
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) Having, in the previous documents, discussed the infraorder Tipulomorpha, we will now discuss the second infraorder, the Bibionomorpha.
Content of the infraorder Bibionomorpha
The infraorder Bibionomorpha is a large complex of two-winged insects and includes six superfamilies (Some alternative names of these taxa are also used. We indicate them by "syn." ) :
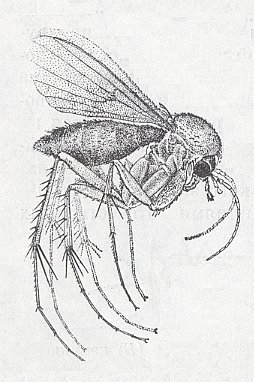
Figure 1 : Mycetophila (Fungivora) fungorum DEG. (Fungivoridae), 4-6 mm.
(After GORNOSTAJEV, 1970, Nasjekomije CCCP (insect guide))
General characteristics of the Bibionomorpha
The various representatives of this infraorder of Diptera can be generally characterized by a compact statement. The predominant way of life of the bibionomorphs is distinguished from that of the tipulomorphs by the absence of a connection with aquatic habitats : the overwhelming majority of forms of the bibionomorphs have their individual development away from the water, they are purely terrestrial insects, although they must live in places with high humidity. The larvae live in the soil [in layers of forest floor], in decaying plant remains [including rotting wood], in excrements of vertebrates, or in living plant tissues. Rarely there are [species] that live freely on the surface of the substrate (for instance, predacious Cecidomyiidae) and then, as a rule, in an environment that is saturated with water vapor.
Very rarely we encounter species of bibionomorph larvae living in a liquid medium (rotting plant materials, temporary accumulations of water or of plant juices on plants, and so on). They, however, never live in water-basins.
As a rule, the larvae possess anterior spiracles being amphipneustic or even peripneustic. Rarely they are metapneustic (only having posterior spiracles) or even having no spiracles at all (Ceroplatidae and others). Generally the larvae have a well-distinguished head, carrying biting [i.e. gnawing, chewing] mouthparts. And in a number of cases there has been developed extra-intestinal digestion and [with it] the reduction of the skeleton of the head [head capsule] and mouthparts (Cecidomyiidea, some Scatopsidea).
The larvae of the Bibionomorpha are for the most part phytophagous [not, however, living freely on (higher) plants of which they eat parts like caterpillars or saw-fly larvae do], some forms are detritophagous [It seems, however, that a great many, not 'some', bibionomorphs feed on vegetable detritus and many others on the fungi associated with that detritus. That is, the larvae of many of them inhabit compost-like masses of material.]
The winged phase (see, Figure 1 ) of the representatives of the bibionomorphs is characterized by the shortening of the body, an increase of the [relative] size of the thorax as a result of the strengthening of the musculature first of all of the longitudinal dorsal muscles pulling the wings down. The shortening of the body was accompanied with changes of the organs of locomotion : the legs were [in evolution] rarely lenghtened and generally belong to the running type and more rarely to the prehensile type. The wings were very often shortened and show different degrees of costalization, sometimes reaching extreme degrees (Scatopsidae). In some cases, in connection with extreme decrease in body size, the wings acquire traits of feather-wingedness [i.e. wings consisting of a median spoke, laterally carrying long hairs, as we see it in Thrips.]. With repect to feeding, in the winged phase of the Bibionomorpha we cannot single out features which would bear witness to some refinement, or to one or another narrow specialization : The majority of [adult] forms feed on liquid vegetable materials or are aphagous [that is, do not feed at all]. Predators and bloodsuckers are unknown.
The features of their reproduction are little refined : The eggs are small and are laid simply on the nutritive substrate. Rarely we will encounter differently built ovipositors (Cecidomyiidea) or other adaptations (Bibionidae) which enable the eggs being laid into concealed sites.
Conflicts and determining tendencies in the historical origin and development of the Bibionomorpha.
Indicating features characteristic to the infraorder Bibionomorpha as a whole is rather difficult as a result of the abundance and diversity of this group, which, after its long history, has reached a extensive development [that is, the group has, by way of its many species, colonized a wide variety of environments all over the world].
Evaluating the chief basic features that characterize the infraorder as a whole, makes it possible to indicate the conflicts which had determined the evolutionary development of these Diptera, that is, their distinctness as a single group within the order [of Diptera].
(1) Living in terrestrial environments, and, (2) connected with that, their individual development in fairly local conditions, (3) their not being connected with water-basins, (4) the initially not refined larval feeding of detritus or plant tissues, (5) the necessity of feeding by the adult fly, and (6), with it, the necessity of refined locomotion -- all this determined the origin of the first conflicts. In this way the detritophagia or the primitive phytophagia of the larvae determined [i.e. were responsible for] the insufficiency of larval feeding for letting to mature the sex products, and [consequently, in that way determined] the necessity of adult feeding. This ancient and widely spread conflict was one of the very earliest at the evolutionary establishment of the Bibionomorpha, and in the consequent history of the different groups of the series this conflict was solved in differently ways. For the ancient bibionomorphs this conflict remained to be basic and determined the refinement of the winged phase, that is, improved their locomotor apparatus (flight and running). The wings transformed into refined costalized organs, able to perform very rapid beats, and, as a consequence, able to develop a strong lifting power and with it the ability of fast take off. In other cases powerful legs were developed that enabled quick running amidst plant remains.
So the determining tendency in the history of the infraorder of bibionomorphs as a whole was centered around the refinement of the motion features of the winged phase of the insect. The features of the nervous system, of feeding, and of reproduction, do not characterize this infraorder of Diptera as a whole, and only in the history of individual superfamilies these features acquired, in some cases, the character of determining tendencies.
Some notes on the systematics of the Bibionomorpha.
Traditionally the order Diptera is divided into three chief groups, viz., the Nematocera, the Brachycera-Orthorrapha, and the Brachycera-Cyclorrapha.
The Nematocera are flies having (in the adult stage) multisegmented antennae (which multisegmentation often, but not necessarily, results in long threadlike antennae). To this group belong all crane-flies, gnats, midges, and mosquitoes.
The Brachycera-Orthorrapha all have short antennae, generally three-segmented. Like in the Nematocera, the opening through which the adult fly liberates itself from the pupa is a linear slit. To this group belong all horse-flies, robber-flies, and many others.
The Brachycera-Cyclorrapha also have short three-segmented antennae, but the opening through which the adult fly liberates itself from the pupa is not a linear slit but is circular in shape. These are all the higher flies like the house-fly, blue-bottle, etc.
Because presently we are discussing the Bibionomorphs, which are Nematocera, we leave -- for the time being -- aside the Brachycera-Orthorrapha and the Brachycera-Cyclorrapha.
The group Nematocera is traditionally considered to be a suborder of the Diptera. ROHDENDORF, 1964, however, thinks of this group to be too heterogeneous to represent a genuine taxon. Other autors, however, such as KRIVOSJEINA, 1969, and all western authors, consider the Nematocera to be a genuine taxon, a suborder alongside the two other mentioned suborders. We ourselves cannot decide on this matter, and consider both views as deserving attention.
The ' Nematocera ' were traditionally subdivided into two large groups, namely the Polyneura (generally having wings with a rich venation) and the Oligoneura (generally having wings with a relatively poor venation). These two groups roughly coincide respectively with ROHDENDORF's Tipulomorpha and Bibionomorpha ( In his 1946 treatment of fossil and recent midges ROHDENDORF still works with the Polyneura / Oligoneura division of the Nematocera, but in his 1964 book on the evolution of the order Diptera he already works with his 'infra-orders' among which are the infraorder Tipulomorpha and the infraorder Bibionomorpha, which are largely ecologically defined, that is, not exclusively on formal morphological terms.).
