
(Sequel to)
The theory on Substance and Evolution to be presented here is a further development of ideas developed in the previous documents, that is, in the documents devoted to the theory of the true and ultimate determinants of the material world, namely the If-Then constants (these constants themselves residing in the Implicate Order), together constituting causal chains, causal webs, and the determinative connections in low-level and high-level material processes, especially when they are involved in the formation of stable, enduring, repeatable patterns in the material world (space-time world, Explicate Order) : Chreodes, Organisms, and Evolution.
e-mail :
 ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam ) A new interpretation of Aristotelian Substance.
Introduction.
In First Part of Website we discussed the Aristotelian-Thomistic metaphysical theory of Substance in terms of modern dynamical systems theory ( Also in present Part of Website, Fifth Part, we discussed Substance : there one finds the exposition of some Aristotelian principles, among which the pros hen nature of Beings, and further one finds there the recent development of the appropriate Logic specifically geared to this metaphysics of Substance, viz., Intentional Logic ).
In the documents after the development of Intentional Logic (Present Part of Website) we have set up a theory about the determinative essence of the material world (space-time world, including organisms). There we found out that, in addition to this material world, there must be some sort of noëtic, that is, thought-like, domain that accounts for the observed determinative structure of the material world. The nature of this noëtic domain is postulated to be more or less like the "Implicate Order" as proposed by David BOHM in 1980. The features of this Implicate Order relate to the corresponding features in the space-time world -- the Explicate Order -- by processes of injection and projection, that is, projection, or unfoldment up-along-the-space-and-time-dimensions, into the Explicate Order, of "noëtic contents" from the Implicate Order, and, conversely, injection, or enfoldment (back) down-along-the-space-and-time-dimensions, of physical space-time patterns from the Explicate order into the Implicate Order, that is to say that having been injected into the Implicate Order the original space-time structures do not extend anymore along space-time dimensions, they are now enfolded, purely noëtic entities comparable with mathematical entities.
In the Implicate Order do, intrinsically and originally, reside the mentioned If-Then constants, which, by processes of injection and projection constitute the determinative structure of the Explicate Order. But, to account for crystals -- as stable patterns of atoms or molecules, but especially to account for organisms and their intrinsic individual development, we had to concede that apart from the elementary If-Then constants (the 'atomic' noëtic entities), originally present in the Implicate Order, gradually more higher-level noëtic entities, that is, noëtic patterns, came to populate the Implicate Order as a result of what happens in the Explicate Order. Definite higher-level patterns, as have come into being in the Explicate Order, have, through injection and projection, their counterparts in the Implicate Order. In the Explicate Order we observe, that is, encounter, those patterns as structures extended along space and time dimensions, while in the Implicate Order we 'see' those same patterns as totally enfolded, resulting in purely noëtic contents, corresponding to the "formal contents" of those same patterns as they are present in the Explicate Order. Of course, in the Explicate Order there are no formal contents existing as such. Any such formal content there is unfolded along space and time dimensions and therefore exists in the form of one or more physical individuals.
Because the Implicate Order is supposed to be space- and timeless we need a special interpretation of "the Implicate Order successively becoming populated with more and more higher-level noëtic patterns as a result of what happens in the Explicate Order".
The 'time element' of the Implicate Order is totally derived from the time element of the Explicate Order. If we say that 'now', say at point in time A, a given (higher-level) noëtic content, let us call it N, has 'for the first time' appeared in the Implicate Order, the point in time A is seen from the Explicate Order : If we contemplate the overall content of the Implicate Order before the point in time A -- as determined in the Explicate Order -- we do not find N (in the Implicate Order). But if we contemplate the content of the Implicate Order at or after the point in time A, then we will find N (in the Implicate Order) (of course provided such a thing as an Implicate Order, as it has been described, really exists, and if we could actually look into it).
The duality of Implicate and Explicate Orders can account for the repeatable generation of certain stable patterns or structures as we see them in the Explicate Order. So the repeatable generation of individual crystals (crystallization) or of individual organisms (morphogenesis) of a given species had acquired a rationale by assuming the appearance of so-called "chreodes" (as complex higher-level noëtic determinative patterns) in the Implicate Order.
