
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) The functional wing types present in the superfamily Bibionidea.
Ancient lifting many-veined (bibionoid) type
Next we present a description of this type as such. As one can see, it occurs not only in the superfamily Bibionidea, but also in other superfamilies (such as the Scatopsidea).
Representatives of the type
Wings of the ancient lifting many-veined (bibionoid) type are possessed in the recent fauna by representatives of the families Bibionidae, Amasiidae, Axymyiidae, Scatopsidae, Synneuronidae, and Canthyloscelididae.
Size of the wings
The absolute size of the wings are variable. They vary from 2 to 10 mm. The relative size of the wings is such that their size is equal to that of the body or larger. The surface area and load of the wings are only known for one species of the family Bibionidae : The surface area is equal to 0.119-0.218 cm2 . The load is equal to 0.023-0.037 gr/cm2 .
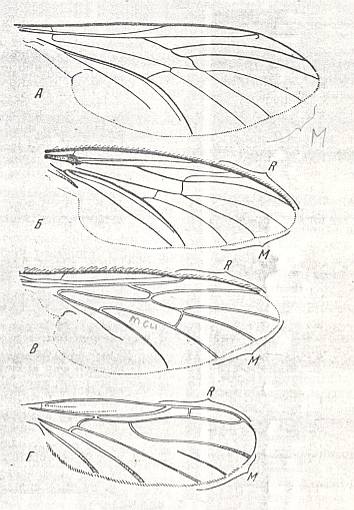
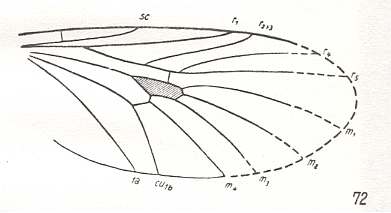
Figure 1 : Wings of recent representatives of the ancient lifting many-veined (bibionoid) type. From top to bottom :
[Here I assume that the names "Canthyloscelididae", "Hyperoscelididae", and "Corynoscelidae", signify one and the same single family, or at most three slightly different families. The family Synneuronidae is probably close to this complex. They all belong to the superfamily Scatopsidea.]
[Although the families Canthyloscelididae and Scatopsidae are closely related to each other (both belonging to the superfamily Scatopsidea), it is not clear to me why ROHDENDORF includes the wings of the Scatopsidae in the present functional wing-type. As we can see in Figure 2, below, veins in the wings of Scatopsidae (if the depicted species indeed represents the wing-venation of the family) are sparse in the sense that their density is (in most parts of the wing) quite low. This contrasts with the other members of the type.
We propose to remedy this as follows :
In principle a given typological boundary, separating two different types, may run right through a genealogical taxon, resulting in a part of this taxon (i.e. a subset of its representatives) belonging to one type, while the other part belongs to another, different, type. In the same way one and the same given type can be represented in several different not genealogically related taxa. But, for theoretical reasons, the discrepancy between the typological system, be it of functional wing-types, ecological types, or whatever, and the genealogical system (in the sense of HENNIG) may not be too great. The nature and variability of typological features is held within limits, limits as set by the immediate genealogical tree to which the species having certain given typological features belong. This means that a given type, such as a wing-type, cannot be, or is at least not expected to be, present in representatives of genealogically widely separated taxa. Even if -- to stick to wing-types -- the wings in such taxa formally look almost identical, they must be in some way different. These differences might be seated in the basiala, in the nature of the flight-muscles, or in some other allegedly minor structural features. On the other hand seemingly clearly different types might not be so different afterall, because they are possessed by, say, two, genealogically closely related taxa. The types can nevertheless be different, but the one type is, in such a case, apparently, the result of a more or less simple transformation of the other type which was also present in the common ancestor of the two taxa.
In this way, perhaps, we must approach the seemingly great typological difference between on the one hand the wings in the family Scatopsidae, and the wings in the complex Canthyloscelididae, Corynoscelidae, Synneuronidae on the other.
In order to make this more clear we will add some more Figures of wing-venation of relevant species and some comments, and integrate it with ROHDENDORF's text.
