
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) After having -- in the previous document -- dealt with the superfamilies Bolitophilidea and Cecidomyiidea, we will now discuss the superfamily Bibionidea [still] of the infraorder Bibionomorpha.
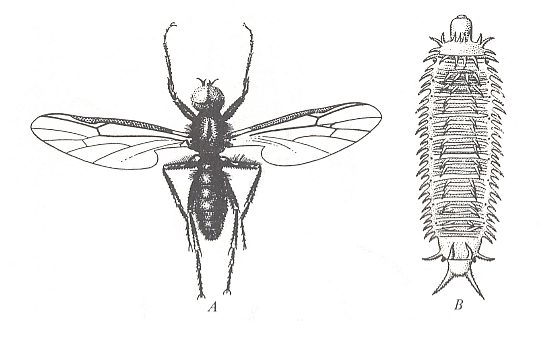
Figure 1 : Bibionidea.
A -- Bibio marci L. Bibionidae, male. General view. Body-length 10 mm.
B -- Penthetria holosericea MEIGEN. Penthetriidae, general view of larva, top view. Length 12 mm.
( From ROHDENDORF, 1964, A, after LINDNER, 1928, B, after HENNIG, 1948-1952)
The superfamily Bibionidea, which is a bit younger group of diptera as compared to the fungivoroids, has its up-to-now earliest representatives in lower Jurassic faunae. Today -- that is, in the recent era -- the Bibionidea include three families with an overall number of species of about 500, of wich more than three quarters belong to the largest family Bibionidae (about 390 species). 100 species belong to the family Penthetriidae [more or less coinciding with Pleciidae], while the family Hesperinidae has only two species [ROHDENDORF, 1964] [According to KRIVOSJEINA, 1969, there are six species, all belonging to the genus Hesperinus WALK.]. This family is a clearly expressed relict group.

Figure 1a : Bibionidea.
A -- Bibio marci L., March-fly, Bibionidae. Feeding from a flower.
(After OLDROYD, 1964)
The features of individual development are known only for the two main families of the Bibionidea and do not include one or another newly developed trait, apart from the armament of the body of the larvae, most distinct in the Penthetriidae (see Figure 1B ).
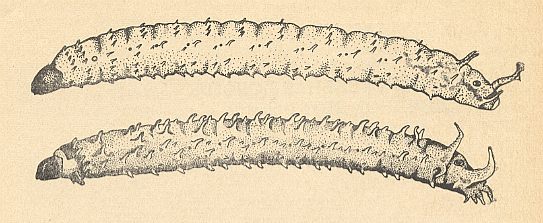
Figure 2 : Lateral view of larvae of Bibionidea [Bibionomorpha].
Top -- Bibio pomonae FABR. [Bibionomorpha-Bibionidae].
Bottom -- Plecia nigra LUND [Bibionomorpha-Pleciidae].
For the larva of Penthetria see Figure 1 B, above .
(After KRIVOSJEINA, 1969)
The nature of reproduction in the Bibionidea is also little known. There only exists information about their laying the eggs in the soil, in which act the armoured fore-legs apparently play a role. The winged form of the representatives of the family Bibionidae feeds from flowers. Those of the family Penthetriidae aparently do not feed. We must stress the insufficiency of our knowledge of the features of the living-conditions of the Bibionidea. All what is known to us about the way of life and organization of these insects, does not allow to draw whatever determined conclusions concerning the character of the conflicts which have determined the origin of the group as a whole. Apparently, the primary determining traits, which were responsible for the branching-off of the first Bibionidea from ancient Fungivoridea (which happened, probably, in triassic times), were the increase of the absolute size of the body, which took place alongside the development of prehensile legs and a moderate costalization of the wings. The conditions which contributed to the increase of body-size are not clear. Perhaps the plentifulness of easily assimilatable food was the cause of this process. This chief pathway of the historical development of the Bibionidea led to the appearance of the jurassic groups Eopleciidae ( Figure 3 ), Mesopleciidae ( Figure 4 ), and Paraxymyiidae ( Figure 5 ), and further to the caenozoic Hesperinidae and Penthetriidae.
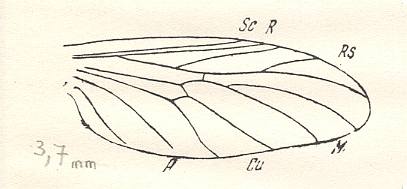
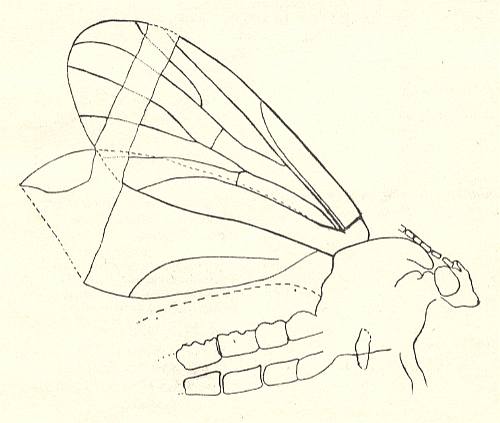
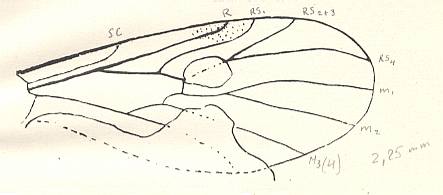
Figure 5 : Wing of Paraxymyia quadriradialis ROHD..
Jurassic of Karatau [southern Kazachstan]. The village Halkino. Specimen PIN No. 2472/584 [as given in ROHDENDORF's figure 50 of the wing, and PIN no. 2452/584, as given in ROHDENDORF's figure 49 of the whole insect].
Length of insect with inflated abdomen 3.75 mm. Actual length about 3.25 mm.
Length of wing 2.25 mm. Male.
(After ROHDENDORF, 1946)
Soon after the branching-off of the original forms of the superfamily, the origin of the still very small Protobibionidae with sharply costalized wings took place, which, in turn, later led to the formation of the caenozoic Bibionidae. Not excluded, however, is the possibility that these early costalized bibionoids were not phylogenetically connected with the Bibionidae, that is, that they were just a parallel similar group that originated significantly earlier. The essence of conflicts which determined the origin of the costalized Bibionidae remains unclear. Regarding this we have insufficient knowledge which does not guarantee an all-sided clarification of the living-conditions of these insects. The armoured and very strong prehensile legs with well-developed appendages of the pretarsus, the sharply expressed sexual dimorphism with respect to the structure of the head and the color of the body, the lifting costalized wings, and, finally, the 'simple' larvae, all point to the peculiar nature of the conflict. But to uncover the latter is difficult.
Next we reproduce an account about the family Bibionidae as given by OLDROYD, 1964 :
The family Bibionidae, though related to the families Mycetophilidae and Sciaridae [we can say, they are Bibionidea that are related to the Fungivoridea], has become more thoroughly terrestrial, both as adults and as larvae. The adult flies are bigger and heavier, and some have a distinctly armoured appearance, complete with formidable spurs and spines on the legs and head (For a general view of the insect see Figure 1(A) and Figure 1a ). The adults are predominantly flower-lovers, and hoverers in the sun, both habits that are shared by a great many flies. They are called 'March-flies' in North America, and Bibio marci has been known in Europe as 'St Mark's fly', both names associating the Bibionids with spring, and a return to sunshine and flowers. Dilophus febrilis and other small Bibionidae swarm over early flowering shrubs.
