the ancient Tipulomorphs.
When, on the other hand, these insects, that is, their larvae, venture into an environment that is only of moderate moistness, that is, relatively dry, the larvae will retain the original condition of their tracheal system, that is, they remain holopneustic and thus still having all the spiracles of their segments in contact with atmospheric air. Now the formal content of the species noëtically reacts with the formal content of the relatively dry environment, and the appropriate c o n t e x t of this noëtic reaction is again the conflict between the terms A and B. But now the outcome of the reaction is a different solution (different from the one above) of the conflict : The appropriate food material, that is, the food material that is now regularly chosen to feed on, has changed, it has higher nutritive value than the original regular food of the larvae. In fact this solution consists of a new formal content of the species. Upon projection of this new formal content into the Explicate Order we observe the evolutionary development of a different feeding habit of the larvae : from formerly feeding on material that was relatively poor with repect to its nutritive content but abundantly present to material very rich in nourishment but significantly more rare and only locally disposed, such as rotting wood and the like. And these insects are the first representatives of the ancient Bibionomorphs.
It is now time to describe this Example 3 in terms of the Implicate Order, and thus in terms of noëtic reactions, as was explained in the second half of the above Remark. Say we have to do with a population of a particular species of primitive and ancient Diptera. The larvae of them breathe by means of having in contact with the atmospheric air all the spiracles of the segments, which we can call the holopneustic condition of the tracheal system.
EXAMPLE 4.
| Terms of [ formal ] conflict | |
| A. Function of Exchange. |
B. Function of Individual Development. |
| Need for retaining contact with atmospheric air during respiration of the larva. | The need for feeding of the larva in and of muddy or detritus masses at the bottom of a reservoir. |
Possible set of external conditions.
In the conditions of slowly flowing small [that is, shallow] reservoirs A turned out to be the chief term [of the conflict], that is, the larvae realized contact with the surface film for respiration. Feeding on muddy masses at the bottom turned out to represent the lesser term [ B ] [of the conflict] which [subsequently] changed : predation was developed, which here means the ability to feed on micro-organisms of the surface film of the water, or of the surface of [aquatic] plants or rocks. As a result of such a solution of the conflict there originated -- through a chain of numerous secondary conflicts -- different groups of Tipulomorphs, namely crane flies (Tipulidea), blood-sucking mosquitos (Culicidea), dixideans (Dixidea).
Alternative set of external conditions.
In conditions of rapid flow, of moving water, and of the difficulty in preserving contact with the surface film for respiration, B turned out to be the chief term of the conflict -- the larvae developed feeding on muddy masses at the bottom. Respiration of atmospheric air and direct contact with the surface film of the water turned out to be the lesser term [ A ]. The larvae obtained the ability of osmotic respiration of oxygen dissolved in the water and were [consequently] able to leave the surface film and populate the reservoir all the way down to its bottom. The tracheal system of these larvae became closed. The origin of a large group of the Tipulomorphs was the result of this solution of the conflict, namely the Chironomidea to which belong the non-biting midges (Chironomidae), the blood-sucking midges (Ceratopogonidae), and the blackfies (Simuliidae).
Exposition of this Example in terms of the Implicate Order.
But if the external conditions were to consist of rapid flowing water causing the preservation of contact (of the larva) with the surface film for respiration difficult, a different solution of the conflict was determined. The formal content of the species now reacts noëtically with the formal content of the just described external conditions, and the noëtic c o n t e x t of this reaction is again the conflict between the terms A and B of the present Example. What we see upon projection of this solution into the Explicate Order is the evolutionary transformation of the original metapneustic larvae into larvae of which the tracheal system is closed. These larvae now osmotically breathe the oxygen that is dissoved in the water and can now populate much deeper layers of the reservoir all the way down to its bottom. What we then see is the origin of the very large Tipulomorph group of Chironomidea, which consists of non-biting midges (Chironomidae), blood-sucking midges (Ceratopogonidae), and blackflies (Simuliidae).