ROHDENDORF himself has also seen the great differences within the present type, that is the ancient lifting many-veined (bibionoid) wing-type. Therefore he, in a more or less integrated way, distinguishes four different subtypes (without naming them, but let us call them I, II, III, and IV) within this type, respectively containing the following families] :
[With this scheme in mind it will be easier to follow ROHDENDORF's text from where we take it up again (We will say so when we do).
As a last remark concerning ROHDENDORF's description and exposition of the present wing-type we say that part of the name given to it by ROHDENDORF is not very significant, namely the part "many-veined" in ancient lifting many-veined (bibionoid) type. Precisely as "many-veined" this type is not significantly contrasting with the representatives of the ancient lifting venationally attenuated (fungivoroid) type. We take it that in naming the present type -- the bibionoid wing-type -- ROHDENDORF has called it "many-veined" after the nature of its most primitive and oldest representative, and that is certainly Eoplecia : see Figure 9, below ]
In HENNIG, 1954, we find a drawing of the wing-venation of yet another species of the genus Amasia, and this genus is included in the family Bibionidae by HENNIG :
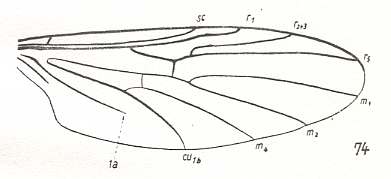
Figure 1a : Wing-venation of Amasia funebris MEIG. ( Bibionidae [or : superfamily Bibionidea, family Amasiidae] ).
(After HENNIG, 1954)
[Some more Figures should be added here] :
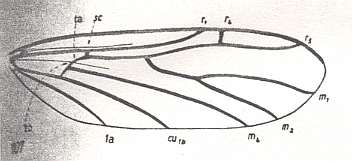
Figure 1b : Wing-venation of Corynoscelis eximia BOH. ( Corynoscelidae).
[The species eximia BOH. we have, as it seems, already encountered in Figure 1 (bottom image), there placed into the genus Eucorynoscelis. As can be seen, they differ with respect to M2. But this could be attributed to mere individual variation within one and the same species.] (After DUDA, 1928, from HENNIG, 1954)
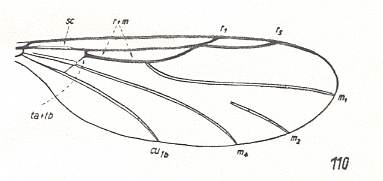
Figure 1c : Wing-venation of Synneuron annulipes LUNDSTR. ( Corynoscelidae [ or: superfamily Scatopsidea, family Synneuronidae] ). In addition to the coalescence of the anterior medial vein with the Radial Sector we see here a coalescence, at one point only, of the Radial Sector and the Radius.
(After DUDA, 1928, from HENNIG, 1954)
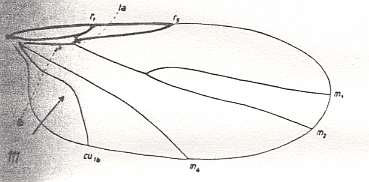
It seems that in the complex Canthyloscelididae-Corynoscelidae-Synneuronidae there is a tendency to reduce the venation eventually leading to that of the family Scatopsidae. The next Figure depicts a wing of a representative of this family which is supposed to include wings of the present type.
In addition to the wings of the Scatopsidae the transformational tendencies in the above mentioned complex (or perhaps some group already within the Scatopsidae) may also have led to the wings of the Cecidomyiidea [ = Itonididea] :
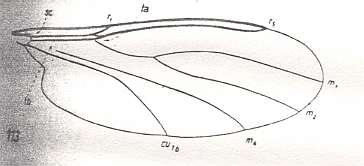
Figure 2a : Wing-venation of Lestremia cinerea MEIG. ( Itonididae [or : superfamily Cecidomyiidea (=Itonididea), family Lestremiidae)].
