
Organic Evolution in terms of the Implicate and Explicate Orders.
Part X :
The evolutionary diversification in the Order Diptera.
B. The Infraorder Bibionomorpha (Diptera) (Sequel).
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Back to Homepage
Superfamily Scatopsidea
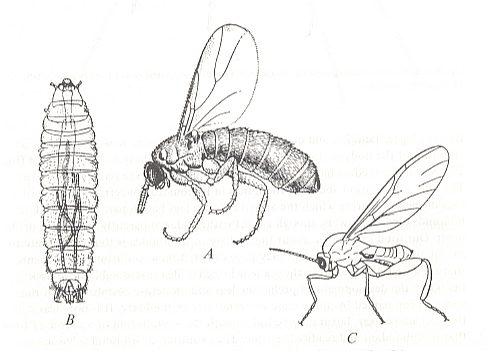
Figure 1 : Scatopsidea
A -- Scatopse fuscipes MEIGEN ( Scatopsidae, female), general view.
B -- The same, larva, top view.
C -- Canthyloscelis antennata EDWARDS ( Hyperoscelididae, male), general view.
(From ROHDENDORF, 1964, A, B, after BURAKOVA, 1931, C after EDWARDS, 1922)
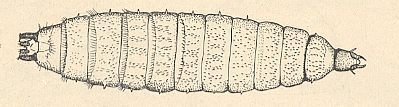
Figure 2 : Larva of Scatopse subnitens VERRALL ( Scatopsidae), top view.
(After KRIVOSJEINA, 1969)
The superfamily Scatopsidea is as old as the Bibionidea : The oldest fossil representatives of the group are, together with bibionideans, found in the middle Jurassic fauna of Karatau [The age of the Karatau fauna is maybe a little bit younger : upper Jurassic] (Family Protoscatopsidae).
In the recent fauna the Scatopsidea are [taxonomically] a small group and consists of only three families from which only one, the Scatopsidae, includes about 130 species. The two other families are clearly expressed relict groups with a few single species (Hyperoscelididae -- 10 species, Synneuronidae -- 1 species). [Much about the Scatopsidea was already considered in the previous document.]
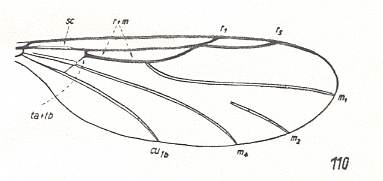
Figure 3 : Wing-venation of Synneuron annulipes LUNDSTR. ( Corynoscelidae [ or: superfamily Scatopsidea, family Synneuronidae] ). In addition to the coalescence of the anterior medial vein with the Radial Sector we see here a coalescence, at one point only, of the Radial Sector and the Radius.
(After DUDA, 1928, from HENNIG, 1954)
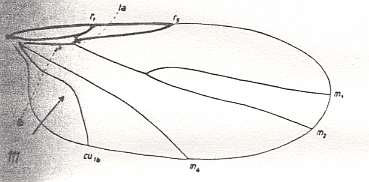
Figure 4 : Wing-venation of Scatopse fuscipes MEIG. ( Scatopsidae).
(After HENNIG, 1954)
The chief features of the Scatopsidea (see Figures 1-4) consist in the refinement of the sense organs of the head (enlargement of the compound eyes, the reduction of the antennae to fewer stout segments), small or minute size of the shortened body, the development of highly costalized wings, and strong prehensile legs. The predatory larvae live in any kind of terrestrial, but moist substrates. In the imago the thoracic section is large with greatly enlarged terga [dorsal shields], and the sternite [ventral shield] of the prothorax is not bridged to the pleura [lateral shields]. The individual development of the Scatopsidea goes differently in the representatives of the known [different] families, apparently including the basic determining features that led to their origin. The most primitive structural type of developmental stages are observed in the representatives of the family Scatopsidae, in which metapneustic larvae (see Figure 6 (4) in Part VIII ) of the FIRST stage are transformed, through amphipneustic larvae [(3) of same Figure] of the SECOND stage, into peripneustic larvae [(2) of same Figure] of the THIRD and FOURTH stages. The larva has chewing mouthparts and the posterior spiracles lie at the end of processes (Figure 1(B) ). The pupa carries prothoracic spiracles.
Considering the first [=original] conflicts in the historical development of the Scatopsidea one should perhaps note the following important elements : The predatory nature of the larvae, which live in moist temporary terrestrial substrates (any kind of rotting matter, excrements of vertebrates, vegetable remains). Living in such conditions and the predatory nature of the larvae at the expense of other minute insects (for example larvae of other diptera), which conditions and features were adopted by the scatopsideans already very early on, determined the development of the lifting type of flight apparatus and the refinement of the sense organs. The body-size did not increase, and also the feeding of the winged phase was not refined, they remained vegetarians.