Here we have to start with the ancient Tipulomorphs, which were the result of a solution of the previous conflict, namely the solution determined by external conditions of increased moisture (for example free water) which led to metapneustic larvae, that is, larvae that breathe by means of the enlarged and complicated pair of the most posterior spiracles, while the other spiracles had been atrophied.
These larvae now enter, or just happen to be in, external conditions of slowly flowing small water reservoirs. In the Implicate Order we now will have a noëtic reaction between (1) the formal content of the given original species (some ancient Tipulomorph) and (2) the formal content of the described environment (slow-flowing small reservoirs), and the noëtic c o n t e x t of this reaction is the formal conflict consisting of the terms A and B of the present Example (Example 4). The solution of the conflict, determined by the mentioned external conditions, is a modified formal content of the given (original) species. What we see upon projection of this new formal content into the Explicate order is the evolution of predative behavior of the larvae, that is (here) the ability to feed on micro-organisms at the surface film of the water, at the surface of aquatic plants and of submerged rocks. What we then see (or conclude from indirect observations) -- after an additional chain of secondary conflicts and their solutions had followed upon this -- is the evolutionary development of several groups of Tipulomorphs, namely crane flies (Tipulidea), blood-sucking mosquitos and also some non-bloodsucking species (both taken together as Culicidea) (about the evolutionary branching of this latter group into Culicidae (genuine mosquitos) and Chaoboridae, see Example 7), and the Dixidea.
EXAMPLE 5.
| Terms of [ formal ] conflict | |
| A. Function of Individual Development. |
B. Function of Locomotion. |
| Need for the nourishment of the larva of [only] locally disposed substrates, that is, fruit bodies of higher fungi and rotting wood which is infested by mycelia. | Demand for retention of weak flight ability and of imperfect running powers. |
Possible set of external conditions.
In conditions of a cold climate with a sharp alternation of seasons, in coniferous forests, A turned out to be the chief term, that is, feeding on local temporary substrates. The lesser term, that is, the ability of locomotion, was modified. Strong running organs, 'muscular' legs, were developed which permit the insects to dig into the litter of woods, and also the organs of flight were improved (powerful muscular system and perfected wings). This solution of the conflict turned out to be the fundamental prerequisite of the formation of the family of fungus gnats (Fungivoridae).
Alternative set of external conditions.
In the conditions of a moderate climate and deciduous forests B turned out to be the chief term, that is, the demand for the retention of weak powers of locomotion. The lesser term A was modified : feeding on dead leaves of plants as a result of perfecting the digestive system of the larvae which acquired the ability of digesting cellulose, which turned out to be a significantly advantageous acquisition and which had strongly broadened the food resources of the insect. Such a solution of the conflict determined the origin of ground midges (Sciaridae), the most important of the [co-]makers of humus.
Exposition of this Example in terms of the Implicate Order.
Alternative external conditions.
The ecological type of Diptera to start with here is the one that originated as a result of the second possible solution of the conflict of Example 3. The larva of this type of Diptera had preserved the ability to breathe with all segmental spiracles. The chosen food material of the larvae of these insects possessed a high nutritional value but was only locally disposed : rotting wood infested with mycelia, the bark of trees, fruiting bodies of higher fungi. Our type of Diptera was one of the first representatives of the ancient Bibionomorphs.
Now, this type of Diptera enters into (or happens to find itself in) the conditions of a cold climate and sharp alternations of seasons in coniferous forests (that is to say, the insects have either moved into such conditions, or the original conditions in which they lived had changed). This did not significantly change the presence of the type of food required by our Diptera as described just above, it only intensifies the local nature of the resources, now also in time (because in the cold winters they are not expected to be in their required state in order to serve as food). However, and naturally, these conditions stimulated the development of stronger and better organs of locomotion of the adult insects to deliver their eggs at the right sites, that is, in order to find the (scattered) nutritive substrates for their larvae.
Now in the Implicate Order a noëtic reaction will take place between (1) the formal content of the original species of Diptera and (2) the formal content of the just described environment. And the noëtic c o n t e x t of this reaction is the formal conflict between the terms A and B of the present Example, i.e. Example 5.