(After HENNIG, 1954)
[After having added Figures and comments, we will now continue with ROHDENDORF's text, where he successively discusses the features of the ancient lifting many-veined (bibionoid) wing-type, to begin with the shape of such wings] :
Shape of the wing
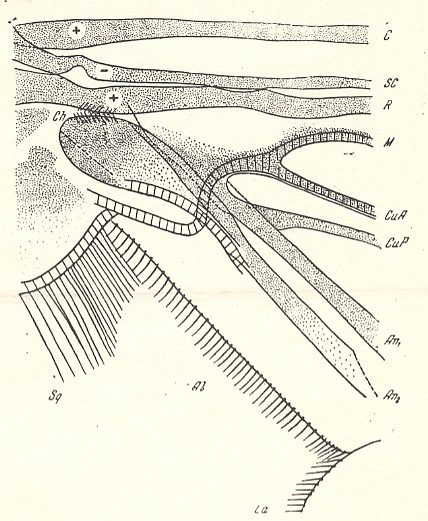
Figure 3 : Bibio japonicus DUDA. ( Bibionomorpha-Bibionidae). Wing-base, basiala. Schematic.
Abbreviations as usual. Ch = chaetarium (chaetarium radiale and chaetarium anale).
(After ROHDENDORF, 1946)
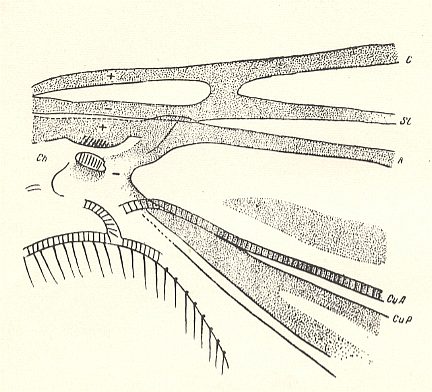
Figure 4 : Basiala of wing of Corynoscelis exima BOH. ( Corynoscelididae). Schematic.
Abbreviations as in previous Figures : from top to bottom : Costa (C), Subcosta (SC), Radius (R), Anterior Cubitus (CuA), and Posterior Cubitus (CuP).
(After ROHDENDORF, 1946 )
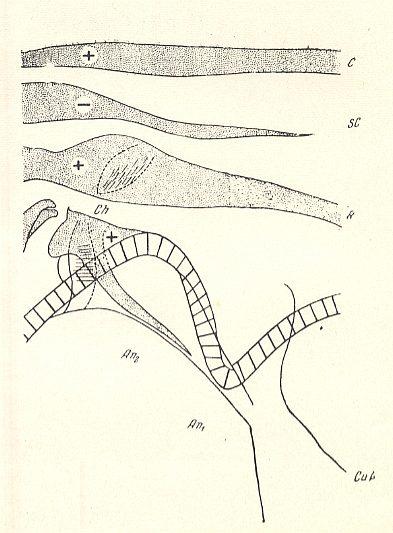
Figure 5 : Basiala of wing of Scatopse notata MEIG. ( Scatopsidae). Schematic.
Abbreviations as in previous Figures.
(After ROHDENDORF, 1946 )
Skeleton of the wing
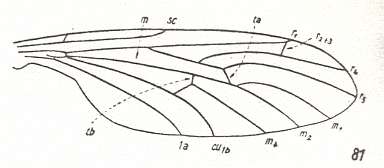
Always three medial branches present. Cross-veins well expressed only in Amasiidae and Axymyiidae (r-m and mcu) and partly so in Canthyloscelididae (only mcu) [the r-m cross-vein has disappeared as a result of the coalescence of the last radial vein and the first medial vein (see Figure 1, bottom image )]
In the majority of forms of the main families Bibionidae and Scatopsidae the cross-veins are, in connection with the development of costalization, strongly modified, being placed differently, or even having been disappeared completely. As a rule the basiala lacks a phragma. Only in the Canthyloscelididae there is developed a peculiar phragma which connects the bases of the radial and cubital veins, by which the wings of these insects are clearly distinguished from the other representatives of the type (see Figure 4, above and compare it with the Figures 3 and 5). There are, in the basiala, large and dense, well expressed chaetaria at the base of the radial and second anal vein.
Coverings of the wing
Functional characteristics
[Before giving ROHDENDORF's exposition of the history of the present wing-type and its subdivision into four subtypes (mentioned ABOVE ), we include here parts of RODENDORF's 1946 discussions of the fossil families Paraxymyiidae, Protobibionidae, and Protoscatopsidae, especially where he discusses the question asking to which recent families (such as Axymyiidae, Bibionidae, Scatopsidae) they are closest. We include these discussions because they not only teach us something about the mentioned fossil families (which are afterall often based on single poorly preserved fossils) but also about the recent families themselves, especially how to interpret the venation of their wings. In addition we shall add Figures and comments.]