The [historically] first Scatopsidea are, up to now, not known, although almost certainly one can say that they belong to ancient fungivorids.
Summarizing all that has been said about the history of this superfamily as a whole, one can say that the refinement of the sense organs, of the wings and legs, the oligomerization [decrease of the number of segments] of the body of the winged form and the refinement of the nature of feeding of the larva, together with the limitation of feeding of the adult, while the latter becoming smaller, were the determining features reflecting the resolution of the conflict which resulted in the formation of the Scatopsidea.
Much more precisely we can clarify the next stage of the history of the Scatopsidea, namely the formation of two distinct families in them, the Scatopsidae and the Hyperoscelididae, of which the characteristics are known. The development of these two groups was determined by different solutions of the chief conflict in their history which [history] includes the necessity to find conditions well-suited to their larvae (abundance of food and protection). The first forms of the family Scatopsidae appeared as a result of the solution of this conflict by way of the refinement of flight, of the sense organs, and the decrease of size of the winged phase, which made it possible to find and colonize the restricted substrates necessary for individual development, substrates in which their predatory larvae might be able to live. The larval phase was almost not refined and acquired only this protective device : a primitive puparium was worked out, which is that in the transformation of the larva into a pupa the last larval skin is not discarded.
Something quite different took place in the history of the Hyperoscelididae. The solution of the conflict which determined the formation of these scatopsoids went into the direction of the refinement of feeding in the larval phase, namely the working-out of extra-intestinal digestion. The flight-apparatus was not improved, and there also was not worked out the formation of a puparium, neither a decrease of body size.
In the previous document we considered the family Canthyloscelididae. This family apparently more or less coincides with the family "Hyperoscelididae" or with the "Synneuronidae" + "Hyperoscelididae" or, possibly with the family "Corynoscelididae" (Corynoscelidae). In any case they belong to the superfamily Scatopsidea. After 1964 (date of publication of ROHDENDORF's book) some more came to be known about them. These new data -- larval features -- have been given by KRIVOSJEINA, 1969 (pp. 239-240), as being data concerning the family "Hyperoscelididae". We will produce these data here :
Family Hyperoscelididae
This family includes the representatives of the genera Synneuron LUND., Hyperoscelis HARDY et NAGAT., and Canthyoloscelis EDW. [The genus Corynoscelis is not mentioned. Because of synonymity? HENNIG, 1954, p. 312, mentions the genera Corynoscelis, Canthyloscelis, and Synneuron as belonging to the family Corynoscelidae (without explicitly saying that these are all the genera of this family), see Figure 3, above and the next two Figures].
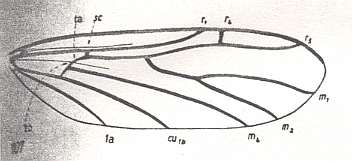
Figure 5 : Wing-venation of Corynoscelis eximia BOH. ( Corynoscelidae).
(After DUDA, 1928, from HENNIG, 1954)
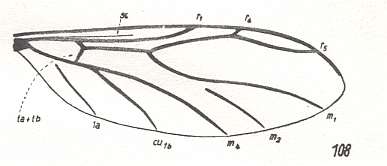
Figure 5a : Wing-venation of Canthyloscelis antennata EDW. ( Corynoscelidae).
(After DUDA, 1928, from HENNIG, 1954)
The representatives of the family were included into the family Scatopsidae as an independent subfamily (Duda, 1928). The revision of the subfamily and its separation as an independent family was carried out comparatively recently (Tollet, 1959). In the book of Rohdendorf (1964) three individual families are distinguished within this group of diptera -- Hyperoscelididae, Synneuronidae, and Scatopsidae. We consider the family Hyperoscelididae as consisting of two subfamilies Synneuroninae and Hyperoscelidinae. The first is represented by one single genus Synneuron LUND., while the other is represented by two genera, namely Hyperoscelis HARDY et NAGAT. and Canthyloscelis EDW.
Until recently the biology of the group was very little studied. The early developmental stages of only Canthyloscelis antennata EDW. [see Figure 1C, above ] from New Zealand were known (Tonnoir, 1927). The acquiring of new data concerning this group (Krivosjeina & Mamajev, 1967) makes it possible to more precisely analyze the mutual connection of the genera Scatopse GEOFFR., Synneuron LUND., Hyperoscelis HARDY et NAGAT., and Canthyloscelis EDW.