In his 1946 book on midges ROHDENDORF considers exclusively the Oligoneura, which division, except for the Pachyneuridae (in 1964 assigned by him to the Tipulomorpha), neatly coincides with the infraorder Bibionomorpha. He (1946) does not yet recognize superfamilies, but discusses all the existing families of the Nematocera Oligoneura one after the other.
We will reproduce (that is, paragraphing from the russian text) precisely that particular Chapter (namely Chapter 5) of ROHDENDORF's 1946 work which enumerates and concisely describes the individual families belonging to the Oligoneura with respect to their way of life (as far as known) and the degree of their relict nature. But before we do so we will show how ROHDENDORF in his later work (1964) has distributed these oligoneuran families over a number of superfamilies :
Infraorder Bibionomopha (Oligoneura)
Bolitophilidea
Bolitophilidae
Fungivoridea ( = Mycetophilidea)
Allactoneuridae
Ditomyiidae
Ceroplatidae
Diadocidiidae
Macroceridae
Manotidae
Lygistorrhinidae
Mycetobiidae
Fungivoridae ( = Mycetophilidae)
Sciaridae ( = Lycoriidae)
Cecidomyiidea ( = Itonididea)
Lestremiidae
Cecidomyiidae ( = Itonididae)
Heteropezidae
Scatopsidea
Hyperoscelididae
Synneurontidae
Scatopsidae
Bibionidea
Penthetriidae ( = Pleciidae)
Hesperinidae
Bibionidae
Rhyphidea ( = Phryneidea, Anisopodidea)
Rhyphidae ( = Phryneidae, Anisopodidae)
Olbiogastridae
Cramptonomyiidae
The reason why we concentrate our attention so much on expositions of ROHDENDORF, despite the fact that these are written so long ago, is that especially ROHDENDORF, as well as a number of other russian authors, shows, according to me, the right attitude of mind when it comes to interpret and investigate the evolutionary process of organisms, namely their insistence of the ecology of the organisms, that is, the relation and interaction between the organism and its immediate environment, being the chief factor, the chief driving force, of the evolutionary process. And further we appreciate their explicit insistence that higher-ranking taxa, such as families and superfamilies, possess a nature of their own and evolve in their own way.
We will now give ROHDENDORF's 1946 account of the Oligoneuran families of the recent fauna as he presents it in Chapter 5 (As always, we paraphrase the russian text, that is, we are not committing ourselves to a literal translation, because we want to be free to better explain things than is done in the text where necessary, and we also want to be free to add or omit things where necessary).
We give the number of species of a taxon under discussion (and the number of subordinated taxa) as given in the 1946 text (according to the latest update in 1936), and with respect to this number of species we add, between brackets, the corresponding number given in ROHDENDORF 1964, as it was updated in 1958.
Further, in the 1946 text, for the three main families of the Oligoneura the alternative names are used : Itonididae, Fungivoridae, Lycoriidae. Their synonymy is :
Itonididae = Cecidomyiidae
Fungivoridae = Mycetophilidae
Lycoriidae = Sciaridae
Figures of complete wings of representatives of the different families are added by me, when necessary, and taken from HENNIG, 1954.
Let us now then, finally, proceed with ROHDENDORF, 1946, Evoljutsia krila i filogenes dlinnoysich dwykrilich, Oligoneura (Diptera, Nematocera) [ = Evolution of the wing and the phylogeny of the long-antennal two-winged insects, Oligoneura (Diptera, Nematocera) ], Chapter 5, p. 34-38, where he discusses the recent fauna of oligoneurans:
The recent fauna of the investigated group of Nematocera Oligoneura includes more than 5700 [7377] species
NOTE of Rohdendorf :
The given numbers are taken from the latest record on the order Diptera, compiled by Hendel (1936), and do not, of course, exhaust the real abundance of the fauna of these diptera. The value of these numbers mainly consists in their expressing the r e l a t i v e specific weight in the fauna of each group, differently said, they express the weaker or stronger development of it at the present day.
distributed over 21 families. Of these species about 5/6 belong to three families -- Itonididae, Fungivoridae and Lycoriidae.
These three families of the Oligoneura are the chief flourishing groups, which, in the present era experience the moment of their greatest development.
The largest family is the Itonididae, with its more than 2900 [3173] species, widely distributed all over the globe, being encountered in all zoogeographic domains. It divides in 6 secondary categories -- subfamilies or tribes -- with a large number of individual genera and species.
Itonididae or gall-midges, are, as compared with all other families of Oligoneura, in the highest degree associated with the plant world. Their larvae live in tissues of living plants. They are specialized monophagous insects. The life of the adult midges is very short. Probably the majority of them is aphagous.
The second largest family of the Oligoneura -- Fungivoridae, or fungus-gnats -- although having only about half the number of species (about 1400 [2248] ) as has the previous family, it is surprisingly more diverse. It subdivides into no less than 8 secondary catgories -- subfamilies -- of very unequal size. Many of these subfamilies carry an evident relict nature and are poor in species.
This family enjoys a wide distribution over all zoogeographic domains. The regions of the relict subfamilies, poor in species, are very narrow or dispersed. The name of the family exactly communicates a trait of its biology : the larvae of the fungus-gnats feed on a variety of fungi : some groups live in the fruiting bodies of higher fungi, basidial- and pouch-fungi. The larvae of other groups feed on rotting wood which is infested with a mycelium of fungi. Monophagy is almost totally not expressed in these gnats. The Fungivoridae are most common in woodland. In steppes, and even more so deserts, they are poorly represented. The period of adult life is, in the majority of forms, probably long. The adult gnats hibernate.
The third large family of Oligoneura, the Lycoriidae, is significantly poorer in species than are the previous two. It includes about 500 [572] species, and is surprisingly more monomorphic than the two previous ones. It divides into a few secondary categories with a small number of genera, but [these genera] including a large number of species. This family is probably evenly distributed over the different zoogeographic domains, and is much less studied than the two previous ones.
The larvae of the Lycoriidae live in different rotting vegetable materials. The adults do not, as it seems, have a long life.
The remaining 18 families of the Oligoneura are more or less strongly expressed relict groups. At the present day they occur in limited areas and possess utterly specialized features together with the presence of many archaic features. This combination of sharply contrasting structural features -- highly specialized traits within a general background of primitive traits -- is very characteristic of relict groups and in fact determines the very essence of the relict. Predominance of specialized, or, on the other hand, of primitive features, clarifies the present condition of the given group -- its relict nature. Together with this the ultimate and the very chief correctivum [adjustment] concerning answering the question about the alledged relict nature, is the actual state of the given group at the present era, that is, the level of its development. The latter is determined [I would prefer : is expressed] by the number of species and genera, and by their distribution, that is, wide or narrow geographically and ecologically. Only the analysis of each individual case can provide an answer to the question about the factors of evolution of the given group.
The whole group of relict families of the Oligoneura can be divided into two parts which are totally unequal with respect to size and diversity :
The first group of relict families includes seven of them :
Bibionidae,
Ceroplatidae,
Scatopsidae,
Lestremiidae,
Penthetriidae ( = Pleciidae ),
Macroceratidae and
Heteropezidae.
They hold about 800 species.
