
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) Following ROHDENDORF, 1964, pp. 30, we discuss the infraorder Nymphomyiomorpha wholly in terms of the family Nymphomyiidae which is the only family known in this infraorder.
Extent, evolution, and system
This very little known group of Diptera was first described in 1935 on the basis of a single species, Nymphomyia alba TOK., found in the mountains of Japan. Apart from the description of the structure of the body of the winged insect and pupa and some very poor information concerning the ecology, up to now [1964] nothing is known of this extraordinary peculiar form. Fossil documents about the history of the Nymphomyiidae are totally absent. The idiosyncratic organization of these insects only allows them to be near to the group Dictyodipteridae [of which three genera are described by ROHDENDORF], which is known only from upper Triassic sediments in Central Asia.
See next Figure.
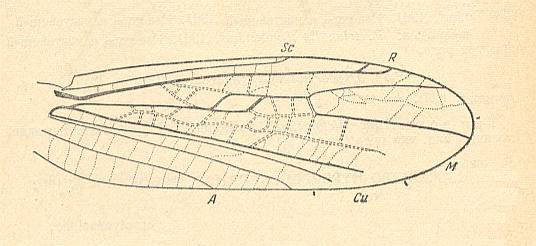
Figure 1 : Wing of Dictyodiptera multinervis ROHD. ( Dictyodipteridae, [extinct] infraorder Dictyodipteromorpha), Triassic of Central Asia. Length 2.81 mm.
(After ROHDENDORF, 1961, from ROHDENDORF, 1964)
[The geological age of the site where the fossil of Figure 1 was found turned out to be wrong. Its age is not (upper) Triassic but lower Jurassic. This is stated in ROHDENDORF & RASNICYN, 1980, p. 115, in Istoritjeskoje razwitije klassa nasjekomig ( = Historical development of the Class of Insects). This is apparently the case for all fossil insects reported to be found in Issic Kul (close to the western border of China)]
The similarity of these diptera [Nymphomyiidae and Dictyodipteridae] can be seen in the shape of their wings, and in this mainly with respect to the shape of the wings of the pupa : their margins are parallel and the wing-blade is moderately pointed at its apex. Placing the Nymphomyiidae near to the extinct Triassic Dictyodipteridae [which themselves belong -- according to ROHDENDORF -- to a distinct infraorder] is indirectly confirmed by the extreme [morphological] distinctness of the Nymphomyiidae from all other recent diptera and forces us to assume that their [historical] connections date from very long ago. [The mentioned morphological isolation of each of these forms might point to a polyphyletic (instead of monophyletic) origin of the order Diptera, and moreover a polyphyly that did not stop after diptera having been arised independently from two (mecopteroid) stocks, but a true polyphyly in which diptera have originated many times independently from a comparatively large number of (mecopteroid) ancestors. We could for example surmise that each of ROHDENDORF's dipterous infraorders arose independently from a different mecopteroid stock, or even that each family of diptera arose in this way, independently of the other families.]
Chief features
Some aspects of the morphology of the adult (nymphomyiid) fly are given in the next two Figures :
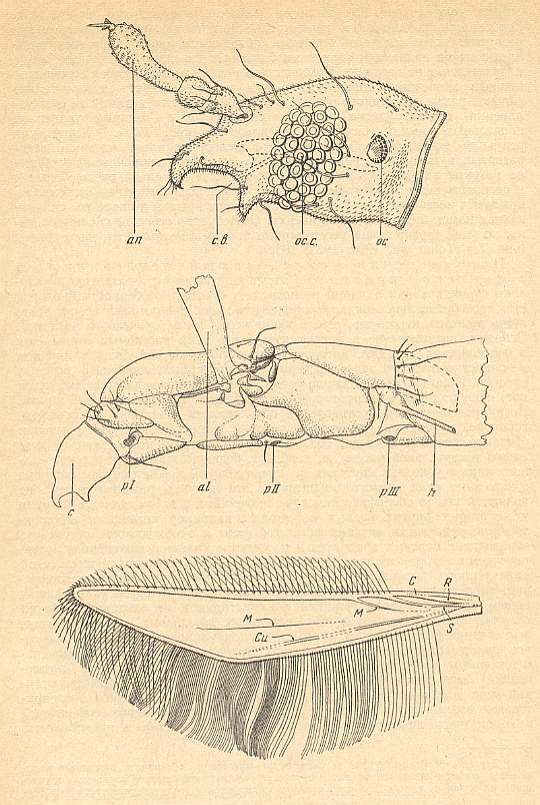
Figure 2 : Nymphomyia alba TOKUNAGA ( Nymphomyiidae), recent fauna of Japan.
Upper image : Head, lateral view.
Middle image : Thorax, lateral view.
Bottom image : Wing.
al = wing-base, an = antenna, c = head, c.b. = mouth-cavity, h = halter, oc = ocellus [simple eye], oc.c = compound eye, pI, PII, PIII = sites of the attachment of respectively the front-, middle-, and hindlegs, S = Radial Sector, C, R, M, Cu = veins of the wing.