Male Bibionids have an enlargement of the upper facets of the eyes, possibly being associated with the habit of dancing in swarms. Bibionids certainly do this.
The larvae of Bibionids are among the most terrestrial, and probably the most primitive, of any flies. They have strong chewing mouthparts, and eat most kinds of vegetable food. In the soil they are often gregarious, and may be found as a mass of grubs in a pocket in the soil. They also breed in dung, and in caves.
As is appropriate to their terrestrial habits, Bibionid larvae have a very complete set of open spiracles, two thoracic pairs and eight on the abdomen.
The families Pleciidae, Hesperinidae, Pachyneuridae, and Axymyiidae
Within the complex of bibionoid and related flies (midges) the systematic status and position of the genera Plecia, Penthetria, Pachyneura, Hesperinus, Axymyia (and also Protaxymyia, Mesaxymyia) was (up to 1964) not so clear. However, new data provided by KRIVOSJEINA, 1969, might throw new light on the problem. These data are about the morphology of the respective larvae.
However, before we extract these data (together with imaginal features, such as the wings, antennae, legs, etc.) we will say something about the general features of larval morphology. For a complete understanding they will not be totally sufficient, but will ease the reading of the ensuing text (of Krivosjeina, pp 229) on these families and associated genera. In this preliminary we will consider the folllowing :
General appearance of the larvae of Bibionomorpha.
Generally, a larva of a fly of the Bibionomorpha has a head, which is more or less sclerotized, three thoracic segments, and about nine abdominal segments. The segments do, or do not, carry projections, spinelets or hairs. These larvae, like the larvae of all diptera, do not possess genuine legs. In most cases the appearance is more or less worm-like. See Figure 2 above .
Tracheal system of the larvae of diptera.
Often the sides of the segments carry spiracles, which are the exits of the tracheal system, guaranteeing contact with the air. Depending on which segments carry spiracles (one on the right side and one on the left side) we can distinguish six types of tracheal (pneustic) systems in dipterous larvae :
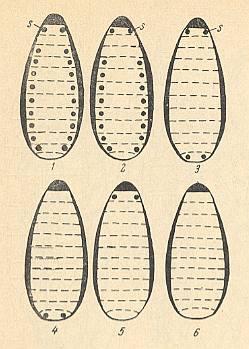
Figure 6 : Types of the tracheal system in dipterous larvae.
1 -- Holopneustic.
2 -- Peripneustic.
3 -- Amphipneustic.
4 -- Metapneustic.
5 -- Propneustic.
6 -- Apneustic.
s = spiracle.
(After KRIVOSJEINA, 1969)
Let us describe -- following KRIVOSJEINA, 1969, pp. 141 -- these types.
Head and mouthparts of larvae of (nematocerous) Diptera
Prior to presenting the data (given by KRIVOSJEINA) concerning the families Pleciidae, Hesperinidae, Pachyneuridae, and Axymyiidae, we will briefly expound the general structure of the head and its appendages (mouthparts) of the (nematocerous) dipterous larvae.
But before we do this we say something about the g e n e r a l structure of the mouthparts of insects (whether adults or larvae) as such. See next Figure, which depicts the mouthparts of a very generalized ( = primitive, original) but still recent (primarily wingless [ = apterygote] ) insect. Here the main parts, such as the maxillae (= lower jaws) and the labium (= lower lip), are still complete, that is, no parts of them have been evolutionarily disappeared or changed beyond recognition.
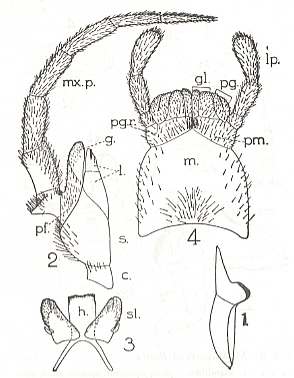
Figure 7 : Mouthparts of Petrobius maritimus (Apterygota, Machilidae).
1 -- Mandible. 2 -- Maxilla. 3 -- Hypopharynx (h) and superlinguae (sl). 4 -- Labium (= lower lip).
The Labrum (= upper lip, not drawn) only functionally belongs to the mouthparts. Morphologically it is associated with the head-shield.
Maxilla : c = cardo, s = stipes, l = lacinia, g = galea, pf = palpifer,
mx.p = maxillary palp.
Labium : m = postmentum, pm = prementum, pgr = palpiger, gl = glossa, pg = paraglossa, lp = labial palp.
(After RICHARDS & DAVIES, Imms' General Textbook of Entomology, 1977)
Essentially, the mouth-apparatus comprises three pairs of appendicular jaws :
The larvae of most representatives of the suborder Nematocera possess a developed (and sclerotized) head-capsule. This head-capsule is like a short tube enclosing the internal parts of the head, first of all its internal skeleton -- the tentorium, but, of course, also the suboesophageal ganglion, the oesophagus (gullet), and the other internal organs. Often the head-capsule is dorsally extended forwardly, more or less forming a roof under which lie the mouthparts-proper -- mandibles, maxillae, and labium. The labrum, which functionally belongs to the mouthparts, is the distal part of the mentioned roof. The dorsal part of the head-capsule consists of a median frontal plate + clypeus (= anterior head-shield), often fused, resulting in one single plate, the fronto-clypeal plate), and distally the labrum is attached to the clypeus. Laterally attached (often by a seam) to the fronto-clypeal plate are the side-plates (pleural plates) of the head-capsule (one plate on the right, the other on the right). They bend down around the sides of the head, and then approach each other again at the ventral side of the head. There they fuse together along a shorter or longer line or remain separated (only connected by a non-sclerotized patch). The eyes and antennae are placed more or less fronto-laterally on the head-capsule. Also the maxillae possess antenna-like sense organs : maxillary palps (one on each maxilla).
Let us now succinctly describe the basic (and often modified or reduced) structure of the head of dipterous larvae as this (general) structure is expounded and analyzed by KRIVOSJEINA, 1969, pp. 55, and pp. 58 :
Up until today [1969] there does still not exist one definite understanding about the number of metamers (body-segments) and the morphological interpretation of the appendages of the insect head. Generally acknowledged is the fact that the head of the arthropods (insects, spiders, lobsters, etc.) is formed by the acron (head-lobe [-blade, -sheet], homologous with the prostomium of the annelids [ring-worms] and some segments coming [morphologically] after it. However, the question about the number of segments which, together with the acron take part in the formation of the head of the arthropods has been differently answered. The majority of investigators is inclined to believe that in the formation of the head of insects five segments are involved : the antennal, intercalary, mandibular, maxillar, and labial segment, where the intercalary segment is only present in embryos.
The head-capsule proper (epicranium) is just the total of outer sclerotized elements of the head, excluding the eyes and the mouth-appendages.
Within the order Diptera we can observe in the larvae all transitions from a completely developed head-capsule to a totally reduced one, where even whatever trace of the capsule and mouthparts can be absent.