The second group of relict families includes the remaining 11 families of the Oligoneura and consists of only about 100 species.
First group of relict families.
Among the first seven families three -- Bibionidae, Ceroplatidae and Scatopsidae -- distinguish themselves as being especially strongly developed -- they include about 550 [774] species. Together with this these families are unified by the presence of highly specialized features of the wing structure and the sense organs. The ranges of distribution of these families are very wide and they [i.e. those ranges] almost do not show characteristic relict features [for example the ranges to be small or dispersed]. The biology of the Bibionidae and the Scatopsidae is more or less similar : their larvae live in rotting plant and animal matter and in excrements.
The Ceroplatidae are fungus feeders, and are abundantly developed in the Australian region.
The Bibionidae include about 250 [388] species, distributed over two subfamilies and three genera, widely spread over all zoogeographic domains.
Species of the family Scatopsidae are less in number (about 100 [128] ), but distributed over a large number of genera, also widely spread over the globe. The Ceroplatidae do not differ significantly from the Bibionidae with respect to number of species. They include about 200 [258] species, distributed, however, over the relatively large number of 14 genera, 6 tribes, and 4 subfamilies.
The remaining four families [still of the first group] are relict families but relatively rich in species. In total they do not include more than 180 species. These families, the Macroceratidae, Penthetriidae ( = Pleciidae), Lestremiidae and Heteropezidae, are about equal in size, but far from equal in their features.
The Macroceratidae are highly specialized insects, which in their larval stage feed on fungi, and, in the adult stage having reduced mouthparts. The species of the single genus Macrocera are widely distributed in the Northern Hemisphere.
The Penthetriidae [Pleciidae], with 50 [102] species, consists of two genera and is most abundantly distributed in the Old and New World tropics. The biology of the larvae is little known. Probably they are vegetable saprophags [i.e. feeding on rotting vegetable matter]. The adult insects possess many archaic primitive structural features, and are, in addition to it, probably aphagous [which is a derived, that is, not a primitive, feature].
The Lestremiidae (about 50 [200] species) possess a fairly rich wing-venation [which as such is a primitive feature], together with specialized features of larval development. [Indeed,] the larva has lost its head capsule. The geographical distribution of this family is wide but insufficiently known. Up until now [1946] a large number of genera, poor in species, has been described.
The last family (of this subgroup), the Heteropezidae, is the smallest with respect to the number of species, counting only about 30 [30] of them, which number, however, depends, more so than in the other families, on the fact that this family of extremely small insects is still little studied. In the form of a small number of genera this family is widely distributed over all zoogeographic domains. The family characterizes itself in virtue of an extreme specialization : The adult insects have a very reduced wing-venation. And in the development of the larva the phenomenon of pedogenesis [reproduction taking place already during the larval phase] is observed (the only example in the order Diptera).
Second group of relict families.
In the second group of oligoneuran relict families, including not more than a hundred species, distributed, however, over not less than 11 families, the relict nature is most strongly expressed. Three of its families, Mycetobiidae, Ditomyiidae, and Bolitophilidae, are relatively 'rich' in species.
The Mycetobiidae includes some 15-20 [8] species of the genus Mycetobia and some closely related tropical genera. The family is little studied : the larvae live in hollows in trees. More precisely their biology is not known. The structure of the larvae is little specialized. The adult forms possess a rich but strongly specialized wing-venation. See next Figure.

Figure 2 : Wing of a member of the Mycetobiidae
82 -- Mycetobia pallipes MEIG.
(From HENNIG, 1954, after FREEMAN, 1951,)
The family Ditomyiidae is about equal in size as the previous family. The geographical distribution of its few species is not sufficiently known. The larvae are, as compared with the Mycetobiidae, significantly more specialized and live in fungi. The features of the adult forms are, on the other hand, clearly more archaic and their wing-venation is not costalized (that is, there is no crowding of veins or strengthening of them at the anterior margin of the wing). See next Figure.
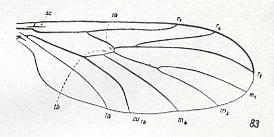
Figure 3 : Wing of a member of the Ditomyiidae
83 -- Ditomyia fasciata MEIG.
(From HENNIG, 1954, after FREEMAN, 1951,)
The family Bolitophilidae includes about 20 [32] fairly rare species of the one genus Bolitophila distributed over all zoogeographical domains. The larvae are very specialized and live in fungi. The adult midges possess a series of archaic primitive traits in their wing-venation and a wing-base (basiala) that is little specialized. In it the usual reduction of the 'handles' (thickened proximal parts) of the cubital and anal veins is barely expressed. See next Figure for the overall wing-venation.
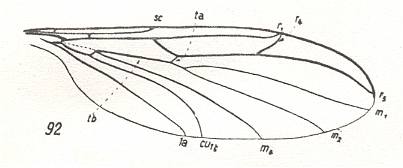
Figure 4 : Wing of a member of the Bolitophilidae
92 -- Bolitophila hybrida MEIG.
(From HENNIG, 1954, after TONNOIR & EDWARDS, 1927)
The last eight families [of the present group] are the most strongly expressed relict forms : All of them are rare insects, which have widely separated areas-of-living (natural habitats), or, on the other hand, being limited just to a single small region. Further, these insects usually possess different primitive features which are always accompanied by highly specialized features. As a result of the significant rarity of these insects their biological features are known for only a few families. This is the case with the two families Diadocidiidae and Corynoscelididae. The biology of the remaining six families is still unknown.
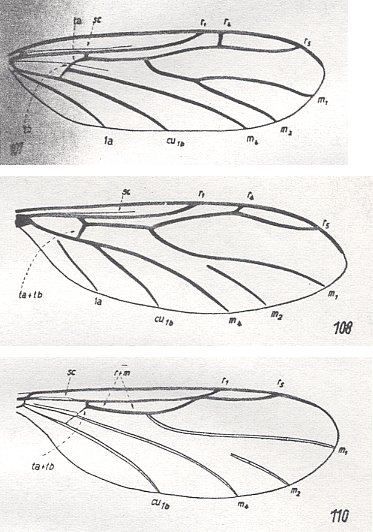
Figure 5 : Wings of Corynoscelididae
107 -- Corynoscelis eximia BOH.
108 -- Canthyloscelis antennata EDW.
110 -- Synneuron annulipes LUNDSTR.
(From HENNIG, 1954, after DUDA, 1928)
Extreme specialization appears in the larvae which lack a head capsule and having a peculiar biology.

Figure 6 : Wing of a representative of the Diadocidiidae.
89 -- Diadocidia ferruginea MEIG.
(From HENNIG, 1954, after EDWARDS, 1925)
The known 3 or 4 [5] species of the family Pachyneuridae [ This family is later (1964) subsumed under the Tipulomorpha (Nematocera Polyneura) by Rohdendorf ], are perhaps most interesting with respect to structural features of the wings which, in this family, are the least specialized as compared to the wings in the other Oligoneura : In the wing any soft and elastic membrane at the wing-base (basiala) is absent, which wing-base does not possess adaptations to enable 'propeller-flight' [that is, to enable the wing to 'imitate' the movement -- and especially the effect -- of a rotating propeller (airscrew) ]. The 'handles' [i.e. the basal sections] of almost all wing-veins are well developed. They are not reduced. The specialization of the wing only consists in the reduction of the basal section of the median vein. See Figure 9 of the previous document . For an overview of the whole wing-venation see next Figure.