Upon projection of the reaction product, that is, the solution of the formal conflict, into the Explicate Order we see (or deduce from indirect observations) the evolutionary development of certain dipterous insects going its way from original primitive members of the infraorder Bibionomorpha to more progressed members of it, namely insects possessing in their adult stage strenghened organs of locomotion, that is, we witness the origin of the fungus gnats (Fungivoridae).
If, on the other hand, our original dipterous insects (primitive Bibionomorpha) find themselves in a moderate climate in deciduous forests, they can in principle find appropriate food almost everywhere and so no material and energetic resourses need to be spended for developing stronger locomotive organs. So the solution of the conflict will in this case, that is under the just mentiond external conditions, be different. In the Implicate Order a noëtic reaction will occur between (1) the formal content of the species of our primitive Bibionomorph and (2) the formal content of the just described alternative external conditions. Again the noëtic c o n t e x t of this reaction will be the formal conflict between the terms A and B. of the present Example. Upon projection of this solution into the Explicate Order we see (or deduce from indirect observations) the evolutionary origin of dipterous insects whose larvae eat dead leaves of plants, which are in abundant supply in the deciduous forests in the areas in the zone of moderate climates. That is, we see the evolution of the digestive apparatus of the larvae leading to the ability of digesting cellulose, and, accordingly, we see the evolutionary appearance of the ground midges (Sciaridae).
EXAMPLE 6.
| Terms of [ formal ] conflict | |
| A. Function of Exchange [Feeding]. |
B. Function of Individual Development. |
| Needs for the nourishment of the larva on tissues of rotting leaves of vascular plants on the surface of the ground. | Need for stable dampness prevailing there where the stages of development live, namely eggs, larvae, and pupae. |
Possible set of external conditions.
In conditions of decreased moisture of plant communities in a moderate, perhaps, subtropical climate, and of rapid desiccation of plant residues, B turned out to be the chief term, the acquisition of stable conditions for [individual] development. The lesser term, A, changed : the ability [of the larva] to live in [and feed on] the tissues of l i v i n g plants was developed. This most important perfection was realized as a result of a change in digestion of food (the development of extra-intestinal digestion) and [was (also) realized as a result of a change of ] the methods of oviposition, and turned out to be the prerequisite of the formation of an extensive group of insects, namely the gall midges (Cecidomyiidae).
ROHDENDORF did not put forward a set of possible alternative external conditions (determining a different solution to the internal conflict).
We might indicate that set of possible alternative external conditions as the one we described in the second half of the previous Example, that is, Example 5 :
Alternative set of external conditions.
In the conditions of a moderate climate and deciduous forests A turned out to be the chief term, that is, the needs for feeding of the larvae on tissues of rotting leaves of vascular plants on the surface of the ground (the forest floor). The lesser term B was modified : The demand for a stable degree of moisture was given up in favor of feeding on dead leaves of plants as a result of perfecting the digestive system of the larvae which acquired the ability of digesting cellulose, which turned out to be a significantly advantageous acquisition and which had strongly broadened the food resources of the insect. Such a solution of the conflict determined the origin of ground midges (Sciaridae), the most important of the [co-]makers of humus. Indeed, this solution co-determined the formation of this group of insects (together with the working-out of a solution of another internal conflict, see Example 5).
EXAMPLE 7.
| Terms of [ formal ] conflict | |
| A. Function of Feeding. |
B. Function of Respiration. |
| Need for the nourishment of all stages of individual development -- larva and winged insect. | Need for respiration of atmospheric air during individual development. |
Possible set of external conditions.
In the conditions of the habitat of the larvae, namely small stagnant [water] reservoirs, poor in oxygen, rich in micro-organisms, in a moderate or warm climate, B turned out to be the chief term [of the conflict], namely the connection with the surface film [of the water] in order to be able to breathe atmospheric air. The lesser term, A, -- feeding -- was modified : the larvae developed microphagia [instead of other ways of feeding], that is feeding on micro-organisms, and the winged insects developed blood-sucking on vertebrate animals [instead of other ways of feeding]. This solution of the conflict turned out to be basis of the origin of blood-sucking mosquitos (Culicidae).