Depending on the stage of reduction one can usually distinguish three types of head-capsule : (1) a well-developed massive head-capsule (Eucephala), (2) a partly reduced capsule (Hemicephala), (3) a totally reduced head-capsule (Acephala).
This division in its most general form expresses the chief direction of the evolution of the [larval] head in the order Diptera. But, while the first and last category are sufficiently definite, groups with partial reduction of the head-capsule include very heterogeneous forms.
The change and refinement of the ability to feed in dipterous larvae have in the evolutionary process led to the origin -- within the Order -- of the most diverse types of head-capsule and mouthparts [oral apparatus]. The overall tendency detected within the Order Diptera is a general simplification, and in a whole series of cases the reduction, of individual elements of the head-capsule and of the mouth-apparatus or of the head as a whole. There takes place a replacement of many elements of the mouth-apparatus (well-developed in the majority of insects and lower diptera) by just one pair of mouthhooks in the cyclorraphic diptera. Such changes lead to a fundamental transformation of the musculature of the head and of the very head-capsule.
One can, as does KRIVOSJEINA, pp. 58, distinguish within each of the three above mentioned groups ( Eucephala, Hemicephala, Acephala ) a number of structural types of the head-capsule and mouth-apparatus in the larvae of diptera. For our preliminary understanding of these larval strucures in the families Pleciidae, Hesperinidae, Pachyneuridae, and Axymyiidae it is sufficient to describe -- as a whole -- the first group, the Eucephala, (consisting of several types), as it is defined by KRIVOSJEINA :
Group I -- Larvae with a well-developed and [thus] not reduced head-capsule (Eucephala).
In the capsule the central part is well-distinguished (frontal plate) and also the lateral plates of the head, which ventrally fuse with each other, that is, touch each other in one or several points or, on the other hand, are more or less separated. The internal skeleton of the head (tentorial rods), if present, lies within the confines of the head-capsule. Mouth-apparatus well developed, of the biting type, all its parts expressed, and the mandibles and maxillae completely [morphologically] independent of each other.
To this group belong the larvae of the majority of nematocerous diptera [midges, mosquitoes, crane-flies, etc.].
Depending on the nature of the structure of the head-capsule and the mouth-apparatus within the range of this group we can distinguish seven basic types of head-capsule : the Trichocerid-Anisopodid, the Chironomid, the Ptychopterid, the Culicid, the Ditomyiid, the Mycetophilid, and the Bibionid type.
Without describing all these types one by one (which is done by KRIVOSJEINA), we will describe one type only, namely the Bibionid type, which is especially relevant for interpreting the genera Plecia, Penthetria, Pachyneura, Hesperinus, Axymyia (and also Protaxymyia, Mesaxymyia). In it we will describe and depict the structure of the head-capsule and the mouthparts of the larvae of a species of the genus Plecia and a species of the genus Bibio ( Bibionidae).
Bibionid type (KRIVOSJEINA (1969), pp. 70).
Head-capsule with massive side-plates which are shaply narrowed on the ventral side. The elements of the mouth-apparatus are heavily sclerotized. The head-capsule expressed by this type is characteristic of the larvae of Bibionidae, Pleciidae, Pachyneuridae, and Hesperinidae. Let us, as an example, analyze the morphology of the head of the little known larva of Plecia nigra LUND. See next Figures.
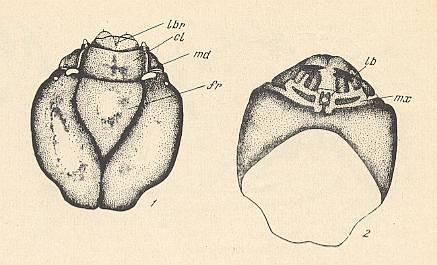
Figure 8 : Head of the larva of Plecia nigra LUND. ( Pleciidae).
1 -- head, dorsal view. 2 -- head, ventral view.
lbr = labrum (upper lip), cl = clypeus, md = mandible, fr = frons, lb = labium (=lower lip: the small light-colored more or less circular [as drawn] part at the center of the ventral side of the head-capsule), mx = maxilla.
(After KRIVOSJEINA, 1969)
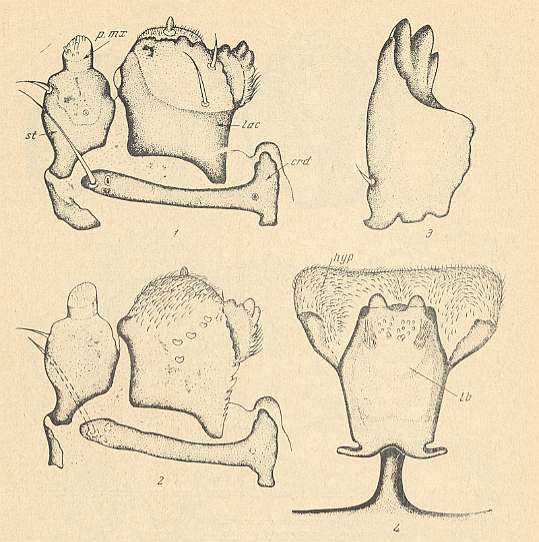
Figure 8a : Mouth-apparatus of the larva of Plecia nigra LUND. ( Pleciidae).
1 -- Maxilla, ventral view. 2 -- Maxilla, dorsal view. 3 -- Mandible. 4 -- Labium and Hypopharynx, ventral view.
crd = cardo, hyp = hypopharynx, lac = lacinia, lb = labium, p.mx = maxillary palp, st = stipes.
(After KRIVOSJEINA, 1969)
In the central part of the head a small triangular frontal plate is clearly delimited. The side-plates of the head-capsule are large, massive, fused at the ventral side. The seam at the place of their unification is black. The clypeus is represented by a massive rectangular plate making an angle (in a lateral view) with the frontal plate and is clearly distinguished from it. The antennae are strongly reduced. They present themselves as elongate-oval lightly-colored humps lying at the base of the clypeus. The eyes are positioned behind them at the sides of the head. They are reprepresented there by small transparent vesicles.
Most similar to the head-capsule of Plecia nigra LUND. is the head-capsule of the larvae of the family Bibionidae. See next Figure, where in its subscript one can click to some magnified parts of this Figure :
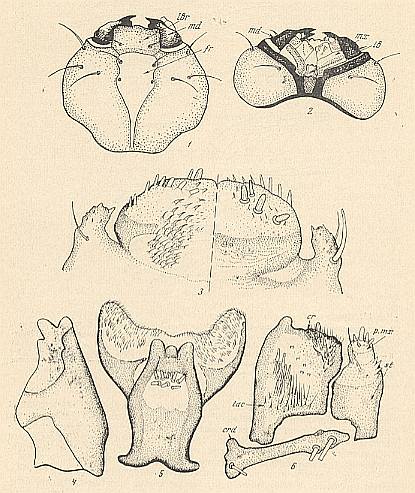
Figure 9 : Details of the structure of the head of the larva of Bibio pomonae FABR. ( Bibionidae).
1 -- head, dorsal view (see HERE, upper figure ).
2 -- head, ventral view (see HERE, lower figure ).