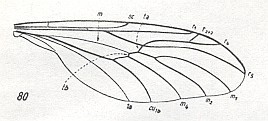
Figure 7 : Wing of a representative of the Pachyneuridae.
80 -- Pachyneura fasciata ZETT.
(From HENNIG, 1954, after DUDA, 1930)
This family is known only from a few locations in Europe, the Far East, and North America. The individual development and biology of these insects is still unknown [ However, at least from 1969 onwards we know something about them, see HERE in the previous document ].

Figure 8 : Wing of a representative of the Hesperinidae.
73 -- Hesperinus imbecillus LW.
(From HENNIG, 1954, after CURRAN, 1934)
Individual development and biology not known.
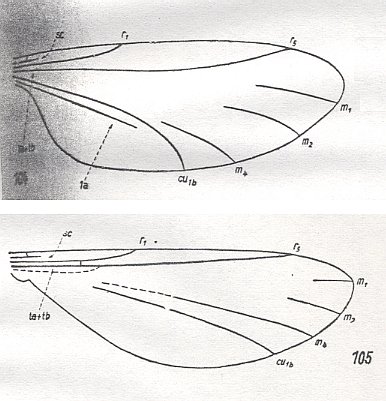
Figure 9 :
104 -- Lygistorrhina spec. (Lygistorrhinidae).
105 -- Manota spec. (Manotidae).
(From HENNIG, 1954, after HENDEL, 1938)
The Manotidae are distributed in North America and in the Palearctic Region ( Europe and Northern Asia ). Characteristic for its representatives is not only the progressed state of reduction of the wing-venation, but also the position of the antennae, the bases of which are placed near the top of the head.
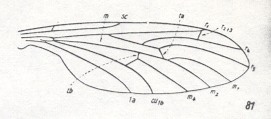
Figure 10 :
81 -- Axymyia furcata McATEE . (Axymyiidae).
(From HENNIG, 1954, after CURRAN, 1934)
The wing-base (basiala) is more or less changed. Individual development and biology unknown.
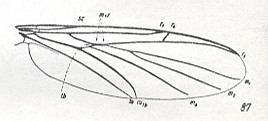
Figure 11 :
87 -- Allactoneura nigrifemur ENDERL. (Allactoneuridae).
[ This species is described by ENDERLEIN. It seems, therefore, that between 1946 and 1954 at least one more species of this family has been found, although ROHDENDORF still lists only one species -- which could be a printing error -- in his 1964 work on Diptera. However, if I have interpreted the subscript of HENNIG's table of 12 (80-91) wings correctly, that is, that "after EDWARDS 1925" also belongs to 87 (and not just to 90), then this species was already found before or in 1925, or, alternatively, MEIJERE had (1909) described it only as a genus (Allactoneura) and the species name (nigrifemur) was later given by ENDERLEIN.]
(From HENNIG, 1954, after EDWARDS, 1925)
This concludes our presentation of p. 34-38 of ROHDENDORF's 1946 account of the recent fauna of Nematocera Oligoneura which largely coincides with the infraorder Bibionomorpha as established by him in 1964.
Before we discuss this infraorder -- its system and content -- as it is presented in ROHDENDORF 1964, and add biological data from other sources, we first discuss the types of habitats of the larvae of bibionomorphs, as they are given in KRIVOSJEINA, 1969, Ontogenes i evoljutsia dwukrilich nasjekomich (in Russian). This book, however, does not discuss the taxonomic groups of the Diptera separately as to their biology, but concentrates on the Diptera as a whole. That is to say, when, say, a particular habitat is discussed its dipterous inhabitants are mentioned whether they turn out to belong to this or that dipterous taxon. A plus-point of such an approach is that we will not be seduced to see, say, the Bibionomorpha as a special group with respect to the ecology of its members. In almost all habitats that will be considered below a great many dipterous larvae, different from those of the Bibionomorphs and in addition to them, turn out to live there (although the precise ecological niches -- biochemically and microphysically defined -- of the larvae of different species must be correspondingly different).
The diversity of ways of life of Diptera is greatest in the suborder Nematocera, and of this suborder we have already discussed its two main groups, the infraorder Tipulomorpha and the infraorder Bibionomorpha (the latter only in an introductory way under the name Nematocera Oligoneura). And it is precisely here appropriate to discuss the various substrates on which the larvae of Diptera feed, while we are more or less familiar now with the families of the Nematocera and anticipate the larval feeding habits of the remaining flies, that is of the 'true flies', the Brachycera-Orthorrapha and the Brachycera-Cyclorrapha.
In our evolutionary account of the Diptera the emphasis on their ecology (mainly larval feeding habits and habitats) is justified because, as explained earlier, we think that the ecologies in any given animal group represent the driving forces of that group's evolution. The colonization of new ecological niches and new habitats by particular members of the group results in speciation, differentiation, and specialization, that is, it results in the evolution of the group. This approach is adopted not only by ROHDENDORF but also by other authors among which KRIVOSJEINA, whose's 1969 text (in Russian) on the ecology of Diptera is instructive.
We will now paraphrase some (large) parts of this text (which can be found in Chapter Two of the above mentioned book of her's).
Woodland zone
The highest abundance and diversity Diptera attain in the woodland zone. The significant amount of deposit, the large stock of rotting vegetable matter (woodland litter [podstilka], wood), the sufficient dampness during the course of the whole summer season, the absence of excessive insulation, the relatively high humidity of the air under the cover of the forest, create favourable conditions for the development of Diptera. The presence of utterly diverse biotopes provides for the possibility to species to (individually) develop, species that is, that possess different ecological potentials.
When characterizing climatic and undergrowth conditions of this or that region we must bear in mind that the abundance of Diptera in different zones is connected with microclimatological conditions of the several parts of that region and with the ecological features of the different stages of individual development.
The connection of the larvae of Diptera with the substrate.
The development of the pre-imaginal stages of Diptera proceeds in the most diverse substrates. Every solid or loose substrate, dry or damp, originating from plant or animal, rich or poor in organic residues, in short, every type of accessible substrate is capable of [letting it] being colonized by different groups of Diptera.
A large number of [species of] Diptera develops in soils of different mechanical composition.
Typically soil-borne are the larvae of Asilidae [robber-flies, Brachycera-Orthorrapha ( = roughly Asilomorpha)], Tabanidae [horse-flies, Brachycera-Orthorrapha ( = roughly Asilomorpha)], the majority of the larvae of Therevidae [Brachycera-Orthorrapha ( = roughly Asilomorpha)], Empididae [Asilomorpha], many Tipulidae [Nematocera-Tipulomorpha] and Rhagionidae [Asilomorpha].
A significant complex of Diptera is for individual development connected with disintegrating wood and the materials coming forth from it. Members of this complex can be encountered within such wood as well as under its bark, in mouldered wood [present] in hollows, or in a fermenting sap. To this complex belong the larvae of Dictenidia BRUL., Tanyptera LATR., Phoroctenia COQ., Ctenophora MEIG. (family Tipulidae) [Nematocera-Tipulomorpha], Hyperoscelididae [Nematocera-Bibionomorpha], Axymyiidae [Bibionomorpha], Pachyneuridae [Tipulomorpha or Bibionomorpha], Hesperinidae [Bibionomorpha], Clusiidae [(suborder) Brachycera-Cyclorrapha or, more or less equivalently, (infraorder) Myiomorpha], Lonchaeidae [Myiomorpha], many Bibionidae [Bibionomorpha], Sciaridae [= Lycoriidae] [Bibionomorpha], Ceratopogonidae [ = Heleidae] [Nematocera-Tipulomorpha], and, finally, Stratiomyidae [Asilomorpha].