(After TOKUNAGA, 1935, from ROHDENDORF, 1964)
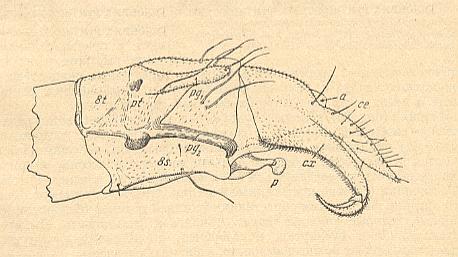
Figure 3 : Nymphomyia alba TOKUNAGA ( Nymphomyiidae), recent fauna of Japan.
End of abdomen of male, lateral view. a = anal aperture, ce = cercus, cx = coxite of ninth segment, p = edeagus, pg1, pg2 = paired outgrowths of the paratergite of eighth segment, pt = paratergite (lateral plate) of eighth segment, 8c, 8t = eighth sternite and tergite.
(After TOKUNAGA, 1935, from ROHDENDORF, 1964)
The discovery of the nymphomyiids in conditions of mountainous relief, nearby a mountain stream, in the benthos [bottom flora and fauna] of which apparently live the yet undiscovered larvae of these insects, is very remarkable. Such conditions form a characteristic ecological niche for various relict forms finding in it a refuge, and thanks to a working-out of a narrow, but perfect stenobiosis the majority of their closest relatives did survive.
Conflicts and determining tendencies in the historical development
The absence of data concerning the [individual] development and the conditions in which the larva lives makes it impossible to uncover the biological features of these most remarkable insects more or less precisely, and only allows for noting some most general features of their historical development.
The basic conflict in the life-activity of these insects expresses itself in the features of feeding, feeding that is, that can only be realized in the larvae, being totally absent in the adult winged insects lacking mouthparts. The development of aphagia took place very early on and was accompanied by (1) a weakening of the legs, (2) paleopteria --
[Note of ROHDENDORF]
The terms "paleopteria" and "neopteria" naturally signify the different position of the wings of the insect at rest.
The absence of the folding-back of the wings along the body thereby orienting them backwards, that is, the position which is visible in ancient insects ( Palaeoptera) [such as mayflies and dragonflies] and also [here and there] in ' Neoptera ' as in ancient moths [Lepidoptera], bee flies [ Diptera, Bombyliidae], and some other groups, can be called paleopteria.
Folding the wings backwards along the abdomen, the wing-blades partially approaching each other, at rest lying on top of each other, which is the case in the majority of Neoptera [like in most flies, bees, wasps, but also in grasshoppers, most moths, etc.], can in turn conveniently be called neopteria.
-- and (3), probably, [was further accompanied by] the shortening of adult life.
The origin of this conflict was, in the history of the Nymphomyiids, apparently determined by the deep-seated differences of the living-conditions of the respective phases of individual development : The larvae living in a habitat that is rich in organic substances (in the form of living plant masses), and, contrasting with this, the scarcity of the animal and plant world right in the living-conditions of the originally predatory winged phase (conditions of bare rocks of a mountainous terrain). These causes of the development of aphagia are not typical of nymphomyiids. It is observed also in the history of other aquatic insects, for example in Mayflies [Ephemeroptera] and Chironomidae : The insufficient feeding of the winged phase, the necessity of guaranteeing the formation of sexual products, and [the necessity of] colonization [together] constitute the major conflict in the history of many groups of diptera having determined the realization of a characteristic further development. Undoubtedly, the mentioned conflict which consists in the features of feeding of the Nymphomyiids is the latest acquisition in their history [and thus is responsible for merely the latest morphological features of them] and does not explain the deep-seated differences between these insects and all other diptera, which [differences] externally express themselves in the structure of their pupae, their apparatus of locomotion, and their sense-organs (large simple eyes [ocelli] ). The absence of data on the organization, and hence on the living-conditions, of the larva which is the chief stage (feeding stage) of the nymphomyiids, preclude the possibility to uncover the essence of the differences that distinguish these insects from the other diptera. It is hardly favourable to present preliminary conclusions about the nature of feeding of the larva based on features of the pupa (worm-like appearance, and apparent mobility) and suppose, for example, a particular hidden way of life of the larva under conditions of abundancy of nutritive substances, or judge the life-activities of the winged insect on the basis of just a superficial inspection of its morphological features (powerful flight-apparatus, qua type coming close to that of broad-wingedness [the wings being broad as a result of the very long hairs on fore and hind margin] ). Comparison with the extinct Triassic Dictyodipterids, who possessed little-costalized wings, indicates possible pathways of the development of the broad-wingedness of the Nymphomyiids. Such a conclusion is all that we can obtain in clarifying the pathways of the historical development of these two groups. Beyond this, the discussion becomes almost completely hypothetical.