3 -- upper lip (labrum), right : dorsal view, left : ventral view.
4 -- mandible.
5 -- lower lip (labium) and hypopharynx.
6 -- maxilla (see HERE ).
cr = comb. crd = cardo. fr = forehead (frons). lac = lacinia. lb = lower lip. lbr = upper lip. md = mandible. mx = maxilla. p.mx = palp of maxilla. st = stipes.
(After KRIVOSJEINA, 1969)
The head-capsule of the larvae of Pachyneura ZETT. and Hesperinus WALK. shows a series of essential differences, but still preserving the same structural type.
In the larvae of Pachyneura ZETT. (see HERE ) and Hesperinus WALK. (see HERE ) the side-plates of the head-capsule are not fused ventrally, The lower lip (labium) is morphologically distinguished from the elements of the head-capsule. The frontal plate and the clypeus are almost fused. They are separated by a weakly expressed seam and clearly distinguished from the upper lip (labrum). The elements of the mouth-apparatus of Hesperinus WALK. are strongly sclerotized just as in the families Bibionidae and Pleciidae. In Pachyneura ZETT. the fore-margin of the lacinia is membraneous. Only a small part of the cardo is sclerotized.
Characteristic of the head-capsule of the larvae of the present type are the strongly sclerotized elements of the mouth-apparatus, the peculiar rod-like cardo, and the rectangular lower lip.
All this indicates the significant specialization of the head as a whole, which has reached its strongest expression in the larvae of Bibionidae.
Of the [mentioned] outgrowth, unifying the head-capsule of the larvae of Bibionidae and Pleciidae with the lower lip, do not exist homologous structures in other groups.
Of the larval head belonging to either the ditomiid, mycetophilid, or bibionid type, is characteristic the narrowing of the side-plates of the head-capsule on the ventral side, often touching each other in a single point only, which is expressed by the position of the head with respect to the body. In the larvae of Hesperinidae, Pachyneuridae, and Bibionidae, the head clearly is hypognathous, which is also characteristic of mycetophilid diptera, although in some cases a secondary expansion of the ventral walls of the head-capsule is observed, or supplementory massive sclerites develop which join up with the structure of the head-capsule.
Having expounded the general appearance, the tracheal system, and the general morphology of the head and mouthparts, of nematocerous larvae, we are now, finally, ready to give the exposition of KRIVOSJEINA (1969) pp. 229 [to which exposition we add things when necessary], dealing with the mentioned nematocerous families : Pleciidae, Hesperinidae, Pachyneuridae, and Axymyiidae, that is, interpreting the genera Plecia, Penthetria, Pachyneura, Hesperinus, Axymyia (and also Protaxymyia, Mesaxymyia), all belonging to the infraorder Bibionomorpha in the neighborhood of the superfamilies Fungivoridea and Bibionidea ( The Pachyneuridae might well belong to the Tipulomorpha, especially by reason of the imaginal features [that is, the features of the adult fly] ).
Now the exposition of KRIVOSJEINA (with additions and small changes where necessary) :
The mentioned families are relict groups of diptera, known in the world-fauna by 1-3 genera :
Plecia WIED., Pleciidae.
Penthetria MEIG., Pleciidae.
Pachyneura ZETT., Pachineuridae.
Hesperinus WALK., Hesperinidae.
Axymyia Mc ATEE., Axymyiidae.
Protaxymyia MAM. et KRIV., Axymyiidae.
Mesaxymyia MAM., Axymyiidae.
Up until today [1969] the systematic position of the listed genera had remained very obscure.
The opinions of the greatest dipterologists of the world fundamentally differ with respect to the taxonomic rank of these groups as well as to their position in the taxonomic system. In the monograph of Duda (Duda, 1930) the representatives of the genera Plecia WIED., Penthetria MEIG., Pachyneura ZETT., Hesperinus WALK., together with Axymyia Mc ATEE., are considered within the confines of the family Bibionidae. Hennig, 1954, while investigating the phylogeny [in the sense of setting up a purely phylogenetic system exclusively based on genetic kinship relations] of the Diptera, considers Hesperinus WALK. together with Penthetria MEIG. and Bibio GEOFFR., but points to their [taxonomic] independency. But Pachyneura ZETT. and Axymyia Mc ATEE. are included by him in the mycetophiloid complex [roughly coinciding with the superfamily Fungivoridea].
Accordingly, some investigators place the representatives of the mentioned genera closely to, or into, the family Bibionidae, while others place them closely to the mycetophiloid complex. The question concerning the taxonomic rank of these groups also remained in dispute. The difference of opinion becomes clear by the fact that in the majority of cases investigators attributed great phylogenetic significance to [mere] secondary features. But the correctness of this or that viewpoint cannot be established because of the absence of expositions concerning the ontogenesis [here it is meant : the morphology of the larvae] of the representatives of these groups.
Our [Krivosjeina and Mamajev] [over many years] continued collecting of larvae have partly met with success. In the Coastal district [Primorski krai] in the years 1964 and 1967 a large amount of larvae of the representatives of these families have been found and collected.
[The larva as well as the imago will be investigated in the present text. The next Figure, taken from KRIVOSJEINA, depicts the wing-venation of the insects under investigation together with that of some other more or less closely related forms.]
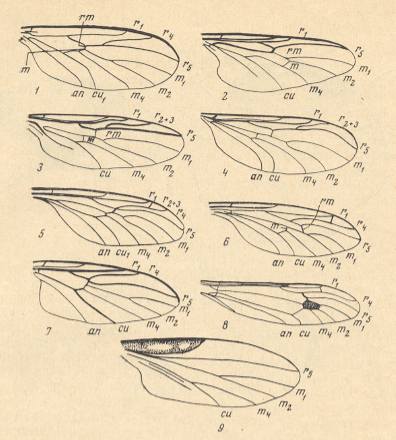
Figure 10 : Wings of the representatives of the families Ditomyiidae, Bibionidae, Pleciidae, Hesperinidae, Pachyneuridae, Axymyiidae, Mycetobiidae, Tipulidae, and Leptoconopidae. In order to ease the reading of the subscript the reader can click on "Figure".
1 -- Ditomyia fasciata MEIG. ( Ditomyiidae ). ( Figure )
2 -- Bibio marci L. ( Bibionidae ). ( Figure )
3 -- Plecia nigra LUND. ( Pleciidae ). ( Figure )
4 -- Hesperinus rohdendorfi KRIV. et MAM. ( Hesperinidae ). ( Figure )
5 -- Pachyneura sp. ( Pachyneuridae ). ( Figure )
6 -- Mesaxymyia kerteszi DUDA. ( Axymyiidae ). ( Figure )
7 -- Mycetobia pallipes MEIG. ( Mycetobiidae ). ( Figure )
8 -- Idiotipula confluens ALEX. ( Tipulidae ). ( Figure )
9 -- Leptoconops borealis GUTZ. ( Leptoconopidae ). ( Figure ).
an = anal vein, cu = cubital vein, m1-4 = medial veins, m and rm = radio-medial cross-vein [we might also write : r-m ), r1-5 = radial veins [ r1 = Radius, r2 + r3 + r4 + ... = Radial Sector ].