With rotting wood are connected different fungi, where fruiting bodies and films of mold harbour their characteristic complex of Diptera ( Mycetophilidae [ = Fungivoridae] [Bibionomorpha] and related families, further, Limoniidae [Tipulomorpha], Platypezidae [Myiomorpha], and others).
Diverse is the content of the inhabitants of forest-litter [podstilka]. Here develop Bibionidae [Bibionomorpha], Tipulidae [Tipulomorpha] (mainly species of the genus Tipula L.), Cecidomyiidae [ = Itonididae] [Bibionomorpha], Ceratopogonidae [Tipulomorpha], Chironomidae [ = Tendipedidae] [Tipulomorpha], Lauxaniidae [ = Sapromyzidae] [Myiomorpha], Fanniinae [Myiomorpha-Muscidae], Lonchopteridae [Brachycera, Musidoromorpha], and others.
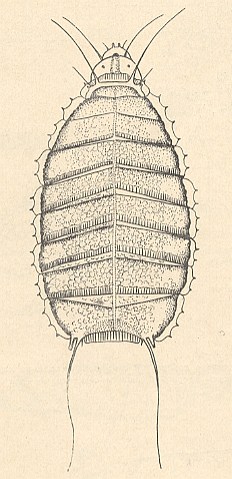
Figure 11a : General view of the larva of Lonchoptera sp. [Musidoromorpha-Lonchopteridae].
(After KRIVOSJEINA, 1969)
A large group of larvae (Cecidomyiidae [Bibionomorpha], Agromyzidae [Myiomorpha], Chloropidae [Myiomorpha], some Drosophilidae [Myiomorpha], Ephydridae [Myiomorpha] ) lives in tissues of living plants.
Character of the distribution of the larvae in their substrates.
It is necessary to succinctly dwell on regularities of distribution of larvae of Diptera in different types of substrates.
In soils [such as a forest floor] the main mass of dipterous larvae is concentrated in the litter layer [podstilka] and in the upper coarse-humus [vegetable mould] layer at 0-40 cm depth, which [latter layer] is most rich in plant roots, pervaded by tunnelings and well-aerated.
Larvae of the representatives of few families are encountered in the soil at a depth until 1 m (larvae of Asilidae [Asilomorpha], Therevidae [Asilomorpha], sometimes Tabanidae [Asilomorpha], Bombyliidae [Asilomorpha].
We can distinguish a definite zonality of the distribution of dipterous larvae in the soil. In tundras, where the accumulative horizon [where, I assume, humus will be produced and accumulate] is not developed, the high-level of groundwater results in the fact that groups of animals, which in other zones are characteristic of the [content of the] soil, concentrate in moss patches. In the zone of the northern and middle taiga the main mass of dipterous larvae is concentrated in the litter layer [podstilka] and in the upper coarse-humus layer [ The Taiga are the dense forests between tundra and steppe]. The concentration of the basic colonization in the upper layers of the soil is the rule, chiefly connected with the high-level humidity of the ground. In the southern regions of the woodland zone, but also in the woodland-steppe and steppe regions, the main animal colonization of soils changes over to within the soil horizon.
Important for the distribution of larvae in rotting wood is its state of disintegration and the present colonization of it by other invertebrates. Questions about the succession of invertebrate animals at different stages of the wood's disintegration is meticulously discussed by B.M. Mamajev.
Analysis of data about the distribution of larvae in aquatic environments indicates that the majority of species is adapted to the shoreline part of water-basins and only a few live at the bottom at larger depths, or right in the water. The main mass of dipterous larvae (Chironomidae [Tipulomorpha], Ceratopogonidae [Tipulomorpha], Tipulidae [Tipulomorpha], Limoniidae [Tipulomorpha], and others), is concentrated in a damp substrate at the bank [shore] and in deposits of mud at a depth of not more than 1 meter. Larvae of Ceratopogonidae [Tipulomorpha], for example, live at a depth down to 40 cm. The width of the shoreline band that can be colonized is significantly dependent on the angle of the slope and the degree of moistness of the substrate lying above the water level.
A comparatively small number of species can develop at the bottom of water-basins, like the above mentioned Blephariceridae [Blephariceromorpha] and Deuterophlebiidae [Deuterophlebiomorpha]. At the bottom of lakes and ponds many Chironomidae [Tipulomorpha], Ceratopogonidae [Tipulomorpha] (Sphaeromias CURT.) develop. One encounters significantly less larvae right in the water where they concentrate mainly at the surface film. Such are the larvae of Culicidae [Tipulomorpha] and some Stratiomyidae [Asilomorpha]. Sometimes larvae make use of various plants and aquatic growths floating on the surface (Ceratopogonidae [Tipulomorpha], Chironomidae [Tipulomorpha].
Nutritive connections of the larvae.
Depending on the way of feeding and the nature of the substrate to be used [consumed] we can distinguish different categories of nutritive regimes. Larvae of Diptera are able to feed on various disintegrating materials and on living tissues of plants and animals. Among dipterous larvae we can distinguish such groups as saprophags, phytophags, mycetophags, coprophags, necrophags, and zoophags.
A whole series of factors must be taken into account when studying nutritive regimes of larvae. Larvae can in different ways utilize the substrates which are at their disposal. They can eat them directly or eat the bacteria and fungi that grow on them. The way of processing the food mass can also be different dependent on the structure of the mouthparts and the particular features of digestion. Finally, one must take into account the fact that in many cases a mixed diet is observed. Many questions concerning the feeding habits of individual groups are still [1969] not clarified.
We will now successively discuss the mentioned nutritive regimes.
S a p r o p h a g o u s larvae (scavengers).
The group of (vegetable) scavengers (saprophags) includes a very wide range of organisms that feed on different decaying remains of higher plants and fungi, and rotting wood [Animals that feed on decaying animal remains are also scavengers, which we subsume, however, under the nutritive regime of necrophagous animals].
Typical saprophagous insects are the larvae of the majority of species of the genera Bibio GEOFFR. [Bibionomorpha-Bibionidae], Sciara MEIG. [Bibionomorpha-Sciaridae], Scatopse GEOFFR. [Bibionomorpha-Scatopsidae], many species of Tipula L. [Tipulomorpha-Tipulidae], for example T. transbaicalica ALEX., T. scripta MEIG., T. excisa SCHUMM., T. nubeculosa MEIG., T. rubripes SCHUMM., and others.
Saprophagous larvae live in the most diverse environments : in humus-rich soil, in woodland litter [podstilka], in deposits of mud in water-basins. They represent the dominating faunistic element in all soils with woodland litter well-expressed or with a developed accumulative humus horizon. Saprophagous larvae play an important role in the disintegration of vegetable remains. And here we must first of all point to the larvae of Bibionidae [Bibionomorpha]. The larvae of Bibio marci L. in the hornbeam forest of the Moldavian Kodr claim the second place in biomass after the earthworms, but in their role of disintegrating woodland litter they surpass the latter. Already by a number of 200 individuals pro 1 m2 the larvae of this species consumes about 15 percent of leaf fall-out, and after 100 days of activity produce about 100 g excrements. The larvae of Bibio marci L. are able to process leaves of oak, beech, hornbeam, and other leaf-carrying species. The larvae of B. clavipes MEIG., B. nigriventris HAL. and others, intensively process the litter in pine tree forests.