Family Paraxymyiidae (Jurassic).
Based on Paraxymyia quadriradialis ROHD. :
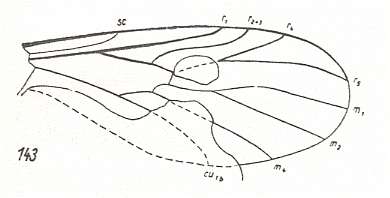
Figure 6a : Wing-venation of Paraxymyia quadriradialis ROHD. ( Paraxymyiidae). Jurassic. Interpretation of wing-venation according to HENNIG. Length of wing 2.25 mm.
(After ROHDENDORF, 1946, from HENNIG, 1954)
Sc [subcosta] moderately short, thin, but clear. It ends up in C [costa]. Rs [radial sector] branches off from R [radius] with a sharp angle and consists of three strong branches that end up in C. The cross-vein r-m lies in the middle of the wing between the two branches that branch off from the trunk of Rs. M [media] weak, having three branches. The common trunk of the medial system of veins is reduced, it is visible [in the fossil] in the form of a weak trace lying along the large basal cell. Cu [cubitus] thin. Anal veins not visible [in the fossil]. The wing is fairly broad.
The described group is idiosyncratic and possesses very primitive features, such as the presence of a radial sector with three branches, all of them strongly expressed and well-developed, as well as some, undoubtedly, features expressing specialization, such as the reduction of the common medial trunk, the weakly expressed Sc, the stronger development (thickening) of the veins of R and Rs together with the weakening of M and Cu.
What then are the relationships of the Paraxymyiidae with the other [nematocerous] Oligoneura? The most characteristic feature of specialization of the wing of this group consists in the reduction of the basal part of the medial trunk which also can be observed in the recent families Mycetobiidae (for Mycetobia pallipes MEIG. see HERE ), Macroceratidae (for Macrocera lutea MEIG. see HERE ), Ceroplatidae (for Zelmira fasciata MEIG. see HERE ), Ditomyiidae (for Ditomyia fasciata MEIG. see HERE ), Diadocidiidae (for Diadocidia ferruginea MEIG. see HERE ), Axymyiidae (for Axymyia furcata McATEE. see HERE ) [here the medial trunk is present, but, I assume, weakly so], Pachyneuridae (for Pachyneura fasciata ZETT. see HERE ) [here the medial trunk is weak], and some others. Another important feature of the structure of the wing in Paraxymyiidae consists in the position of the branches of the Radial Sector, of which the first [that is, the most proximal] one branches off from the common trunk [of Rs] significantly proximally from the cross-vein r-m. This latter cross-vein thus lies between the two anterior branches of Rs [that is, lies on that level].
Such a positioning of the anterior branch of Rs is, among Nematocera, observed in only a few families, such as in Axymyiidae (for Axymyia furcata McATEE. see HERE ), Phryneidae [ = Rhyphidae] (for Phryne fuscipennis MACQ. see HERE ), and for a part [that is : approximately or almost] in Mycetobiidae (for Mycetobia pallipes MEIG. see HERE ) and Pachyneuridae (for Pachyneura fasciata ZETT. see HERE ). This process of specialization of the wing-venation is, in the mentioned families, apparently already a more or less final stage : At first there takes place the reduction of branches of the Radial Sector.
In Mycetobiidae (for Mycetobia pallipes MEIG. see HERE ) and Phryneidae [ = Rhyphidae] (for Phryne fuscipennis MACQ. see HERE ) the second branch [that is, the distal one of the two anterior branches of Rs] has completely disappeared, while in Axymyiidae (for Axymyia furcata McATEE. see HERE ) and Pachyneuridae (for Pachyneura fasciata ZETT. see HERE ) the two branches [originally branching off from the common trunk of the Radial Sector] have partly coalesced to form a single forked vein.