(From KRIVOSJEINA, 1969, 1-7, 9 after KRIVOSJEINA, 1969, 8 after HENNIG, 1954)
On the basis of this material, let us analyze the mutual connection of these very ancient groups with the families of the bibionid and mycetophilid complexes of diptera, first of all with the Ditomyiidae as representing the least specialized among the mycetophilids, and also with family Bibionidae, in which the representatives of the mentioned genera were placed by many investigators.
Ditomyiidae, larva.
The larvae of the representatives of the family Ditomyiidae possess a well-developed head-capsule, with side-plates which ventrally meet each other along a short line, and with a clypeus clearly distinct from the frons (frontal plate). See HERE for Symmerus annulatus.
The upper lip (labrum) is broad, transverse. The mandibles are massive and rectangular. The maxillae and lower lip (labium) blade-like, weakly sclerotized.
Body with clearly expressed 12 segments without secondary segmentation, naked, only with rare small spinelets on the dorsal as well as on the ventral side.
Tracheal system peripneustic (see Figure 6 (2), above ), the spiracles are placed on the prothorax [ = anterior part -- first segment -- of thorax] and on 8 abdominal segments, each with a large central moult seam and a grated peritrema [We will not expound these latter features as to what they precisely are.].
Middle gut with two blind sacs at its anterior section. The Malpighian tubules are simple, isolated. [ To give the reader an impression of the digestive system of larvae of Nematocera, we have a drawing of this system as it is in Mycetophila, which genus does not, it is true, belong to the Ditomyiidae, but to the closely related family Mycetophilidae ( = Fungivoridae). And both families belong to the superfamily Fungivoridea. The drawing, then, is HERE ].
Ditomyiidae, imago.
The imago of the representatives of the Ditomyiidae has usual, a little elongated legs.
The wing-venation is weakly costalized, and a clear radial sector is characteristic. The long r1 (Radius) ends (in the Costa) at the level of the distal half of the wing. See next Figure (In our text, in the signs for the veins, we do not use subscripts, we place the numbers just next to the letters).
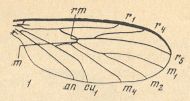
Figure 10_1 : Wing of Ditomyia fasciata MEIG.. (1 of Figure 10).
The Radial Sector branches (into two branches) distally from the cross-vein r-m , and R5 ends up at the wing-tip. The cross-veins r-m and m lie in the central part of the wing and make an angle with each other, where r-m lies (a bit) more distally than m. The medio-cubital cross-vein (mcu), like the medial cross-vein (m), makes an angle with r-m [In the interpretation of KRIVOSJEINA the short vein that connects M4 with Cu1 is the medio-cubital cross-vein (mcu), while in the interpretation of other authors it might just be the proximal part of the anterior branch of the cubital fork]. Both veins (m and mcu) look little like cross-veins and make only a small angle with the costal vein of the wing. The central trunk of the medial system of veins is absent. The discoidal cell is absent [ For the discoidal cell, see Figure 10 (8), where this cell is present and marked in the drawing.] The anal vein (An1) goes all the way to the posterior wing margin.
Bibionidae, Larva.
Bibionidae, imago.
In the adult form of the representatives of the family Bibionidae the first antennal segments are clearly distinguished and short. Fore-tibiae are thickened and shortened, with combs of spines or massive spurs.
The wing-venation is strongly costalized. Only two long radial veins exist [Radius and an unbranched Radial Sector]. The first ends in the fore-margin past its middle, the second ends up at the costal vein, it does not continue down to the wing-tip. See next Figure.
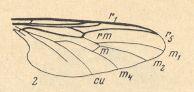
Figure 10_2 : Wing of Bibio marci L. (2 of Figure 10). [ For comparison with the wing of Ditomyia, see Figure 10 (1)(2) ]
The cross-veins r-m and [especially] m are shifted towards the wing-tip, they are almost parallel to each other and make an angle with the costal vein of almost 90 degrees. The trunk of the medial system of veins is well developed, it forks at the level of the medial cross-vein (m). Discoidal cell is absent. Anal veins clearly do not reach the wing-margin.
Let us now compare to the Ditomyiidae (Fungivoridea) and Bibionidae (Bibionidea) representatives of the genera Plecia WIED., Hesperinus WALK, Pachyneura ZETT., Axymyia Mc ATEE., Protaxymyia MAM. et KRIV., Mesaxymyia MAM., that is to say, to the representatives of the families Pleciidae, Hesperinidae, Pachyneuridae, and Axymyiidae.
Family P L E C I I D A E
The majority of dipterologists included and include Plecia WIED. and Penthetria MEIG. in the family Bibionidae, in which usually these genera were distinguished as representing a subfamily. However, detailed investigation of the larvae of Plecia WIED. and articles in the literature concerning the larvae of Penthetria MEIG., bear witness to the significant idiosyncrasy of the larvae of both genera, making it legitimate for us to consider the subfamily Pleciinae as being in fact a separate family, Pleciidae. This conclusion, which was based on a study of the larvae, is also confirmed by data concerning the imago.
In the larvae of Plecia WIED., see Figure 8 and Figure 8a , we can observe differences with the Bibionidae in the structure of the fronto-clypeal region, of the spiracles, and of the topography of projecions [outgrowths] on the body. The clypeus is clearly distinct from the frons [frontal plate] and makes an angle with it, while in Bibionidae we have to do with a single undivided [fused] structure. The lacinia is very peculiar, [in] having an extra side-plate (see Figure 8a-2 ). The last [pair of] spiracles in this genus (Plecia), as in many diptera, are simple, represented by a plate carrying a moult seam in the center and grated holes at the periphery.
The middle gut with two lateral anterior blind sacs and with one posterior sac. Malphigian tubules close to each other.
Accordingly, in the structure of the head, of the mouth-apparatus, the spiracles, and of the digestive system, of the larva, we can observe clear differences from the larvae of Bibio GEOFFR. and Dilophus MEIG., which we consider to be representatives of the family Bibionidae. But the larvae of Plecia WIED. possess a whole series of features which connect them with the Bibionidae. The similarity expresses itself in the global shape of the body, the development of soft conical projections on the segments (see Figure 2 ), the degree of sclerotization of parts of the mouth-apparatus, and the position of the Malpighian tubules. But also these features have their specifity. Thus in the larvae of Plecia WIED. along the posterior margin of the last segment, laterally and ventrally are placed eight soft conical outgrowths which are absent in the larvae of Bibionidae (see Figure 2 ).
The structure of the wing, as well as of the legs, of the imago, sharply distinguishes the representatives of Plecia WIED. from those of the Bibionidae.
In the imago the antennae have short but clearly visible first segments. The legs are simple, without series of combs. Fore-tibiae normal, elongate, not thickened.
The wing-venation, see next Figure, is weakly costalized.