The significant role played by the larvae of Bibionidae [Bibionomorpha] in the intensification of the disintegration of woodland fall-out is connected with features of their ecology. The larvae are gregarious and their density in some cases reaches mythical magnitudes. The density of larvae of Bibio pomonae FABR. in the pine forests of the Archangel region [in the North of european Russia] in certain parts reaches 200-300 individuals pro 1 m2. But a most large number of larvae of Bibio GEOFFR. is observed in the woodland litter of leaf-carrying forests. Such in the litter of oak forests on the slopes of the hills in the nature reserve "Kedrovaja padj" in the autumn of 1964 on an area of 10 m2 were found 12 colonies of larvae of B. nigriclavipes HARDY & TAKAHASI. In each colony were found up to 300 individuals. In Hungary on an area of 72 m2 were found 35 colonies of B. marci L., while in each of them were found 500 up until 2000 larvae.
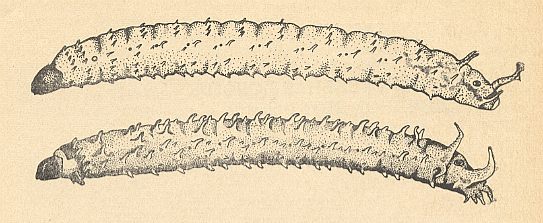
Figure 11b : Lateral view of larvae of Bibionidea [Bibionomorpha].
Top -- Bibio pomonae FABR. [Bibionomorpha-Bibionidae].
Bottom -- Plecia nigra LUND [Bibionomorpha-Pleciidae].
(After KRIVOSJEINA, 1969)
Comparatively intensively, minute larvae of Sciaridae [Bibionomorpha], also living gregariously, process woodland litter transforming it into humus. The number of larvae in the litter in certain plots in the Kadnikovski woodland of the Vologodski region reached 100 individuals pro 1 m2. As to the significance of the larvae of this family in the process of the formation of humus was also pointed to by several other authors. The larvae of Neosciara modesta STAEG. are capable of processing leaves of different sorts of trees and bushes but prefer leaves of elder, spindle tree, and elm. The larvae coming forth from 20 pairs of midges were, in the course of three weeks, able to process 30 g leaves as to their dry weight.
Figure 12 : Larvae of Ctenophora tricolor LOEW. [Tipilomorpha-Tipulidae] in pale wood of lime tree.
(After KRIVOSJEINA, 1969)
In coarse dark wood live the larvae of Symmerus annulatus MEIG. [Bibionomorpha-Ditomyiidae] and Epiphragma ocellaris L. [Tipulomorpha-Limoniidae]. See next Figure.

Figure 13 : Larva of Epiphragma ocellaris L. [Tipulomorpha-Limoniidae] in dark wood of lime tree.
(After KRIVOSJEINA, 1969)
In totally soft wood, in wood rot of hollows [in wood] or of tree-stumps develop larvae of Dictenidia bimaculata L. [Tipulomorpha-Tipulidae]. In damp dark-brown pine wood develop larvae of Sciaridae [Bibionomorpha] : Trichosia sp. (in wood of fir-tree) and Scythropochroa quercicola WINN. (in wood of silver fir). In pieces of damp dark wood or in tree-stumps live larvae of Pachyneura sp. [Tipulomorpha-Pachyneuridae]. See next Figure.

Figure 14 : Larvae of Pachyneura sp. [Tipulomorpha-Pachyneuridae].
(After KRIVOSJEINA, 1969)
The larvae of the majority of the considered groups live a solitary life. In large aggregations live the larvae of Hesperinus WALK. [Bibionomorpha-Hesperinidae] penetrating by, circular in cross section, tunnels pieces of rather solid and hard wood. See next Figure.

Figure 15 : Larvae of Hesperinus rohdendorfi KRIV. et MAM. [Bibionomorpha-Hesperinidae] in wood of elm.
(After KRIVOSJEINA, 1969)
Also living gregariously in wood are larvae of certain species of Bibionidae [Bibionomorpha], for example Bibio marci L., B. venosus MEIG., Dilophus febrilis LOEW., Pleciidae (Plecia nigra LUND.). See next Figure.

Figure 16 : Larvae of Plecia nigra LUND. [Bibionomorpha-Pleciidae] in wood of tjozenije [Cannot give the English name of the plant]. [See also Figure 11b ]
(After KRIVOSJEINA, 1969)
In some cases in logs a great abundance of larvae of Pachyneura sp. [Tipulomorpha-Pachyneuridae] is observed.

Figure 17 : Larva of Temnostoma vespiforme L. [Myiomorpha-Syrphidae].
(After KRIVOSJEINA, 1969)

Figure 18 : Larva of Mesaxymyia kerteszi DUDA [Bibionomorpha-Axymyiidae] ejecting wood rot from the tunneling.
(After KRIVOSJEINA, 1969)
The larvae themselves build the tunnelings and empty them of wood rot.
P h y t o p h a g o u s larvae
The second nutritive regime as it occurs among dipterous larvae consists in feeding on living higher (green) plants [feeding on living fungi is yet another nutritive regime observable in Dipterous larvae which will be discussed farther below].
The existing phytophagous larvae are very diverse with respect to the way of utilizing the substrate and the way of feeding.
The larvae of Tipula paludosa MEIG. and Tipula oleracea L. [Tipulomorpha-Tipulidae], the majority of the species of the genus Nephrotoma MEIG. [Tipulomorpha-Tipulidae], and some Bibionidae [Bibionomorpha], gnaw the roots of plants and in some cases cause significant damage. Thus, in coastal regions of Latvia the quantity of Tipula paludosa MEIG. and Tipula oleracea L. sometimes reached 300-400 individuals pro 1 m2, while as alarming is already reckoned a number of 30-50 individuals pro 1 m2. The density of larvae of Dilophus femoratus MEIG. [Bibionomorpha-Bibionidae], which gnaw roots of plants, in open meadow plots in the Archangel region reached 50-100 individuals pro 1 m2. The large larvae of Bibio hortulanus L. are widely known as damaging of sugar turnips and potatoes. Sometimes the damage caused by them is very significant -- up until 90 percent of damaged tubers, because in several cases the larvae are encountered in the fields in enormous numbers.
There exists a whole group of specialized phytophagous larvae which can be considered to be parasites of plants. Penetrating into the tissue of stems and leaves they mine them while they develop within the plants usually in the course of the whole vegetational period. Pupation takes place, as a rule, within the dead stems and leaves. Larvae of Chloropidae [Myiomorpha] and Opomyzidae [Myiomorpha] [in Rhodendorf 1964, p. 108 : Opomyzinae, a subfamily of the family Anthomyzidae] mine mainly stems of grasses. The larvae of Agromyzidae [Myiomorpha] develop in leaves. Larvae of Ephydridae [Myiomorpha] mine stems of various aquatic plants.
M y c e t o p h a g o u s larvae
The group of larvae representing the third type of nutritive regime includes larvae that feed on fungi. This group evermore grows in correspondence with the accumulation of known facts principally at the expense of larvae which were considered to be saprophags. Such are the majority of the larvae of gall midges (Cecidomyiidae) which are until now placed by many authors in the category of saprophagous larvae. In fact they are mycetophagous larvae feeding on threads of fungi. The larvae of certain species of Sciara MEIG., which develop in wood invested with dry rot, feed on fungi of this rot, and not on wood.