But then the phenomenon of organic evolution, that is, of historical development of organisms during vast expanses of time, demanded still more adjustments of the determinative theory (nomological theory) so far developed. Organic evolution is characterized by the emergence of adaptations of organisms to new environments or to new aspects of existing environments. Many of these adaptations are so complex, and, if we might say, so 'smart', that current biological science cannot reductionistically explain them by its -- we could say, one and only -- method of random genetic mutations (on the level of single organisms or of populations) PLUS natural selection. This method will, in most, if not, all cases, run into a "reductionistic nightmare", that is, it will become bogged down into an impenetrable entanglement of causal factors and trajectories. We have assumed that the cause of this situation is not the alleged insufficiency of scientific methods and techiques available today. The insufficiency of the reductionist scientific method to fully explain the mentioned biological adaptations, as they are observed in the living world, is one in principle. According to us it points to the existence of a domain which is ontologically different from the space-time world : the noëtic Implicate Order. Having already accepted this domain in order to have a foundation for repeatable higher-level patterns as they are seen to be generated in the space-time world, we had to extend its character still further, namely to assume the regular occurrence in it of 'reactions' between noëtic entities, reactions analogous to chemical reactions.
Indeed, in this way we succeeded to account for the generations of complex adaptations as we see them in the evolution of organisms. Such a noëtic reaction was then supposed to take place between (1) the noëtic (formal) content of some given organic species and (2) the noëtic (formal) content of one of the species' potential or actual ecological niches. While the reaction with the formal content of the actual ecological niche of the species could result in a modification of this species' formal content such that an existing adaptation becomes strengened (still noëtically, and only visible in the Explicate Order upon projection into it), the reaction with the formal content of a potential ecological niche of the species could result in a modification of the species' formal content such that an entirely new adaptation had appeared (still noëtically, and only visible in the Explicate Order upon projection into it).
But in addition to progressive evolution, resulting in biological diversification, there is also regressive evolution, resulting in a decrease of diversity : species can become extinct. When all species of a given taxon (such as a genus, family, etc.) become extinct, the taxon itself has become extinct. When, on the other hand, not all, but neverthless most species of a taxon have become extinct, the taxon will represent a so-called relict group (of animals or plants).
If we -- in our theory -- also want to account for evolutionary extinction and the presence of relict groups, as they definitively occur in organic evolution, and if we want to further consolidate results obtained earlier with respect to evolution, we need to r e v i s e the concept of Substance (in its metaphysical sense) as it was, a long time ago, given by Aristotle and St. Thomas Aquinas and then further worked out in First Part of the present website.
A new interpretation of Substance.
While in the mentioned earlier discussions about Substance in the metaphysical sense (namely as intrinsic being) (First part of Website) we had acknowledged Reality to consist of only one ontological domain, the space-time world, we now have assumed Reality to consist of two interconnected and, indeed, interacting, ontologically different domains, viz., the Explicate Order (space-time domain) and the Implicate Order (noëtic domain). Taking this ontological duality of Reality for granted, we will now accordingly revise our earlier concept of Substance.
To begin with, any given genuine substance (inorganic or organic) in the metaphysical sense stands with one foot in the Explicate Order and with the other foot in the Implicate Order. And these Orders are in constant communication with each other as a result of the processes of injection (enfoldment) and projection (unfoldment). So one and the same given substance (where "the same" here means "having the same intrinsic whatness") can be seen in both Orders, which does not, however, mean that we have two substances. The one given substance represents itself differently in different Orders (that is, in the Explicate Order and in the Implicate Order). In the Implicate Order the given substance is only encountered as the noëtic formal content of its Essence, that is, this noëtic formal content represents the intrinsic whatness of that substance. In the Explicate Order, on the other hand, that same substance is only encountered in the form of material individuals. Also here the substance's Essence is the mentioned particular formal content. But this formal content is here not encountered as existing on its own (like it is in the Implicate Order), but always existing in each material individual of the species representing the given substance. After having observed and analyzed these individuals with respect to common features we might obtain an idea as to what their specific Essence is, that is, not the essence qua individual but qua species. This Essence is, it is true, the essence of one or another individual pointed to with the finger, but only insofar as this individual represents -- as does any other con-specific individual -- the particular substance in question.