The larvae of the genus Scatopse GEOFFR. (see Figure 2, above ) (the biology of the other genera of this family are, until now, unknown) have a well-developed head-capsule (see HERE for a Figure ), an elongate upper lip, masive elongate mandibles, and plate-like maxillae. The body is dorso-ventrally flattened, lighly narrowed at the anterior and posterior ends. The last (ninth) body-segment is small, shifted a little ventrally. Body-segments dorsally and laterally covered with long spinelets, hairs and spines, usually gathered in groups. The segments ventrally with series of minute spinelets and hairs. The last segment is small, rounded, and at its end with two long sclerotized projections. The tracheal system is peripneustic (see Figure 6 (2) in Part VIII ), and the spiracles are in the form of small sclerotized bumps which on their surface carry up to 10 openings. The last pair of spiracles in the form of large sclerotized bumps, sometimes very long, equal in length to the projections of the last body-segment. The larvae develop in various rotting vegetable substances, in dung, wood-rot, under rotting bark of trees, etc.
The larvae of the remaining three genera (Synneuron, Hyperoscelis, Canthyloscelis) do not have anything in common with the larvae of Scatopsidae. See next Figure.
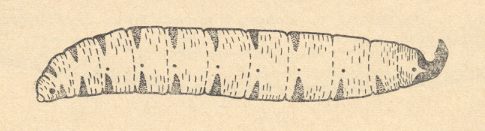
Figure 6 : Larva of Hyperoscelis eximia BOHEM ( Hyperoscelididae), lateral view. Compare with Figure 2, above .
(After KRIVOSJEINA, 1969)
The larvae [of Synneuron, Hyperoscelis, and Canthyloscelis] are acephalous [that is, they lack a clearly distinct head], usually with a dark gut that shines through, and the end of the body with two big sclerotized and dorsally bent-up hooklets. The head-capsule is reduced, but there is still a well-developed head-segment. The body is bald, without any outgrowths or spinelets. It is cylindrical, not flattened, lightly narrowed towards the ends, consisting of a small head-, three thoracic, and nine abdominal, segments. The first thoracic segment is secondarily subdivided. The majority of the body-segments dorsally and ventrally with patches formed by minute spinelets. The spiracles are placed at the prothorax, at the first seven and at the ninth abdominal segments. Around the prothoracic spiracles there is an elongate-oval large plate. A similar plate is also present around the last pair of spiracles. The larvae develop in advanced stages of disintegrating wood in which mycelium has penetrated.
The study of the larval forms has indicated that both groups are, undoubtedly, independent branches which have separated [that is, which have 'individualized' themselves] already a long time ago, and [thus] the Hyperoscelididae and Scatopsidae can be determined as being independent families. The features of the imago just confirm such a point of view. The structure of the head, antennae, eyes, and the wing-venation, clearly let the representatives of the Hyperoscelididae be distinct from those of the Scatopsidae. [As to the wing-venation being different, we can add, following HENNIG, 1954, p. 312 : The wing-venation of the Scatopsidae differs from that of the (basic plan of the) Corynoscelidae (which here must be seen as synonymous with the Hyperoscelididae) by the following features : Radial Sector consists of just one single vein. The anterior branch, present in (the basic plan of the) Corynoscelidae is absent, see Figure 5, above and Figure 4, above. Further, the bifurcation point of the Cubitus (in the Figures the veins CU1b and M4 ) is markedly shifted towards the wing-base, resulting in the cross-vein tb being positioned markedly obliquely (in fact parallel to the longitudial axis of the wing) and being shifted almost into the position of the original [but reduced] basal part of M (see the mentioned Figures). Finally, the first anal vein (1a) is absent.]
* * *
Superfamily Rhyphidea
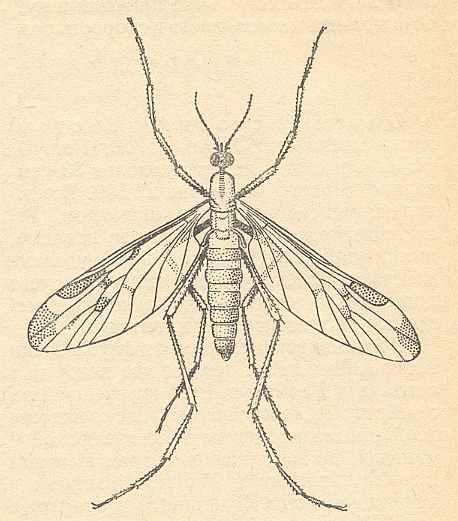
Figure 7 : Rhyphidea
Rhyphus fenestralis SCOPOLI ( Rhyphidae), female.