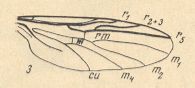
Figure 10_3 : Wing of Plecia nigra LUND. (3 of Figure 10). [ Next to the cross-vein (m) between M1+2 and M4 there is drawn the sign "m" ]. [ For comparison with the wings of Ditomyia, and Bibio, see Figure 10 (2)(3) ]
There are three, not two, radial veins [namely R1, R2+3, R5 ]. The first runs into the wing-margin past its mid-point. The third ends up before the wing-tip. The Radial Sector forks into two branches distally from the cross-vein r-m (radio-medial cross-vein). The cross-veins (r-m and m) lie in the middle of the wing, and r-m is parallel to m, making a right angle with the Costa. The common trunk of the medial system of veins is well developed. It forks distally from the medial cross-vein (m). The discoidal cell is absent. The anal veins do definitely not reach the wing-margin.
Family H E S P E R I N I D A E
This family is represented by only the one genus Hesperinus WALK., which has 6 species.
For a long time the dipterologists did not agree concerning answering the question about the taxonomic rank of the genus as well as the question concerning its position in the taxonomic system of the Order. Hendel (1928, 1936) sees it as constituting the independent family Hesperinidae. Duda (1930) includes the genus Hesperinus WALK. in the subfamily Pachyneurinae of the family Bibionidae. Edwards (1930) also considers the genus Hesperinus WALK. as belonging to the family Bibionidae. Hardy (1945, 1958) repeats this same inaccurateness. Finally, Hennig (1948) and Rohdendorf (1964) placed Hesperinus WALK. in an independent family next to the Bibionidae.
How can we clarify this when considering features of the larvae as well as of the imago?
The larvae [of Hesperinus] superficially partly recall the larvae of the bibionids, and partly those of the mycetophilids.
The head-capsule is strongly sclerotized, almost black (see HERE, for a drawing ). The side-plates of the head-capsule not touching each other ventrally [meant is apparently that they are not ventrally fused with each other]. The clypeus is clearly distinct from the frontal plate. The structure of the mouth-apparatus (see HERE, for a drawing ) has much in common with that of the larvae of Bibionidae : Upper and lower lip of the same structural type, so also the mandibles and the maxillae. But the elements of the lower lip and of the hypopharynx are somewhat more independent. The body consists of 12 clearly distinct segments with a secondarily segmented prothorax. The segments lack projections [outgrowths] of any sort. Only series of spinelets are visible on the body. See next Figure.
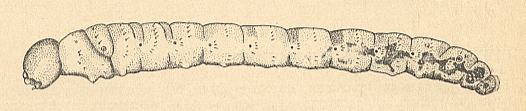
Figure 11 : Larva of Hesperinus rohdendorfi KRIV. et MAM. ( Hesperinidae), lateral view.
(After KRIVOSJEINA, 1969)
Tracheal system holopneustic (see Figure 6 (1), above ). The spiracles lie on the first and last thoracic segments and on the first 8 abdominal segments (meaning that there is no inversion of the spiracles -- that is, no shift of the last pair from the eighth to the ninth abdominal segment). Such a spiracle is represented by an oval plate with a large central moult seam and a grated peritrema. The last body segment lacks spiracles. Middle gut with only two anterior blind sacs. Malpighian tubules close to each other.
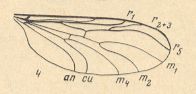
Figure 12 : Wing of Hesperinus rohdendorfi KRIV. et MAM. ( Hesperinidae). (4 of Figure 10).
The wing-venation is moderately costalized. All in all there are three radial veins : Of them R1 runs into the wing-margin past its center. R5 is long, it reaches the wing-tip. Radial Sector forks distally from r-m. This cross-vein r-m runs parallel to the cross-vein m, and both almost make a right angle with the costal vein. The common trunk of the medial veins is well developed. It forks distally from the medial cross-vein (m). Discoidal cell absent. Anal vein reaches the wing-margin.
Family PACHYNEURIDAE
The family Pachyneuridae is represented by the one genus Pachyneura ZETT., which has five species, from which in Europe P. fasciata ZETT. is known. Let us give some short information concerning the history of its interpretation. Hendel (1928, 1936) acknowledges the independent family Pachyneuridae with two genera Pachyneura ZETT. and Axymyia Mc ATEE. Duda (1930) considers it, with that same content, within the family Bibionidae as a subfamily. Edwards (1928) again acknowledges the Pachyneuridae as an independent family, including also the genus Axymyia Mc ATEE, and has them closely related to the Anisopodidae [ = Rhyphidae]. Hennig (1954) also considers Pachyneura and Axymyia, but places them into the mycetophilid complex. Rohdendorf (1964) acknowledges the independent family (without Axymyia ), but places it on the basis of the structure of the wings into the Tipulomorpha. Finally, in the compilation of North American Diptera of the American authors (Stone et al., 1965), the Pachyneuridae are taken in the earlier content (with Axymyia ) and are considered to be closely related to the Anisopodidae. Accordingly, a part of the authors considers the genus Pachyneura to be closely related to the Bibionidae, while the majority places it more closely to the Anisopodidae.
The larvae of Pachyneura are so peculiar that they superficially do not seem to be dipterous larvae at all. See next Figure.

Figure 13 : Larvae of Pachyneura sp. ( Pachyneuridae ).
(After KRIVOSJEINA, 1969)
They look more like caterpillars with their large quadratic head and the long spinelets on the body. But this is just a mere superficial impression. In the structure of the body, its coverings, and the head-capsule, the larvae are certainly close to those of the Bibionidae as well as to those of the Hesperinidae. For the morphology of the larval head, see HERE . The head-capsule is short and broad, the side-plates of the head do not ventrally touch each other. The frontal plate almost fused with the clypeus. It is distinct from it by a weakly expressed seam. The mouth-apparatus is structurally very close to that of Bibionidae [ see HERE ][use BACK button to return] and of Hesperinidae HERE ][use BACK button to return] (blade-like upper lip, massive mandibles, sclerotized lower lip). The cardo of the maxilla is just a very small sclerite, but rod-like in shape. The anterior margin of the lacinia is not sclerotized, it is membranous. On the ventral side of the head, under the side-plates, there are massive sclerotized rods, which can be considered to represent the tentorium.
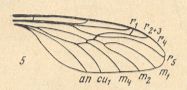
Figure 14 : Wing of Pachyneura sp. ( Pachyneuridae ). (5 of Figure 10).
[In interpreting the wing-venation of Pachyneura we must realize that the common median trunk here is in fact only a weakly expressed vein (according to HENNIG, 1954)]
All in all there are four radial veins. R1 is long, it reaches the distal fourth of the wing. The Radial Sector forks at the level of the cross-vein r-m. R2+3 branches off from R4 almost at the wing-tip. R4 and R5 close to each other at the base, not parallel. R5 long, it reaches the wing-tip. The cross-veins m and mcu make an angle with r-m, and a small angle with the fore-margin of the wing. The cross-veins r-m and m make an angle with each other. The common trunk of the medial veins is developed, it forks distally from the medial cross-vein (m). Discoidal cell absent. Anal vein reaches the wing-margin.
Figure 15 : Wing-venation.
Left -- Ditomyia fasciata MEIG. ( Ditomyiidae ).
Right -- Pachyneura sp. ( Pachyneuridae ).
(1 and 5 of Figure 10).
Similarity is not observed with respect to the number of veins, but to the general scheme of their mutual position, first of all to the relative position of the cross-veins. In Pachyneura ZETT. as well as in Ditomyia WINN. the cross-vein r-m makes an angle with the medial (m) and the medio-cubital (mcu) cross-vein, while the latter makes a very small angle with the wing margin, R5 runs into the Costa at the wing-tip.