Formerly we considered any given genuine substance to be a definite material individual thing -- like a single crystal, or a single organism -- to which we can point with the finger. Any other material individual thing, harbouring, however, exactly the same specific Essence, was considered to be another substance but "another" only in the sense that it is encountered not here but there (or not now but then). Individuality was considered to be an intrinsic feature of something being a substance.
Now -- that is, in the context of the present theory -- we consider a substance a bit differently : First of all we consider a given substance not anymore as one or another existing individual material thing, but as the total of all individuals representing it. Indeed, existing as a collection of material individuals is the very way we encounter the one (given) substance in the Explicate Order. As such this collection of material individuals results from the unfoldment, or, equivalently, the projection, of one definite noëtic formal content from the Implicate Order into the Explicate Order, and this noëtic formal content is in turn the result of enfoldment, or, equivalently, injection -- from the Explicate Order into the Implicate Order -- of the formal-content-in-its-material-condition (that is, as its condition of residing in material things). So while we encounter the one substance in the Explicate Order as a collection of material individuals, we encounter it in the Implicate Order as a single noëtic formal content.
This re-interpretation of the concept of Substance is necessary for the integration of evolutionary phenomena with our general theory of the Implicate and Explicate Order, that is, it is necessary in order to provide a 'rationale' for these phenomena. In organic evolution we not only see individuals 'living their way' as they interact with their environment, but, in a way, also species, and also the higher systematic taxa such as a genus, a family, a superfamily, etc. Provided a given systematic taxon -- as an element in one of our systems of biological classification -- is correctly established, that taxon is not only some abstract element in one or another such a classificatory system, but also a genuine biological unit, that is to say, a unit with its own ' self ' and its own history and tendencies. This is evident when we observe, say, families. Thus the family of Tachinidae, made up of all entomoparasitic (two-winged) flies (comprising several thousands of species), has its own typical morphological, physiological, behavioral, and ecological features. All its representatives are internal parasites of insects (mainly caterpillars) and adult representatives of the family are easily recognized in the field. However, in the Explicate Order we do not actually see such a family acting as a unit. We only see individuals interacting with their environment (feeding, etc.). Only upon comparison of as many individuals as possible we get the impression that the family acts as a single genuine self-contained biological unit. And so also its lower-level systematic taxa such as its genera and species, provided that they are correctly delimited by science, but also, and under the same conditions, the higher-level systematic taxa such as the superfamily and infraorder. The conception that such taxa interact and evolve (concluded from existing fossil remains) as single self-contained biological units is only a conception and nothing more, and is concluded from observable features present in the individuals representing the taxon. Indeed, all we actually observe is the interaction of material individuals. This is the way we encounter taxa, that is, this is the way we encounter them in the Explicate Order. The very ground that allows us to conclude that the given collection of observed material individuals (organisms) forms a self-contained biological unit lies in the Implicate Order : There -- if our theory is correct -- we will encounter the taxon in question as a single self-contained noëtic entity or content. So the same taxon shows itself as a collection of related material individuals in the Explicate Order, while in the Implicate Order it shows itself as a single noëtic formal content. And now we see that we have discovered another type of Substance, namely a higher-order substance, that is, not even a species -- which was already more or less conceded by traditional metaphysics -- but also a genus, family, etc. Projection of this noëtic content from the Implicate Order into the Explicate Order results in the presence of observable related material individuals, each one interacting with its environment.
The fact that biological taxa are genuine self-contained units in the way described, derives from the fact that they have evolved in the sense that they have become what they are by more or less gradual complexification (or, in other cases, simplification, resulting in a relict taxon) in the course of time. On the other hand, inorganic taxa, for example ' mineralogical taxa ', if such entities truly exist, that is, inorganic taxa having higher-than-species rank, such as "silicate crystal" (of which there are many species), have not historically evolved from simpler precursors, and so they remain just abstract categories of some classificatory system, while a single crystal, and at most its species, is a self-contained inorganic unit, that is, an inorganic substance (in the metaphysical sense).