Fossils belonging to the upper Liassic group of Nematocera, especially the Protorrhyphidae (for Protorhyphus stigmaticus HANDL. see HERE ) and the Eopleciidae (see Figure 9, below ), show, with respect to the structure of the radial system, a great similarity with the Paraxymyiidae [see Figure 6a, above ]. Especially remarkable is the fact that the branching pattern of the Radial Sector as it is present in all mentioned Nematocerous families is common to that in the Brachycera-Orthorrapha and even in Brachycera-Cyclorrapha : In all these diptera the cross-vein r-m lies distally from the branching-off point of the first branch of the Radial Sector. See, for example the wing of Rhagio scolopaceus LW. (Rhagionidae) (top image) and of Erinna atra MEIG. (Erinnidae) (bottom image), both belonging to the Brachycera-Orthorrapha, HERE .
On the other hand, the high degree of specialization of the wing-venation -- reductions in the medial system, weakening of subcosta -- make it impossible to see the Paraxymyiidae as the ancestors of whatever recent group coming close to them in regard of the branching structure of the Radial Sector. The Axymyiidae, as well as the Paraxymyiidae, originated from ancestors which were close to the Liassic Eopleciidae. The Paraxymyiidae acquired a series of features of high specialization (here : features of reduction) while at the same time having preserved some primitive features.
Family Protobibionidae (Jurassic)
Based on Protobibio jurassicus ROHD. :
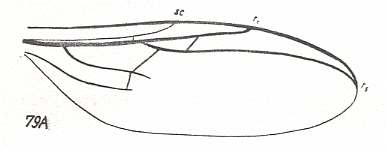
Figure 6b : Wing-venation (as far as preserved in the fossil) of Protobibio jurassicus ROHD. ( Protobibionidae). Jurassic. Length of wing 1.5 mm.
(After ROHDENDORF, 1946, from HENNIG, 1954)
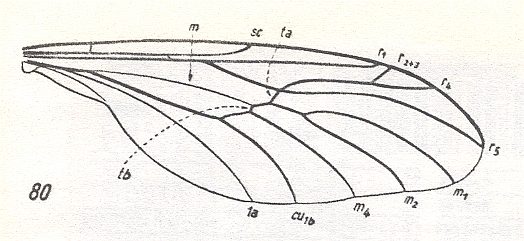
Sc [subcosta] well developed, a little shorter than half the wing. It ends up at C. Rs [radial sector] splits into two branches of which the longer (the posterior) branch reaches the wing-tip, while the short (the anterior) one ends up into R [radius] already quite a distance from the latter's end-point.The cross-vein r-m lies obliquely, and is long. M [media] and Cu [cubitus] weak. Because of bad preservation their structure is not clear. There is a transverse mcu. C, R, and Rs, strong, significantly thicker and darker than M and Cu. The wing is elongate, almost 3 times as long as wide.
The family Protobibionidae, described on the basis of a single genus, is very interesting : In itself it connects purely fungivoroid characters with those of the specialized wings of the recent family Bibionidae :
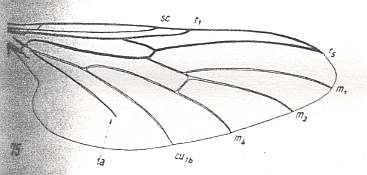
Figure 6c : Wing-venation Bibio marci L. ( Bibionidae). Length of wing about 8.5 mm. (After HENNIG, 1954)
The original nature of the branching-off of Rs from R places the genus Protobibio close to the families Allactoneuridae [recent] (see for Allactoneura nigrifemur ENDERL. HERE and Pleciofungivoridae [fossil] (see for Pleciofungivora major ROHD. HERE ).
On the other hand, the considerable length and oblique position of r-m, the significant thickening of the veins C, R, and Rs, brings the Protobibionidae close to the family Bibionidae, especially to the genus Philia (=Dilophus). The characters by which the Protobibionidae differ from the recent Bibionidae consist in the presence of the anterior, albeit short, branch of the Radial Sector, the stronger, but short, Subcosta, the different shape of the basal parts of M and Cu, and the presence of a true mcu. Some of these characters are clearly more progressive in comparison with Bibionidae (such, for instance, the shortened Sc). They do not make possible to see the Protobibionidae as direct ancestors of the Bibionidae. But it is evident that the Protobibionidae came from ancestors of Bibionidae not long before [the origin of Bibionidae] and should be placed in the direct proximity to that [phylogenetic] trunk.