(After LINDNER, 1930, from ROHDENDORF, 1964)
The superfamily Rhyphidea [ = Phryneidea] is one of the most ancient groups of Diptera, apparently represented already in the upper Triassic fauna [See HERE ]. Today [that is, in the recent fauna] the superfamily consists of three monotypic families, which have a clearly expressed relict nature, and includes about 100 species ( Rhyphidae -- 66 species, Olbiogastridae -- 31 species, Cramptonomyiidae -- one species).
These diptera are of great theoretical interest in being nearest to the original ancestral groups of the infraorder Asilomorpha and its derivatives and hence of many other, younger groups of the order ( Myiomorpha and some parasitic diptera).
[The next Figures depict the basiala and wing-venation of representatives of the superfamily Rhyphidea] :
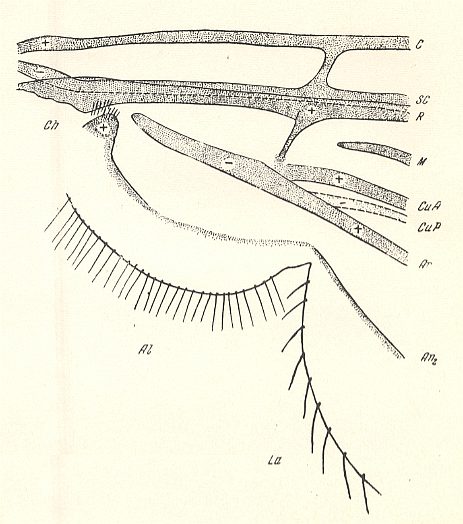
Figure 8 : Basiala of wing of Phryne [ = Rhyphus] sp. ( Phryneidae [ = Rhyphidae] ). Schematic.
Abbreviations of veins as in earlier Figures. Above the second anal vein (An2) lies the first anal vein (An1). CuP is a fold-like structure present in the basic plan of dipterous wing-venation. Al = alula, Ch = chaetarium.
(After ROHDENDORF, 1946 )

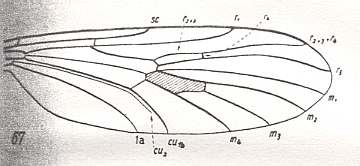
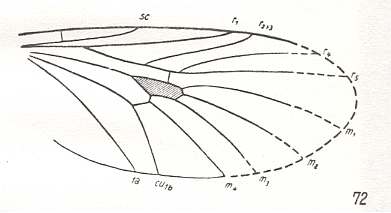
Figure 9 : Rhyphidea
Top image : Wing-venation of Phryne fuscipennis MACQ. ( Rhyphidae [=Phryneidae] ). (After HENNIG, 1954).
Middle image : Wing-venation of Cramptonomyia spenceri ALEX. ( Cramptonomyiidae). (From HENNIG, 1954, after ALEXANDER, 1931).
Bottom image : Wing-venation of Protorhyphus stigmaticus HANDL., from the upper Liassic of Mecklenburg. ( Protorhyphidae). (From HENNIG, 1954, after HANDLIRSCH, 1938).
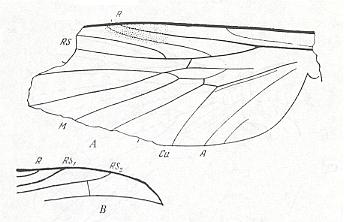
Figure 10 : Archirhyphus asiaticus ROHD. ( Protorhyphidae), holotype, Coll. PIN No. 2452/334. Middle Jurassic, Karatau (Southern Kazachstan).
A -- Left wing. B -- Tip of right wing.
(After ROHDENDORF, 1964)
The basic features of the Rhyphidea include the presence of a great many "primitive" features of the organization of the winged phase and of the mouthparts of the larva.
The head of the imago with long antennae (see Figure 7, above ), and 4-segmented palpi. Eyes of moderate size, and a proboscis capable of sucking vegetable liquid.
The convex thoracic part carries broad, little-elongated wings and long legs of the thin type suitable for active running. The sternite of the prothorax almost completely detached from the pleural sclerites.
The wing-venation (see Figure 9, above ) is very complete, almost without any shifts or reductions of individual veins.
The basiala (see Figure 8, above ) is well expressed and possesses a strong phragma, free and complex 'handles' [=basal ends] of the chief veins (consisting of the cubital and first anal veins), reduced second anal vein, its basal part taking part in the formation of a chaetarium, and, lastly, with a well expressed alula.