Indeed, we have now discovered substances which are the product of some evolution and which can, in principle evolve and diversify further or become extinct or become a relict taxon. From its early beginnings onward the higher, that is, higher-ranking, taxon evolutionarily generally enriches its intrinsic formal content bit by bit, that is, the substance enriches its Essence as time goes by, or its Essence is impoverished in the case of a taxon becoming a relict taxon. The impoverishment of a taxon, and its consequent transformation into a relict taxon, can be understood by the ongoing extinction of more and more of its species, visible in the Explicate Order as the disappearance of more and more individuals belonging to this taxon, and 'visible' in the Implicate Order as a simplification of the noëtic formal content of the taxon, that is, its gradually becoming more and more monomorphous. The resulting relict taxon now consists of only a few species, the individuals of which are highly adapted to some environment of refuge but in addition still possess some ancient features. We do not consider such a degradation of a whole taxon (not to be confused with the evolutionary degradation or complete reduction of one or another organ system taking place in a given group of organisms) as evolution, at least not as evolution in its proper sense. We will consider genuine organic evolution to be a process of structural complexification (which could include local simplifications) and qualitative enrichment.
Evolution (enrichment and complexification) of a higher-ranking t a x o n as this evolution is seen in the Explicate Order and in the Implicate Order.
In the Explicate Order we see the enrichment of the taxon as follows : Its individuals (organisms) interact with their regular environment and a number of them invades other environments. For some of these new environments special adaptations are evolutionarily developed. Other environments turn out not to contain the proper conditions sustaining existence of these individuals. Again new environmental types are invaded and new adaptations are developed. As a result the number of different ecological types in the taxon, we could call them species of the taxon, increases, that is, the internal diversity of the taxon increases ( Science cannot, of course, actually observe evolutionary processes, such as the step by step development of an at least major adaptation to a new environment, but can conclude that such processes have taken place, or are still taking place, in the evolutionary life-lines of the organisms under investigation, and science can also conclude that such processes can take thousands or even millions of years).
In the Implicate Order, on the other hand, we 'see' the enrichment of the taxon as follows : The one noëtic formal content of the taxon is initially rather simple and monomorphous (where "initially" refers to the point in time in the Explicate Order at which we ' look ' down into the Implicate Order [as explained above] ). In the course of time (time in the Explicate Order) this noëtic content, as residing in the Implicate Order, becomes more and more complex as a result of inclusion and assimilation of newly injected noëtic contents, or, we can say, as a result of noëtic reactions.
Continued discussion of the origin and evolution of a higher-ranking taxon.
We assume that every genuine (higher-ranking) taxon is not only a mere abstract category in some classificatory system set up by science, but is, when correctly recognized, a real biological unit consisting of a number of certain organic individuals.
We further assume that such a taxon is in all cases a so-called monophyletic group of organisms. While the ultimate elements -- the 'atoms' -- of a taxon are its constituent individual organisms, we can, in the process of descending to the elements of the taxon, legitimately stop at its constituent species rather than its individuals, and consider them, that is, its species, as the elements of the taxon, because science is, within limits, fairly able to theoretically define what a biological species in fact is, and verify it empirically for individual cases. And it is indeed species -- and not individuals -- of which the formal content is injected into the Implicate Order.
Well, a taxon, that is, a monophyletic group, consists of a number of species, say the species A, B, C, D, and E, that have one common ancestral species, and this latter species is the ancestral species of only the species A, B, C, D, and E, and thus not of other species. Of course there are also monophyletic groups such that each of them consists of several smaller monophyletic groups (each in turn consisting of species) having - as a group of (smaller) monophyletic groups -- one common ancestral species. So the constituent species (or groups of them) are related to each other by having a common ancestor. As to the further nature of this "being related " we will not speculate, although generally it is assumed to be genetic. See next Figure.
Because the different species of a taxon have desended from a definite ancestral species, the respective formal content (as residing in corresponding individuals) of each of these descended species must contain some residue of the formal content of the ancestral species. Nevertheless, the taxon consists of spatially separated descended individuals and of the individuals of its ancestral species which were separated from each other spatially and are separated in time from the decended species. So the formal content as residing in individual organisms of the descended species are, in the Explicate Order, not only separated from each other, and here spatially so, but also from that of their ancestral species, and here the separation is in time.