Family Protoscatopsidae (Jurassic)
Based on Protoscatopse jurassica ROHD. :
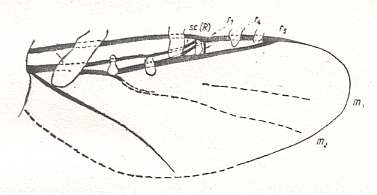
Figure 6d : Wing-venation (as far as preserved in the fossil) of Protoscatopse jurassica ROHD. ( Protoscatopsidae). Jurassic. Length of wing 2.0 mm.
Identification of veins according to HENNIG. The latter assesses the first vein after the Costa as subcosta, which might be problematic because of the considerable thickness of this vein in the fossil wing. ROHDENDORF assesses it as Radius, which is probably better despite the Radial Sector then being very peculiar.
(After ROHDENDORF, 1946, from HENNIG, 1954)
The venation of the wing of Protoscatopse is strongly contracted and shifted toward the anterior wing-margin. A large part of the wing is without veins, it is membranous. Sc is absent. R almost straight. Rs with three branches. The Costa [C] runs a little past the end of the last branch of Rs. It does not reach the wing-tip. M delicate and thin. It has two branches. Cu almost straight. It is strong only up to the middle of the wing, and continues in the form of a fold. The wing is fairly narrow.
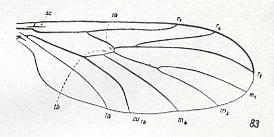
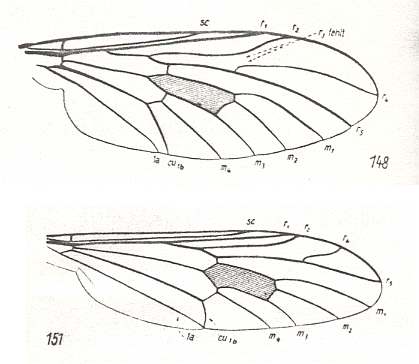
This newly described form is closest to the representatives of the recent family Scatopsidae, but differs from it by a series of clearly more primitive features. See Figure 2 which we reproduce here :
Such primitive features are : the presence of branches of the Radial Sector (developed two short anterior branches) lying in the cell between R and the main trunk of Rs. They demonstrate the secondary reduced condition of the venation of the system Rs in recent Scatopsidae. Then we have -- still in Protoscatopse -- the nature of the coalescence of M with Rs (these veins are here positioned idiosyncratically, being apparently coalesced along a certain distance). The point of branching-off of M from the common trunk M + Rs lies markedly more distally than the R-Rs fork, and lets us remember the wing-venation in representatives of the family Corynoscelidae [ROHDENDORF writes : Corynoscelididae], see Figure 1b (and 1c), above .
These features of the wing-venation of the described Jurassic form (Protoscatopse) allow us to come up with serious corrections as regards the existing interpretation of the wing-venation of recent Scatopsidae on the one hand, and clarify the mutual genetic connections of these groups on the other. Until now [1946] (Duda, 1928) the wing-venation in Scatopsidae is compared with that in true Oligoneura, namely with that in Lycoriidae [ = Sciaridae], and therefore it was held that Rs in its basal part has been transformed into a transverse vein and that the real radio-medial cross-vein (r-m) [ta in the next Figure] became integrated with the system of the straight strong vein M-Rs. See next Figure.
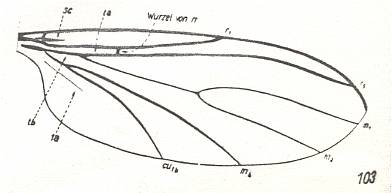
Figure 6e : Wing-venation of Lycoria bicolor MEIG. ( Lycoriidae). Compare with Scatopse fuscipes MEIG. ( Scatopsidae), Figure 2, above .
(After HENNIG, 1954)
The position of the veins in Corynoscelididae did not fit into this system, and therefore it was held that this group was an aberrant [phylogenetic] branch [See Figure 1c, above]. The clear relict- and very ancient nature of the Corynoscelididae was not taken into account by the majority of authors who ignored the very interesting and instructive wing-venation of these rare insects.