Alternative possible set of external conditions.
In conditions of large reservoirs like lakes, or flowing cold reservoirs, rich in oxygen and plankton, A turned out to be the chief term [of the conflict], namely the completeness of feeding. The lesser term -- the respiration of the larva -- changed : the larvae became active predators and they acquired the ability of osmotic respiration. They [as a result] left the surface film and populated the whole volume of water. The winged insects started to reduce their ability to feed, up to complete aphagia [that is, to not taking any food whatsoever, which is a form of beginning desimaginazation ( = the reduction, and eventually complete loss, of the adult state) ]. This solution of the conflict turned out, through a chain of intermediary conflicts, to be the base of the origin of the group called Chaoboridae [non-biting mosquito-like diptera].
Because -- referring to the alternative set of external conditions -- the water is rich in oxygen, it will be rich in plankton. And plankton consists principally of small aquatic organisms larger than true micro-organisms. Therefore plankton is a food source richer in required nutritive substances than are micro-organisms. So when the larva feeds on plankton the adult insect can dispose of feeding.
EXAMPLE 8.
| Terms of [ formal ] conflict | |
| A. Function of Locomotion. |
B. Function of Shelter. |
| Need for [the ability of] fast locomotion, that is, of flight or running, for securing food, spread, and reproduction. | Need for protection against predators and against [adverse] abiotic factors (rain). |
We had to restructure ROHDENDORF's exposition of the solutions of this particular internal conflict, because in his version things seem to be accidentally mixed up.
Possible set of external conditions.
In conditions of open landscape and vast spaces A -- retention or development of strong flight powers and/or running powers -- turned out to be the chief term, while B -- need for protection -- turned out to be the lesser term. If we assume that the first winged insects did not have the ability to fold their wings along the body during periods of rest, then conditions of open terrain would favor the retention of the original large wings remaining spread-out during rest (like we still see in dragonflies) allowing the insect quickly to resume flight when necessary. This solution of the conflict -- determined by the prevailing external conditions -- was a prerequisite for the formation of the Paleoptera.
Alternative set of external conditions.
In conditions of dense vegetation and only narrow spaces B turned out to be the chief term, while A turns out to be the lesser term and changed : the originally paleopterous insects gave up on their ability of fast powerful flight, because their long spread-out wings would make it impossible for them to inhabit places with a dense plant cover. Therefore they developed the neopterous condition, that is they developed shorter wings and developed the ability to fold them along their body when not flying. And certainly, having their wings thus folded, it was then possible for them to live in such habitats and be relatively hidden from airborne predators.
Instead of assuming such a situation -- that is the supposed fact that certain ancient insects came to find themselves in conditions of dense vegetation -- we could surmise that the appearance of dense vegetation was an opportunity for some of the existing insects to enjoy an alternative form of protection against airborne predators : instead of strong flight powers allowing the insect to escape from these predators they evolutionarily shortened their wings and folded them along their body in order to be able to hide themselves in the dense vegetation. The neopterous condition is present in many recent insects such as grasshoppers and locusts, beetles, flies, bees, wasps, while the paleopterous condition can still be found in recent insects such as dragonflies and mayflies.
EXAMPLE 9.
| Terms of [ formal ] conflict | |
| A. Function of Individual Development. |
B. Function of Reproduction. |
| Need for the most rapid individual development, that is, the most rapid attainment of the fully mature condition. | Need for the greatest reproduction, that is, for the maximal amount of offspring. |
Possible set of external conditions.
In conditions of warm waters and [consequently] harbouring a moderate amount of plankton, and a great number of predators, A turned out to be the chief term -- the most rapid attainment of the fully mature condition. The lesser term, B, -- maximal amount of offspring -- changed : the animals developed viviparity, thereby [that is, as a consequence] strongly decreasing the amount of offspring from [any]one individual but, on the other hand, the most exposed period of individual development was, as a result, much shortened. This kind of solution of the conflict was realized in the history of many groups, for example in sharks (Elasmobranchii).
Alternative set of external conditions.