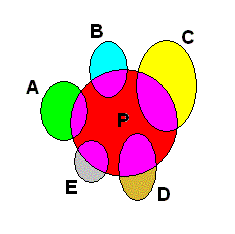
Figure 2. The same monophyletic group as in Figure 1, but now as 'seen' in the Implicate Order. The group -- taxon -- consists of the species A, B, C, D and E, and its (unique) ancestral species P, and is represented in the Implicate Order as a cluster of noëtic contents which partly overlap qualitatively (that is, noëtically) : Each formal content of the descended species contains a part of the formal content of the ancestral species.
So the first stage of complexification (evolution) of our taxon consists in the splitting up of the ancestral species (P) into five species (A, B, C, D, and E). As seen in the Explicate Order we, as investigators, conclude that all individuals (organisms), which are encountered as spatially separated from each other, and turned out to belong to five species, have descended from formerly existing individuals of an ancestral species. In the Implicate Order, on the other hand, we see a complexification of an initial noëtic formal content, resulting in one more complex noëtic formal content :
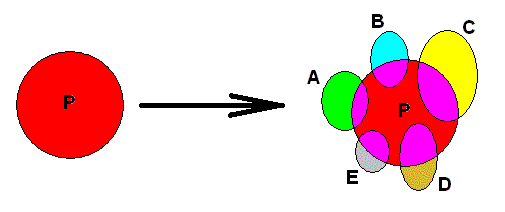
Figure 3. First step in the evolution of the taxon depicted in Figures 1 and 2, as seen in the Implicate Order. The initial noëtic formal content complexifies, but remains one single noëtic content or entity.
Although this way of drawing things, that is, expressing partly noëtic identity by partly overlapping circles or ellipses, is perfectly correct, it becoms inconvenient when still more complex noëtic contents are to be expressed this way. Therefore we go back to Figure 1. As it stands the diagram just depicts the splitting up of one species into five as a process that has taken place in time, that is, along the time dimension. If, on the other hand, we want a slightly different version of this type of diagram to express the taxon in its form of the noëtic unity, in which form it resides in the Implicate Order, we shall interpret the lines connecting the ancestral species P with each of the species, A, B, C, D, and E, having directly descended from it, as expressing the partial noëtic overlap between the noëtic formal content of the ancestral species and those contents of the descended species. In contrast to Figure 1 the lines will now not only continuously connect the formal contents but be part of it, and in this way expressing the partly overlap :
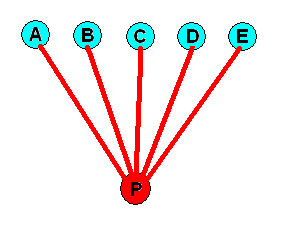
Figure 4. The taxon (P, A, B, C, D, E) of the previous Figures, depicted as it is seen in the Implicate Order : Here the taxon is present as one single noëtic entity or content. The lines connecting the noëtic formal content of the ancestral species with those of the descended species express the partly overlap of the respective noëtic contents. Moreover, the diagram, while depicting the situation in the timeless and spaceless Implicate Order, refers to the time dimension as this will be added when expressing the taxon as it is seen in the Explicate Order : The original species has, in the course of time, subdivided into five descendants.
So in this way we express the taxon as it, as a single noëtic content, resides in the Implicate Order, and at the same time express it as it is seen as a set of species consisting of an ancestral species and species descended from it, that is, expressing it in the form by which it is encountered in the Explicate Order. Of course the only thing we actually see in the Explicate Order are individuals, that is, individual organisms. That they represent species is not directly observed but concluded from observed features. But because physiological appropriateness to interbreed, which is the main criterion of conspecifity of certain given organic individuals, is a real (in contrast to purely theoretical) feature that can in principle directly be observed, we consider species (provided they are correctly established as such by science) as (albeit indirectly) observable entities, that is, entities that can and will as such be encountered in the Explicate Order. But, it must again be stressed, that they are not encountered there as agents, that is as entities that interact with their environment or with each other. In the Explicate Order it is only individual organisms that so interact. Only as seen in the Implicate Order species can interact with each other or with environmental features, but here those interactions are in fact purely noëtic reactions.
In the course of evolution the taxon generally further complexifies by subdividing into yet more different species often involving the above described internalization of a part of the external extent into the internal content of a given species. See next Figure.