In comparing the wing-venation in Scatopsidae, Corynoscelididae, and the new Jurassic form [Protoscatopse) we directly see the remarkable similarity in the branching of the veins Rs and M between Corynoscelididae and Protoscatopsidae : This similarity consists in the coalescence of the trunks of Rs and M1+2, precisely as in Ceroplatidae, see next Figure :

Figure 6f : Wing-venation of Zelmira fasciata MEIG.
( Ceroplatidae-Macroceridae (=Zelmiridae)). (After HENNIG, 1954)
and [precisely as] in other related groups. Compare Corynoscelididae Figure 1c, above with Protoscatopsidae Figure 6d, above . Together with all this, the general nature of the specialization of the wing-venation, that is, its shift to the fore-margin and its strengthening there -- especially the mutual approach of the trunks R and M+Rs in Protoscatopsidae and Scatopsidae : Compare Protoscatopsidae Figure 6d, above with Scatopsidae Figure 2, above -- cleary demonstrates that the base of Rs, that is the Radial Sector's branching off from R, has been disappeared in Scatopsidae, and that the so-called "base of Rs" is in Scatopsidae no more than the shifted proximal anterior branch Rs-Rs1 .
Now it is the right place to investigate the question whether the described Jurassic group [Protoscatopsidae] is a direct ancestor of recent Scatopsidae and Corynoscelididae. Right from the beginning we must, answer this question, with a definite "no" with respect to the Corynoscelididae. This latter group carries a series of most primitive structural features, forbidding to see it as the descendant of the already highly specialized Protoscatopsidae. Such a primitive feature of the Corynoscelididae is the weak mechanical specialization of their wings. The family Corynoscelididae is undoubtedly a very ancient -- relict -- group, which branched off from the common trunk of "scatopsoid" forms much earlier than the Protoscatopsidae.
Something totally different we see in the relationships between Scatopsidae and Protoscatopsidae. The absence of any trace of a Subcosta can be accounted a specialization of Protoscatopsidae as compared with the recent Scatopsidae. All of the rest of the venation as described above lets me reckon this Jurassic group to be very close to the first original forms of the recent Scatopsidae.
After having clarified the position of Corynoscelididae and, especially Scatopsidae, we can now continue with the exposition of the Ancient lifting many-veined (bibionoid) wing-type, as described by ROHDENDORF, 1951. Already given was :
The history of the type and its transformations
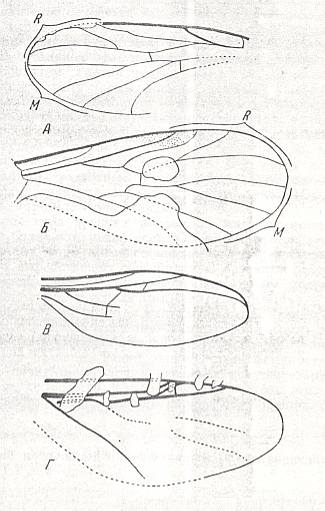
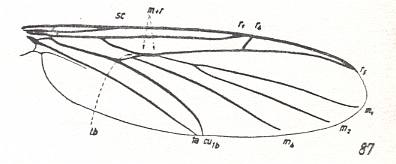
Figure 7 : Wings of fossil representatives of the ancient lifting many-veined (bibionoid) type. From top to bottom :
[As can be seen in Mesopleciella (top image in Figure above) the present type is hard to distinguish from the ancient lifting venationally-attenuated (fungivoroid) wing-type in which are included -- among others -- the wings of Hesperinus] :

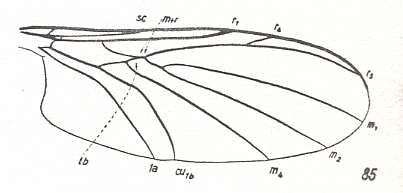
Figure 8 : Wing-venation of Hesperinus imbecillus Lw. ( Hesperinidae, Bibionidea). (After HENNIG, 1954)
[Compare this venation with that of Amasia funebris MEIG., HERE, which wings are supposed to belong to the present type.]