The abdomen is cylindrical, consisting of 8 free segments.
The larvae with a sclerotized head and with a complete chewing mouth-apparatus (with a prosteca). They are amphipneustic (see Figure 6(3) in Part VIII ), elongate, with subdivided body-segments (the anterior part of the segment is distinct). See next Figure.
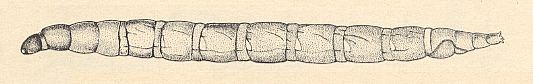
Figure 11 : Larva of Sylvicola fenestralis SCOP. ( Anisopodidae [ = Rhyphidae, Phryneidae] ).
(After KRIVOSJEINA, 1969)
The larvae live in the most diverse moist habitats and are polyphagous.
The pupae are elongate with short prothoracic spiracles. Development in the known species of the genus Sylvicola (syn. Rhyphus, Phryne) is very rapid.
The distinguishing features of the recent families of the Rhyphidea are still little known and do not make possible to say something about the historical development of the superfamily. Only a discussion of the characteristic features of the group as a whole, chiefly based on our kowledge of the main group, the Rhyphidae, may partly clarify the essence of the conflicts that determined its [that is, the superfamily's] origin. The chief features, to which we must direct our attention, probably must be, on the one hand, the rapidity of individual development and the polyphagy of the larva, and, on the other, the development of large wings with a well-distinct basiala, long thin legs, suitable for running, and the feeding in the winged phase on vegetable juices. These features clarify the reasons for the larvae to live in extensive moist substrates rich in decaying vegetable substances, substrates that, although widely distributed, are relatively less permanent (shoreline of streams or the soil of temporary water-basins in tropical forests, etc.). Rapid development and polyphagia of the larva together with the development of moderate locomotion of the winged phase (in conditions of a moist, shaded habitat), undoubtedly have resulted in the formation of the ability of a low-speed lifting flight (moderate wing-beat frequency and the absence of a need to costalize the wing-venation) and in the preservation of running-ability -- such, apparently, were the ways in resolving the conflicts in the historical development of the Rhyphidea. We must add that the supposed ways of [historical] development of the Rhyphidea certainly might be connected by [/with] conditions of high temperatures and moisture in the absence of the influence of direct sunlight, in short, in the situation of tropical forests, where, we may legitimately suppose, the origin of this superfamily has taken place.
In considering the determining features warding off the conflicts which conditioned the appearance of the Rhyphidea, we may, with great definiteness, note that they consist in the acceleration of development, in polyphagia of the larva, and in refinement of locomotion [running] of the winged phase.
This analysis of the features of the historical development of the Rhyphidea does not sufficiently characterize the whole process. Remarkable is the fact that relatively few aspects of the insect's organization are representing "new" features in resolving the conflict. In the history of the group a whole series of organs has not been transformed or did so very little. Such a nature of the history of the Rhyphidea is not simply unexpected, but intrinsically connected with a great many organisms which carry the name of "primitive relicts". Not further following this process of the formation of relict forms, but returning to the concrete consideration of the history of the Rhyphidea, we, in concluding, must note that although we have tried to uncover the actual conflicts in the history of these flies in noting the fully determined refinements of development (polyphagia of the larva, accelleration of development, and mobility of the winged insect), we in essence by far haven't yet uncovered the real causes and ways of the formation of these phenomena. What precisely caused the polyphagia and rapidity of [individual] development in Rhyphidea remains unclear.
* * *
With the superfamily Rhyphidea we have now come to an end of our exposition of the infraorder Bibionomorpha.
What comes next is the exposition of a few relict infraorders, namely the Nymphomyiomorpha, the Deuterophlebiomorpha, and the Blephariceromorpha. Their interpretation as "infraorders" (instead of mere families that belong to one or another of the known infraorders) is certainly not generally accepted. Such disagreement is often caused by the fact that the term "kinship" (expressing some relations between organisms) can be used in various different ways.
e-mail : ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part XI.
Back to Homepage
Back to Contents
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V
Back to Logic Part I
Back to Logic Part II
Back to Logic Part III
Back to Logic Part IV
Back to Logic Part V
Back to Logic Part VI
Back to Logic Part VII
Back to Nomological Part I
Back to Nomological Part II
Back to Nomological Part III
Back to Nomological Part IV
Back to Evolutionary Part I
Back to Evolutionary Part II
Back to Evolutionary Part III
Back to Evolutionary Part IV
Back to Evolutionary Part V
Back to Evolutionary Part VI
Back to Evolutionary Part VII
Back to Evolutionary Part VIII
Back to Evolutionary Part IX