Wrapping up our consideration of the features of the Nymphomyiids, we must note just the most general and latest conflict in their history, having expressed itself by the aphagia of the winged phase and note as determining factors of development : the formation of peculiar totally refined larvae which became the chief feeding stage of the [individual] development of the insect.
Conclusions
The survey of those few expositions about the Nymphomyiids that exist in the literature did allow us only to draw very general and preliminary conclusions. Apparently the determining tendencies in the history of the Nymphomyiids were the refinement of individual development, namely of the organization of the larval phase. Together with this we can see processes of degeneration of the winged phase which became aphagous, that is, a process of desimaginization [ = driving back the imaginal stage in favor of the larval stage]. Because of the absence of enough data, clarification of the essence of the basic differences of this peculiar group of insects, [these differences] placing them far away from all other representatives of the Order, cannot be done with the required exactitude.
Additional data
To the data as obtained from ROHDENDORF's 1964 account of the Nymphomyiidae we can add the following data taken from RICHARDS & DAVIES, 1977, Imms' General Textbook of Entomology. Here the Nymphomyiids are placed into the superfamily "Psychodoidea" as the family Nymphomyiidae together with two other families, viz., the Tanyderidae and the Psychodidae. The description they give reads :
Fam. Nymphomyiidae. Small flies with reduced venation, with long fringes to the wings, mouthparts atrophied, antennae with three stout segments and a short style, third segment sometimes annulated, eyes meeting beneath head but separated above, 2 ocelli present. Larvae aquatic.
These extraordinary flies were discovered by Tokunaga in 1932 in a fast-flowing stream, in Japan. He has since published several studies of this species which seems to be a highly specialized Nematoceran, retaining, however, some archaic features. The larva has not been described, but the pupa shows some affinity to the Psychodidae. Ide (1965) and Cutten and Kevan (1970) have described two more genera, each with a single species, from the E. Himalayas and Canada. The second paper also describes the larva which has 7 abdominal and 1 anal pair of pseudopods bearing claw-like pectinate spines. It does not at all resemble a Psychodid larva.
[ I -- Jaap Bax -- should like to add that the morphology of the winged phase and, according to the last remark of the mentioned textbook authors, of the larva, of the representatives of this family clearly point to its singularity within the rest of the Diptera. Probably the same (but to a lesser extent) can be said of the two other groups -- discussed in the next two documents -- the Deuterophlebiidae and the Blephariceridae, which are, just like the Nymphomyiidae, regarded by ROHDENDORF as separate infraorders of the order Diptera.
While these infraorders, and also some other such infraorders (such as Termitoxeniomorpha, Streblomorpha, and Nycteribiomorpha) clearly, as it seems, stand alone and independently within the rest of the Diptera, that is, isolated from them, the other infraorders as recognized by ROHDENDORF, such as the Tipulomorpha, Bibionomorpha, Asilomorpha, etc., might also stand on themselves, that is, having their own features separating each one of them from the other infraorders. And thus it might be that each infraorder is formed as a result of its own history, independently of that of the other infraorders. And maybe this is the case already at the family-level. Indeed, in constructing a genealogical tree of the Diptera the genetic connections of the families with each other must be projected farther and farther back into geological history because of the often occurring phenomenon of crossing of specializations : If family "A" possesses some specialization (i.e. a specialized state of a given feature), which another, but similar, family "B" does not, then family A might evolutionarily be derived from family B. But when at the same time family B is, with respect to some other feature, specialized as compared with family A, then one cannot evolutionarily derive Family A from B, because then one is forced to assume an evolutionary course running backwards with respect to the mentioned feature. And one cannot do so according to a general and widely accepted (and, I think, legitimately so) law of evolution (Dollo's Law). This Law states that a structure that has been evolutionarily lost cannot evolutionarily come back again, unless it is now formed by other morphological units, different from, that is, not homologous to, the original building-blocks. And because of the same reason (but now with respect to the other feature) family B cannot evolutionarily being derived from family A. To solve this problem one projects the point of divergence of the two families farther back into geological history, up to a time where one can assume that the one or the other feature was not yet in its specialized state. With all this, the common ancestor of the two families often becomes a higly hypothetical entity, perhaps not even biologically viable.
This phenomenon of having to project the alleged common ancestor backwards in time almost indefinitely is well-known in evolutionary research. And according to me it points to a polyphyletic origin of many groups, which here means the independent and genealogically isolated evolutionary development of each of the subgroups of such a group. In our case it could be that the infraorders of Diptera, or perhaps even most of the families of Diptera, have originated and evolutionarily developed independently of each other. And this means to suppose a polyphyletic (as contrasted with a monophyletic) origin of the Diptera as a whole! We will investigate this interesting problem in a later document.]
This concludes our exposition of the infraorder Nymphomyiomorpha. In the next document we will deal with the infraorder Deuterophlebiomorpha.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XII.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V
Back to Evolutionary Part VIII