Mycetophagous larvae are principally connected with the mycelium of fungi which develop on wood. On the surface of various fallen tree trunks, on parts covered with mycelium, larvae of many species of Platypezidae [Myiomorpha] live. The flat larvae, living gregariously with 10-20 individuals as gray oval spots are well visible against the background of snow-white mold which covers the surface of logs, of various branches, laying on the forest floor. The larvae quickly devour the mycelium and thus destroying the snow-white spots of mold.
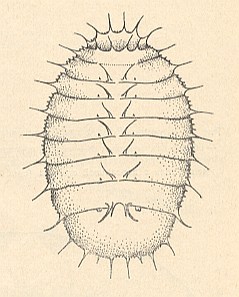
Figure 18a : General view of larva of Platypeza dorsalis MEIG. [Myiomorpha-Platypezidae].
(After KRIVOSJEINA, 1969)
In similar conditions develop larvae of Cecidomyiidae [Gall midges] [Bibionomorpha] : Peromyia monilis MAM., Dichaetia pusilla MAM. (on logs of lime tree), Acoenonia europaea MAM. (on beech). On mycelium, which overgrows thin pieces of alder lying on the forest floor, develop Brachyneura fungicola MAM. [still] (Cedidomyiidae). In the thin film of mycelium under bark on tree-stumps and logs of pine trees develop larvae of Aprionus similis MAM., Winnertzia nigripennis KIEFF., Camptomyia maxima MAM., and in tree-stumps and logs of leaf-bearing trees develop larvae of Micropteromyia ghilarovi MAM., Karschomyia aceris MAM., and larvae of other species of gall midges.
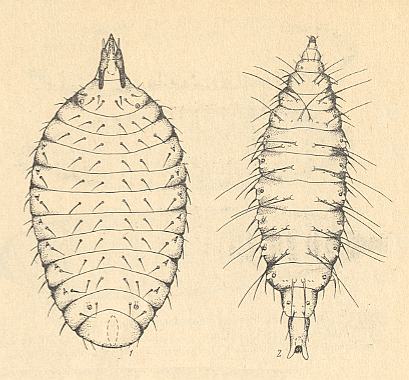
Figure 18b : Dorsal view of larvae of Cecidomyiidae [Bibionomorpha].
1 -- Rhizomyia sp.
2 -- Parepidosis ulmicorticis MAM.
We have inserted this figure to show the peculiar shape of at least these two Cecidomyiids.
(After KRIVOSJEINA, 1969)
A peculiar ecological group form the larvae that develop in the tubular hymenophores of tree-fungi. On the surface of such fungi from the underside often under a thin transparent film develop large greyish larvae of Ceroplatidae [Bibionomorpha] looking like slugs. See next Figure.

Figure 19 : Larva of Ceroplatus tipuloides BOSC. [Bibionomorpha-Ceroplatidae] on the surface of a fungus.
(After KRIVOSJEINA, 1969)
In the body of fungi large semi-transparent larvae of Metalimnobia quadrimaculata L. [Tipulomorpha-Limoniidae] are common. Often one encounters in such conditions the larvae of some species of gall midges [Bibionomorpha-Cecidomyiidae]. In the fruiting bodies of Polystictus versicolor, which [fungi] develop on birch, one may encounter the peculiar larvae of Ditomyia fasciata MEIG. [Bibionomorpha-Ditomyiidae], and Platypeza infumata HAL. [Myiomorpha-Platypezidae].
C o p r o p h a g o u s larvae
Larvae connected with the fourth nutritive regime -- the coprophags -- feed on animal (whether vertebrate or invertebrate) excrements.
N e c r o p h a g o u s larvae
Larvae connected with the fifth nutritive regime -- the necrophags -- feed on dead animals.
Concerning Coprophagia and Necrophagia
As distinct independent categories of nutritive regimes coprophagia and necrophagia are listed in KRIVOSJEINA 1969, p. 26 and 28.
Myself I [JB] think that, at least in insects, and at least under natural conditions, coprophagia and necrophagia are not independent categories of nutritive regimes. In both coprophagia and necrophagia decaying organic matter is fed upon. They stand out only because this decaying matter originates from animals, that is, not from plants. So both categories should be better subsumed under saprophagia sensu lato. They are, so to say, extensions of the basic habit of feeding on decaying vegetable matter (saprophagia sensu stricto) to feeding on decaying animal matter. Vegetable matter, it is true, differs from animal matter, but, although there still remains a difference, decaying vegetable and animal matter are becoming more similar as the process of disintegration progresses. And ecologically, with respect to finding them, excrements, dead animals, and rotting vegetable matter (such as dead trees), are similar.
Z o o p h a g o u s larvae (carnivores and internal parasites)
The last type of nutritive regime is shown by larvae that feed on living animals. For dipterous larvae this means that they either hunt and devour other invertebrate animals, mainly insect larvae, or that they parasitize them, that is, being internal parasites. The difference from feeding on, and parasitizing in, living plants lies, of course, in the biochemical nature of the food (because in both cases it is fresh), calling for a different feeding apparatus and a different digestion. Also ecologically they differ : finding the right food plant, and hunting or parasitizing the right animal, demand different adaptations (morphological and behavioral).
Clearly expressed carnivores are the larvae of the majority of families of the Brachycera Orthorrapha [largely belonging to the infraorder Asilomorpha, such as horse-flies, robber-flies, and the like, that is, true flies, but not the 'truest' flies (such as fruitflies or the house-fly and its allies)], among which many or all species live at the expense of different invertebrates.
Larvae of Rhagionidae [Asilomorpha] feed mainly on earthworms, the larvae of Tabanidae [Asilomorpha] and Therevidae [Asilomorpha] feed mainly on larvae of insects. Actively attacking earthworms are larvae of Rhagio tringarius L. [Asilomorpha-Rhagionidae] See next Figure.
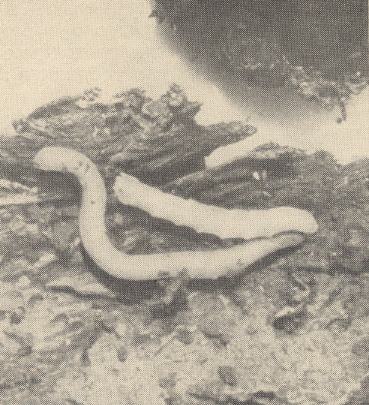
Figure 20 : Larva of Rhagio tringarius L. [Asilomorpha-Rhagionidae], attacking an earthworm.
(After KRIVOSJEINA, 1969)
Not less energetic behave larvae of Therevidae [Asilomorpha], which have their habitat in soils of light mechanical content or in wood rot. Very agile, they easily move in the soil and change places in order to find food over significant distances. Such a way of life is also characteristic of larvae of Tabanidae [Asilomorpha].

Figure 21 : Larva of Laphria flava L. [Asilomorpha-Asilidae].
(After KRIVOSJEINA, 1969)
In laboratory conditions they accept as food larvae of many other insects [meaning that they are, at least physiologically, not specialized in eating larvae of long-horned beetles].

Figure 22 : Larva of Xylophagus ater MEIG. [Asilomorpha-Xylophagidae].
(After KRIVOSJEINA, 1969)
They eagerly eat larvae of bark-eaters, namely of long-horned beetles, of crane-flies, of gall midges, of fungus gnats, and even larvae of click beetles which possess a solid integument. Very rarely, but nevertheless recorded, are cases of feeding on earthworms and cases of cannibalism. The larvae pierce the integument of the prey, discharge digestive saps into the body, and then suck the content such that only an almost empty skin is left. The larvae of some species, for example Xylophagus cinctus de GEER. [Asilomorpha-Xylophagidae] are able to feed on remains of dead insects.