In conditions of moderate or cold waters [consequently] rich in plankton, and relatively poor in predators, B turned out to be the chief term -- maximal amount of offspring -- while A was the lesser term which changed : The speed of individual development was altered. An extreme high degree of fertility developed and during individual development special larval forms appeared, and the length of the developmental period had increased. Such a kind of solution of the conflict was realized in the history of very many groups of fish-like organisms for example in herrings (Clupeiformes).
EXAMPLE 10.
| Terms of [ formal ] conflict | |
| A. Function of Locomotion. |
B. Function of Exchange. |
| Need for fast locomotion for the recovery of food and protection from enemies. | Need for protection of the body from overheating. |
Possible set of external conditions.
In conditions of rich vegetation A turned out to be the chief term -- the need for fast locomotion [and protection from enemies]. The lesser term, B, that is, the protection against heat, changed : The animals developed the ability to rise on their posterior limbs to realize a better overview of the region [and they did not develop structures to protect them from overheating because in terrains of dense vegetation such structures are not needed]. This production of a bipedal condition turned out to be the most important pathway of historical development, which became realized in many groups of reptiles.
Alternative set of external conditions.
In conditions of exposed terrain B turned out to be the chief term -- the need for protection from overheating, which was realized by the development of a thick bony or skin armour. A, the lesser term, changed : The animals did not develop the ability of fast locomotion, they became relatively slow moving. This way of solving the conflict determined the origin of the group of armoured archosaurs, namely the ankylosaurs.
EXAMPLE 11.
| Terms of [ formal ] conflict | |
| A. Function of Respiration. |
B. Function of Feeding. |
| Need for continuous respiratory movements for exchange of air in the lungs. | Need for prolonged presence of food in the mouth cavity for its grinding and swallowing. |
Possible set of external conditions.
In the conditions of a warm climate and the necessity of grinding the food before swallowing, the term A became the chief term -- continuous respiration which is necessary for an intensive exchange in conditions of high temperatures. The lesser term, B, -- holding food in the mouth -- changed. The animal developed a special adaptation which allows uninterrupted respiration when food is present in the mouth -- the secondary palate was developed. Such a solution of the conflict turned out to be the most important prerequisite for the origin of the first mammals.
EXAMPLE 12.
| Terms of [ formal ] conflict | |
| A. Function of Feeding. |
B. Function of Feeding. |
| Need for devouring of a possibly large quantity of vegetable material -- grassy plants or leaves of woody plants. | Need for strong grinding resulting, in finer particles, of vegetable material for complete digestion. |
Possible set of external conditions.
In conditions of the presence of unfavorableness in pastureland -- abundance of predators and the necessity often to run -- A turns out to be the chief term of the conflict, that is, devouring of a possibly large quantity of vegetable material. The lesser term, B, changed : In the animals there was developed the ability to swallow unchewed food, which later on, in a safe resting place, after regurgitation, is ruminated (chewing the cud). Such a solution of the conflict was one of the prerequisites for the formation of the group of ruminants ( Ruminantia ).
Alternative set of external conditions.
In conditions of comparatively less adverse circumstances in the pasture, that is, of a lower number of predators, and better running abilities, B turned out to be the chief term of the conflict -- the need for the highest degree of grinding of food when it is eaten. The lesser term, A, -- devouring of possibly large quantities of food -- was perfected. In the animals there were developed particularly strong lophodont teeth, which made it possible to grind plants when eaten. At the same time in the intestinal tract there developed a special symbiotic fauna of flagellate protozoa assisting digestion. Such a kind of solution of the conflict turned out to be a prerequisite for the formation of some groups of hoofed mammals, namely the odd-toed ungulates ( Perissodactyla ).
This concludes the list of examples, given by ROHDENDORF, of internal conflicts and their solutions as they are supposed to be achieved in the history of organisms.
In the next document we will continue to investigate the general features of organic evolution and try to give all of them a rationale in terms of our extension of the concept of Reality by the concept of the Implicate Order. This next document represents Part I of the thematic study of the mentioned general features.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for a full study of the general features of organic evolution and their interpretation in terms of our theory of the Explicate/Implicate Order, Part I.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V