Family A X Y M Y I I D A E
Even more disagreement existed about the position of the genus Axymyia McATEE in the system of Diptera. Soon after its description Shannon (1921) placed the new genus in the family Anisopodidae [ = Rhyphidae] as the subfamily Axymyiinae. After Duda (1930), Enderlein (1936) included Pachyneura ZETT. and Axymyia McATEE in the family Bibionidae. Hendel (1928, 1936) brings the two genera closely together constituting an independent family. The same position is held by Edwards (1925), Hennig (1948, 1954) and, finally, by the collective of American authors (Stone et al, 1965). A different position is held by Rohdendorf (1946). He lets Axymyia McATEE represent an independent family which is related to the extinct Paraxymyiidae and Eopleciidae. Newly available material concerning the present group of diptera pointed to the fact that there are at least three genera, namely Axymyia McATEE, Protaxymyia MAM. et KRIV., and Mesaxymyia MAM., which are very closely related to each other in imaginal as well as in larval characters. Their structure is in such a degree idiosyncratic that there will not be any opposition against their forming one single family Axymyiidae. In the study of the larvae, which allegedly had to affirm the position held by the majority of investigators who considered the genera Axymyia McATEE and Pachyneura ZETT. to be closely related, totally unexpected biological features were clarified. [The close relationship between the Axymyiidae and Pachyneuridae is at least suggested by the wing-venation :
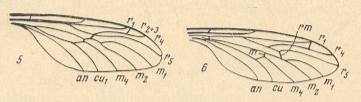
Figure 16 :
5 -- Pachyneura sp. ( Pachyneuridae ).
6 -- Mesaxymyia kerteszi DUDA ( Axymyiidae ).
(After KRIVOSJEINA, 1969 (5 and 6 of Figure 10))
Obtained from yet another source, we again give here the wing-venation of representatives of the Pachyneuridae and the Axymyiidae] :
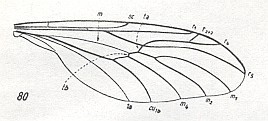
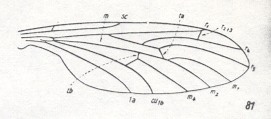
Figure 16a :
80 -- Pachyneura fasciata ZETT. ( Pachyneuridae ). (After DUDA, 1930).
81 -- Axymyia furcata McATEE. ( Axymyiidae ). (After CURRAN, 1934).
(From HENNIG, 1954)
The larvae, judging from their structure, do not have anything in common with any of the other groups under discussion. See next Figure.
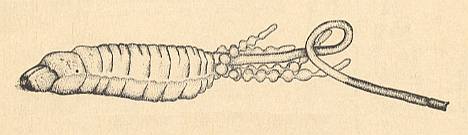
Figure 17 : Larva of Mesaxymyia kerteszi DUDA ( Axymyiidae ). Lateral view.
(After KRIVOSJEINA, 1969)
See HERE for the structure of the head and mouth-apparatus of the larva of Protaxymyia melanoptera MAM. et KRIV. (Axymyiidae).
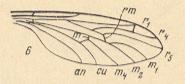
Figure 18 : Wing of Mesaxymyia kerteszi DUDA ( Axymyiidae ).
(6 of Figure 10).
All in all there are four radial veins. R1 long, it reaches the distal fourth of the wing. Radial Sector forks proximally from the radio-medial cross-vein r-m. R2+3 branches off from R4 almost at the apex of the wing. It ends up at the costa very close to [or at] R1. R5 independent, it reaches the wing-tip. R4 and R5 run parallel to each other. The common trunk of the medial system is present, it forks distally from the medial cross-vein (m). Discoidal cell absent. The anal vein reaches the wing-margin [The russian text says it does not, but that is probably a printing error, judging from the above Figures.].
Remark :
[ This latter statement of KRIVOSJEINA's, p. 238, is not very convincing.
As far as I can see, the overall or general plan of the wing-venation in Pachyneuridae and Axymyiidae is identical, and this plan in turn is very similar to that in Mycetobia as well as to that in the mycetophilid complex (for instance in Ditomyia WINN.). The wing-venation of Mycetobia only differs from both Axymyiidae and Pachyneuridae in having a Radial Sector with only two branches, and in clearly having lost the common trunk of the medial veins -- but Pachyneura is in the process of loosing it. Further -- as one more difference -- the wings of Mycetobia possess a broad anal lobe (in contrast to those of Pachyneura and Axymyia ). And, apart from the reduction of the Subcosta, the venational plan in Ditomyia (of the family Ditomyiidae, belonging to the mycetophilid complex (Fungivoridea) as one of its least specialized forms) is identical to that in Mycetobia ].
[ For the wing-venation of Ditomyia WINN., see Figure 10-1 above ].
The general plan of the wing-venation as we see it in Pachyneura, Axymyia, Mycetobia, and in Ditomyia, can be described as follows :
The Axymyiidae (see Figure 18, above ) have four radial veins, and R2+3 ends up in the Costa at the endpoint of R1 or directly into that vein. R4 and R5 run parallel to each other, they do not come closer to each other, r-m is placed significantly distally from the fork of R4+5. Finally, r-m and m lie significantly far away from each other, and r-m lies in the distal half of the wing.
All listed features are characteristic neither of Bibionidae, Hesperinidae, and Pachyneuridae, nor of the mycetophilid diptera. On the contrary, elements of similarity in these respects with a whole series of groups of tipuloid diptera are observed.
Of many groups of Tipulomorpha, characteristic or even typical is the approach of R2 to R1, which is observed for example in Antocha alpigena MIK (Limoniidae), and in many Tipulidae.
See next three Figures and comments of mine.
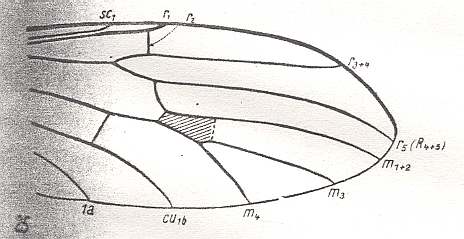
Figure 19 : Venation of apical part of the wing of Antocha alpigena MIK ( Limoniidae ) (the cross-vein tp, drawn as a dashed line, is absent in this species, but is present in other species of this genus. R2 is only very weakly expressed).
(After HENNIG, 1954).
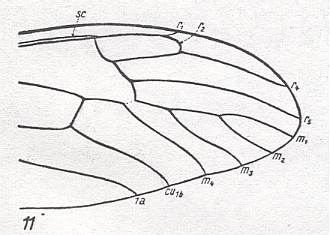
Figure 20 : Venation of apical part of the wing of Dolichopeza albipes STRöM ( Tipulidae ).
(After HENNIG, 1954).
The position of r-m distally from the separation of R4 and R5 (that is, the point where the Radial Sector first forks) is also typical of many groups of the Tipulomorpha ( Tipulidae, Limoniidae, Ptychopteridae), see Figure 19. In Figure 20 (Dolichopeza) we can see this feature also, but here the Radial Sector itself branches off very distally from the Radius (R1).