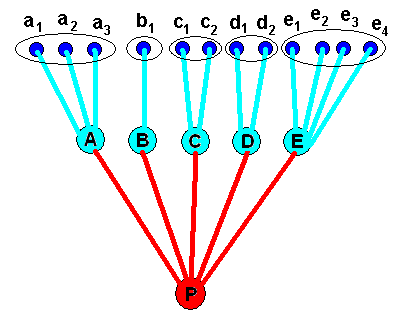
Figure 5. The taxon (P, A, B, C, D, E) has complexified further by subdividing into more different species. Five subtaxa have now appeared : ( A, a1 , a2 , a3 ), ( B, b1 ), ( C, c1 , c2 ), ( D, d1 , d2 ), ( E, e1 , e2 , e3 , e4 ). Also each subtaxon is, as seen in the Implicate Order, a single noëtic unit in the following way : First of all it contains the complete formal content of each of its latest species, and in addition to that it contains part of the formal content of its direct ancestral species and also part of the formal content of its remote ancestral species (P). For example, the subtaxon ( A, a1 , a2 , a3 ) contains the complete formal contents of the species a1 , a2 , and a3 , further (it contains) a part of the formal content of the direct ancestral species A, and, finally, it also contains a part of the formal content of the remote ancestral species P (and of all other still farther remote ancestral species).
The whole taxon can now be written as :
( P, A, B, C, D, E, a1 , a2 , a3 , b1 , c1 , c2 , d1 , d2 , e1 , e2 , e3 , e4 ).
And of course this complexification can go on, albeit not indefinitely.
All the above diagrams, depicting a (given) taxon, only very generally and schematically express the complexity of its formal content. They do not at all express the magnitude of the differences between its species and subtaxa, neither do they express the qualitative nature of these differences and with it the qualitative content of each species and each subtaxon.
The relation of a given animal species with its environment.
Any given animal species consists of one or more populations of individuals, that is, individual animals belonging to that species. When it is said that given individual animals belong to one and the same species we mean that they can in principle interbreed, apart from the fact that they often do not, as in cases where the individuals belong to geographically separated populations, that is, populations separated by areas that cannot be crossed over by their individuals. The interbreeding criterion is meant to be ultimately physiological in nature. And this criterion of "animal individuals belonging to one and the same species" -- their conspecificity -- should not be understood as a rigid condition, but as a first datum that could play a role in determining whether given animal individuals belong to the same species or not. Some, probably rare, cases of conspecifity of given animal individuals, divided over two populations, must perhaps be conceded, in spite of the established fact (in such a case) that interbreeding has turned out not to be (physiologically) possible, or maybe such a case is evidence that we witness the fact that the initially conspecific populations are in the process of splitting up into two different species.
The environment, say the environment E, in which a given animal species, say the species S, lives, that is, in which that species' individuals as a rule live, is a certain type of 'terrain' (terrestrial or aquatic) regularly shaped by (1) prevailing abiotic factors, for instance macroclimatic conditions such as temperature (and temperature fluctuations), moisture (and with respect to the air, humidity), and (amount of average daily) sunshine, and (2) (the terrain is regularly shaped by) prevailing general biotic factors such as the presence, and if so, the density and nature, of the plant cover, and all the animals (as to their species composition and densities) to which it offers a suitable habitat. So this terrain is an environment or habitat not only for the just mentioned animals but also for the individuals of our species S.
But within every (type of) environment, and thus also within this one particular environment or habitat E, every resident animal species has its own special place : Our environment E consists of a number of qualitatively different ecological niches, and each resident species, that is, each animal species that regularly lives in environment E, occupies within it a particular ecological niche to which it is especially adapted. This "being adapted " of the (individuals of the) animal species to its ecological niche boils down to the possession by each animal individual of an ensemble of a number of specific adaptational structures which can be morphological, physiological, or behavioral (such as special instincts). In fact, as we had said earlier, such an adaptation is a kind of "internalization", that is, a certain representation of a part of the organism's environment in that organism. So the organism is qualitatively connected with precisely that part of its environment or habitat that forms the organism's ecological niche. This ecological niche is physically external to the organism, but still at the same time also internally represented in it, namely by the organism's ensemble of adaptational structures.