A special form of zoophagia can be considered internal parasitism, and also is the habit of certain dipterous larvae. Even [at least] a whole family, the Tachinidae, is such that their larvae live as internal parasites in other insects, mainly caterpillars. It is clear that such endoparasitic groups (but the same can be said of ectoparasites which also occur in Diptera) as such represent a special ecological type, comparable to that of the parasitic wasps (Hymenoptera). In KRIVOSJEINA, 1969, little attention is given to parasitic Diptera (the scope of the book is evidently already broad enough!). For the family Tachinidae a lot is expounded in HERTING, B., 1960, Biologie der westpaläarktischen Raupenfliegen, Dipt., Tachinidae.
Let us continue with KRIVOSJEINA, 1969, p. 30.
Larvae of parasitic Diptera as a rule parasitize various invertebrates. The host, which is infested by the parasite, keeps on developing. It does not show features of strain. Pupation and flying-off of the parasites usually coincide with the conclusion of the infested larva's development. The parasitic larvae find themselves in the body of the host during the whole period of the parasite's development, and leave it before pupation. Only in some cases, where massive infestation has taken place, for instance in the infestation of earthworms by larvae of Onesia ROB.-DESV. [Myiomorpha-Calliphoridae], the latter may migrate out of one worm into another. This happens in the case of death of the earthworms which were the initial hosts.
There are larvae of Diptera, Pipunculidae [Myiomorpha], that develop in cicadas, Conopidae [Myiomorpha] in bumblebees, wasps, and bees, Pyrgotidae [Myiomorpha] in the body of scarabeid beetles, Sarcophagidae [Myiomorpha] in the body of earthworms, and Calliphoridae and Tachinidae in isopods, earthworms, and in various insects.
Insofar as earthworms are known to actively process the soil, the question concerning larvae as parasites of worms constantly drew the attention of investigators. Thus, the larva of Pollenia rudis FABR. [Myiomorpha-Calliphoridae], develops in the body of the worm from its first to last instar. Presently P. rudis FABR. is recorded as parasite of Allolobophora chlorotica SAV., Eisenia rosea SAV., Lumbricus terrestris L., A. caliginosa SAV. Also parasitizing earthworms are Sarcophaga carnaria L. [Myiomorpha-Sarcophagidae], Onesia ROB.-DESV. [Myiomorpha-Calliphoridae].
After having given this list of nutritive regimes as they are encountered in larvae of Diptera, KRIVOSJEINA (p. 31) concludes with some general observations.
The listed trophic groups [categories] do not cover all diversity of the nutritive connections that are observed in Diptera. Much remains unclear. The biology of a series of groups is still totally unknown. With respect to the ecology of certain individual groups there does not as yet exist one definite opinion. Some reckon, for instance, the soil-dwelling larvae of Asilidae [Asilomorpha] as being phytophagous, others as predators. The larvae of Stratiomyidae [Asilomorpha] are considered to be predators by some authors. Unclear is the nature of feeding of many larvae of Empididae [Asilomorpha] and others.
Resolving the question concerning the nature of feeding of this or that group is often hampered by the fact that nutritive connections usually do not present themselves in such a pure way that one can, without errors, determine their category. The larvae of many species of Stratiomyidae [Asilomorpha] develop under bark (for example Pachygaster minutissima ZETT., P. orbitalis WAHLB., and others), are mainly saprophagous, they feed on various minute particles of wood, [namely] wood rot. But it can often be observed that they concentrate around dead bodies of various insects and eat them. The same can be observed also in the aggregations of Solva WALK. [Asilomorpha-Stratiomyidae], in many Lonchaeidae [Myiomorpha] and Pallopteridae [Myiomorpha, see ABOVE ]. In laboratory conditions the larvae-xylophags of Tanyptera LATR. [Tipulomorpha-Tipulidae] and Ctenophora MEIG. [Tipulomorpha-Tipulidae] pupate sooner when they are offered as food not only pure wood but also protein-containing food in the form of dead bodies of insects. The same phenomena can be observed not only in Diptera. Among the click beetles Corymbites LATR. and Selatosomus STEPH. there is a group of omnivores with strongly expressed phytophagia of the larvae, while for a normal development of the latter food originating from animals is also required.
Sometimes, in cases of the absence of appropriate objects [to feed on], cannibalism is observed, which is intrinsic to larvae of many species of Stratiomyidae [Asilomorpha], Rhagionidae [Asilomorpha], Xylophagidae [Asilomorpha], Lonchaeidae [Myiomorpha], and others. It is possible that such behavior is clarified by the fact that it is necessary for larvae developing in a meagrely nutritive substrate for their final development to obtain some quantity of protein-containing food.
The dual character of feeding is observed in many saprophags. In some cases typical saprophags are, in conditions of insufficient wetness, able to feed on living tissues of plants (for example Bibio hortulanus L.). Similar phenomena are characteristic to many groups of insects.
There exists a definite dependency between the systematical position of [any] given Diptera and the way of feeding of their larvae. The larvae of the majority of families of Nematocera [midges, gnats, mosquitoes, crane-flies, etc] are saprophagous. Cases of being carnivorous are rare (some species of Cecidomyiidae [Bibionomorpha], Ceratopogonidae [Tipulomorpha], Limoniidae [Tipulomorpha] ). A comparatively small group form the phytophagous Diptera larvae, namely some Dilophus MEIG. [Bibionomorpha-Bibionidae], Tipula L. [Tipulomorpha-Tipulidae], Nephrotoma MEIG. [Tipulomorpha-Tipulidae], and many species of Cecidomyiidae [Bibionomorpha].
Very compact regarding its nutritive connections is the group of Brachycera-Orthorrapha [horse-flies, robberflies, and the like]. The larvae of the majority of families are clearly expressed zoophags (predators or parasites). Literally unique are the saprophags among them (larvae of some species of Stratiomyidae [Asilomorpha], Tabanidae [Asilomorpha], and Rhagionidae [Asilomorpha].
The larvae of Cyclorrapha [hover-flies, fruit flies, house-flies, bluebottles, and the like] are connected with diverse habitats, but coprophags and zoophags predominate. Phytophags are a little less represented among them, and only an insignificant part of them are saprophags.
With all this we conclude the part concerning feeding habits and habitats of dipterous larvae, as it is taken from KRIVOSJEINA, 1969, p. 14-32. We will use this account when separately considering dipterous families. The Diptera Nematocera are in this respect the most diverse and will offer much to understand the evolutionary process as driven by the colonization of new habitats and by the occupation of new ecological niches in an existing or new habitat.
Of the Nematocera the infraorder Bibionomorpha is very promising in this respect because of the presence of the very diverse and special family of gall midges (Cecidomiidae) and of a number of relict families. But the infraorder Tipulomorpha is also interesting, especially as a result of the presence of the extensive and diverse family Limoniidae which shows many evolutionary 'excursions' to most diverse larval habitats, and of the presence of the very ancient family Tanyderidae.
In the next document we will consider again, but now more extensively, the infraorder Bibionomorpha, basically following ROHDENDORF's 1964 book on the historical development of Diptera. We will discuss the superfamilies -- one after another -- of this infraorder as they are presented in ROHDENDORF, with additions and alterations where necessary.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part VI (Sequel to Diptera-Bibionomorpha).
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V