From the Jurassic fauna of Karatau are known representatives of the families Protopleciidae (3 genera, see Figure 7, top image ), Eopleciidae (one genus)
[At least until 1964 Eopleciidae have not been found in Karatau (middle or upper Jurassic). The only representative of this family is Eoplecia primitiva HANDL., found in the upper Liassic -- the Liassic is the oldest subdivision of the Jurassic -- of Mecklenburg (Germany, now Poland) and described by HANDLIRSCH] :
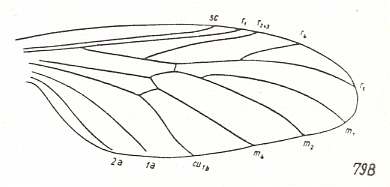
Figure 9 : Wing-venation of Eoplecia primitiva HANDL. Upper Liassic of Mecklenburg.
[It clearly belongs to the present wing-type.]
(After HANDLIRSCH, 1938, from HENNIG, 1954)
and Paraxymyiidae (one genus, see Figure 7, second image ), which possessed wings that let them be close to later Caenozoic and recent Amasiidae and Axymyiidae. In the same period (Jurassic) in this fauna (Karatau) there existed representatives of the other group of the type, namely species of the families Protobibionidae, see Figure 7, third image , and Protoscatopsidae, see Figure 7, bottom image , which already had specialized wings, that let them connect to the Caenozoic Bibionidae and Scatopsidae.
The above described heterogeneity among the representatives of the bibionoid wing-type evidently bears witness to the presence of secondary groupings, subtypes.
The first most primitive subtype is represented by the forms of wings in the recent Amasiidae, Axymyiidae, the fossil Eopleciidae, Protopleciidae, and Paraxymyiidae. This subtype distinguishes itself by weakly developed costalization in the wings, their large size, the presence of dense strong microtrichia on the wing-membrane. Flight in these insects is apparently slow and heavy : some non-flying species are known.
The second subtype is characteristic of the representatives of the families Bibionidae and Protobibionidae. Costalization is well expressed by the thickening and strengthening of the anterior veins. As in the previous subtype, strong and dense microtrichia are well developed. As regards the other features this subtype is close to the first one. Undoubtedly it is its derivative.
The third subtype sharply distinguishes itself from the two previous ones. It is characteristic of the rare relict forms of the family Canthyloscelididae [As has been said above, the "families" Canthyloscelididae, Corynoscelidae, Corynoscelididae, Synneurontidae [Synneuronidae], and Hyperoscelididae, apparently belong to a single group, which might be called "Corynoscelidae" or "Corynoscelididae".]. The most typical features of this subtype are : the development of a phragma in the basiala, see Figure 4, above , a rudiment [or beginnings] of the anal lobe, absence of costalization (see, for example, Figure 1b, above ), lengthening of the wing, and the development of micro- and macrotrichia.
This subtype is undoubtedly the source of the next and most specialized subtype of wings, that is developed in the representatives of the family Scatopsidae. This subtype distinguishes itself by an extreme development of costalization, complexification of the structure of the basiala, see Figure 5, above , a strong development of the anal lobe (see Figure 2, above ), shortening of the wing, and the presence of only short microtrichia.
The origin of the type [the type as a whole] of the ancient lifting many-veined (bibionoid) wings is more or less clear. On the one hand this type branched off from first forms of the ancient lifting venationally-attenuated (fungivoroid) type, which had worked out a distinct basiala and having reduced the venation, and in this way giving rise to the pleciofungivoroid subtype together with the first subtype of bibionoid wings (Protopleciidae + Amasiidae). On the other hand, the wing-subtype in the representatives of the family Canthyloscelididae is a derivative of one or another significantly more specialized form of wings, most probably of still unknown representatives of the fungivoroid type that had refined the basiala. This dual path of formation of the bibionoid type apparently precisely expresses the actual history and is, moreover, confirmed by the features of the [taxonomic] system of these insects : Bibionidea and Scatopsidea are undoubtedly way out distinct superfamilies.
(End of the exposition of the bibionoid wing-type, and at the same time the end of the exposition of the superfamily Bibionidea).
In the next document we will consider the superfamilies Scatopsidea and Rhyphidea, both also belonging to the infraorder Bibionomorpha.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part X.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V
Back to Evolutionary Part VIII
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}