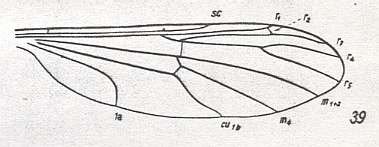
Figure 21 : Venation of Bittacomorphella nipponensis ALEX. ( Ptychopteridae ).
(From HENNIG, 1954, after ALEXANDER, 1927).
In Bittacomorphella r-m lies only a little bit distally from the primary fork of the Radial Sector. And also here -- as well as in Liriope scutellaris MEIG. (also belonging to the Ptychopteridae) ( HENNIG, 1954, Figure 38) -- R2 approaches R1 such that it ends up in it.
The same [that is, similar situation in Axymyiidae and certain Tipulomorpha] can be said about the position in the wing of r-m, namely that it lies in its distal half.
As a result [of enquiries] we [KRIVOSJEINA, p. 238] can find many examples demonstrating the similarity in the structure and distribution of veins in the wing of Axymyia McATEE and certain tipulomorphs. Only the presence of an extra cross-vein [which in HENNIG, 1954, is called tp, and which closes the discoidal cell of the wing-venation] distinguishes [for example] the wing of Idiotipula confluens ALEX. ( Tipulidae) from that of Axymyia. See Figure 10 (8), above . This we can find also in other Tipulomorpha, such as in Pales dorsalis FABR. ( Tipulidae), Xiphura atra L. ( Tipulidae), Phalacrocera replicata L. ( Cylindrotomidae), Limnophila ferruginea MEIG. ( Limoniidae), and Trichocera regelationis L. ( Trichoceridae). See next two Figures.
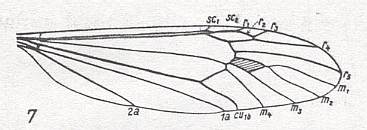
Figure 22 : Venation of Pales dorsalis FABR. ( Tipulidae ). Discoidal cell present (that is, closed by the cross-vein tp ).
(After HENNIG, 1954).
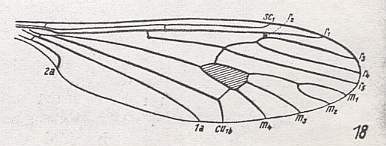
Figure 23 : Venation of Trichocera regelationis L. ( Trichoceridae [ = Petauristidae] ). The Trichoceridae belong to the superfamily Tipulidea of the Tipulomorpha. Discoidal cell present (that is, closed by the cross-vein tp ).
(After HENNIG, 1954).
In Antocha alpigena MIK. ( Limoniidae) -- see Figure 19, above -- there is even no such cross-vein. The discoidal cell [therefore] is absent, although present in other species of this genus.
Analysis of the larval and imaginal features of the Axymyiidae demonstrates that they cannot be closely related to Pachyneura ZETT. or to bibionids. The larval features of Axymyiidae show them to be most closely related to the group Anisopodidae-Mycetobiidae, but this is very conditional. One must admit that an analysis of imaginal features (especially of the wing) and larval features do not provide us with the possibility to judge about to whatever known group of diptera the Axymyiidae are [phylogenetically] related. All this at least indicates the existence of this group as a whole, not only as an independent family, but also as a superfamily.
After having dealt with the genera Plecia, Hesperinus, Pachyneura, and Axymyia, and having shown them to represent independent families, it is perhaps instructive to discuss yet another problematic genus, namely Mycetobia, normally considered to belong to the mycetophilid complex. It represents the family Mycetobiidae.
Family M Y C E T O B I I D A E
The family consists of the single genus Mycetobia MEIG. with a small number (less than 10) of species. Based on imaginal features, the genus was for a long time included into the Mycetophilidae. The study of the early stages of [individual[ development had shown that this was totally wrong. The amphipneustic larvae (see Figure 6 (3), above ), with their elongate mouthparts and 2-segmented mandibles, and, finally, with secondary segmentation of the body, do not have anything in common with the larvae of mycetophilid diptera. See next Figure.
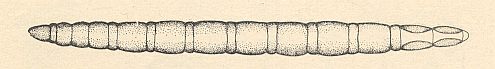
Figure 24 : Larva of Mycetobia sp. ( Mycetobiidae).
Compare the general appearance of this larva with that of a representative of the family Anisopodidae [ = Rhyphidae], namely Sylvicola fenestralis SCOP., HERE . Compare it also with a larva of the family Mycetophilidae [ = Fungivoridae] (belonging to the mycetophilid complex) : HERE .
(After KRIVOSJEINA, 1969)
For details of the head of the larva of Mycetobia, see HERE . To compare this with the larva of Symmerus ( Ditomyiidae, Fungivoridea [ = mycetophilid complex] ), see HERE [press BACK button to return]. [We see here the great difference between the head of the larva of a representative of the genus Mycetobia and that of a representative of the mycetophilid complex, being an indication that Mycetobia taxonomically stands far away from the mycetophilid complex of diptera.]
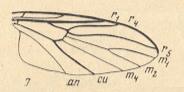
Figure 25 : Wing-venation of Mycetobia pallipes MEIG. ( Mycetobiidae).
( 7 of Figure 10)
The Radial Sector consists of two branches, so all in all there are three radial veins. Forking of the Radial Sector takes place at the level of the cross-vein r-m. R5 runs into the Costa at the wing-tip. The common trunk of the medial system of veins is absent. The cross-veins r-m and m lie almost along a straight line, that is, they do not make an angle, and they make an almost right angle with the anterior wing-margin.
Figure 26 :
7 -- Wing-venation of Mycetobia pallipes MEIG. ( Mycetobiidae).
1 -- Wing-venation of Ditomyia fasciata MEIG. ( Ditomyiidae).
5 -- Wing-venation of Pachyneura sp. ( Pachyneuridae).
( 7, 1, 5 of Figure 10)
The idiosyncratic posistion of the cross-veins indicates that the representatives of Mycetobia stand far away not only from the representatives of Pachyneura, but also from those of Ditomyia. [For all of them, the cross-vein r-m connects the Radial Sector with the Media (M1+2), while the cross-vein m (often called mcu) connects the Media (M1+2) with the Cubital Fork, and this Cubital Fork itself consists of Cu1 and M4 (so the cross-vein m in fact connects M1+2 with M4). But the three genera differ among each other in the position and direction of these two cross-veins.]
This concludes the discussion concerning certain problematic nematocerous genera, as we had found it in KRIVOSJEINA, 1969, who was determining the taxonomic position of them on the basis of larval morphology and wing-venation.
In the next document we will continue with the superfamily Bibionidea of the infraorder Bibionomorpha, namely determining to what functional wing-types the wings of their representatives belong.
As has been explained earlier, the wing-type is the morphological expression of the flight-regime of the possessors of such wings, that is, a particular functional wing-type expresses a particular way of flying of the insect. And this particular way of flying -- flight-regime -- is compatible with, that is, neatly fits into (in the sense that it may not interfere with) the precise way the insect experiences its environment by means of its antennae, but especially by means of its eyes (which are well developed in almost all diptera). In this way, the functional structure of the wings expresses the environment of the fly (as it is experienced by it).
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part IX.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V
{kind=link}