As seen in the Explicate Order the evolutionary development of the ensemble of adaptations of the indidividuals of a given animal species to a certain ecological niche is the projection or unfoldment from the Implicate Order of the product of a noëtic reaction having taken place in the Implicate Order between the formal noëtic content of the animal species and the formal noëtic content of the ecological niche, which initially was one of the species' potential ecological niches. All this was already expounded earlier.
A higher taxon consists of, generally, many related species, and individuals of these species might find themselves at some time in another part of the species' environment than the one they are adapted to (their ecological niche). This could be the result of them having arrived at, or ended up in, that part, or it could be the result of a change of that part of the environment which was their ecological niche. And if this new part of the environment happens to be a potential niche of the species (that is, an environmental part that does not, by its nature, beforehand deny any possible existence of the mentioned individuals of the species), it is then by itself a genuine content because it is precisely defined and delimited by the species' formal content and will be noëtically present in the Implicate Order. It is then able to noëtically react with the injected formal content of the animal species in question, resulting in a noëtic reaction product which, upon projection (unfolding), is seen in the Explicate Order as the evolutionary appearance of one or more adaptive structures. Normally we see these structures appearing in individuals of one of the several existing populations of the species (and not in the other ones), and eventually this will lead to the splitting up of the original species into two (or more) : speciation.
As a number of species of the given taxon undergoes the just described process of speciation, the taxon, as a result, complexifies : It 'stretches out' its 'tentacles' into evermore new ecological niches. It cannot, however, represent itself (by anyone of its species) in any type of environment or ecological niche whatsoever : Its central intrinsic nature sets limits as to in what types of environment and ecological niche its individuals can live, that is, its intrinsic nature sets limits as to what kinds of adaptational structures can be evolutionarily developed in the individuals of the taxon's species. And often, many of the species' potential ecological niches can turn out to be already occupied by animals of other, albeit closely related, species. It also happens that a particular species of the taxon, that has, in the course of the taxon's evolution, specialized itself to occupy a particular new ecological niche, later (in evolution) returns to the ecological niche occupied by one of the taxon's (evolutionarily) early species, or, which is more probable, to a niche very close to that niche. In this way such a species can also come to occupy an ecological position very close to that of a species of another (but related) taxon. More broadly this means that the species settles down into the same environment as is inhabited by the other species. So a higher-level taxon is rather ecologically flexible, but always within certain limits set by its central intrinsic nature (which is the nature of its ancestral species).
Intermezzo on environment and ecological niche (of a given animal species).
Here it is important to clearly distinguish between, on the one hand, the environment or habitat of a given animal species, and its ecological niche on the other. The environment or habitat of a given animal species is the type of terrestrial or aquatic 'terrain' its individuals as a rule live in, and which is shaped by a number of gross abiotic and biotic factors. Such environments are, for example, (the zone of) coniferous forest, tundra, tropical rainforest, open grassy landscape, mud flat, more or less stagnant small fresh water reservoirs, mountain streams (fast-flowing cold water), larger lakes, the banks or shores of these aquatic environments, desert landscapes, seashores, the pelagic zone of the oceans, the bottom of oceans, etc. But each of such an environment can, of course, harbour a great many different animal species. And the individuals of the different species living in one and the same environment do not compete there for the same food resources, egg-laying sites, or whatever (think, with respect to feeding, of the many species of mud-flat birds that feed in the same environment but are adapted to different sorts of food as a result of the different lengths of their beaks). They seem to have their own place within that particular environment, that is, the individuals of one given animal species occupy a strictly determined type of site in the particular environment or habitat, while the individuals of another animal species, also living in this environment, occupy another, but also strictly determined, type of site in this very same environment. Such a determined type of site in a given environment (which, also contains sites of other types) is a certain aspect of that environment, or, maybe better expressed, it is a certain aspect of some given animal species whose individuals live in that environment, that is, a certain extension of the phenotype of these animals into the environment and in this way comprising or seizing the mentioned aspect of the environment. This particular site of the environment is biochemically fully compatible with the physiology (including physiological chemistry) of feeding, digestion, exchange, locomotion, etc, that is intrinsic to a given animal species occupying this particular site of the environment. And the nature of the physiological correspondence between a given animal species and this site is so specific that the individuals of that animal species connected with this site do not in any way interfere with other animal species living in that same environment, because these latter species are connected with another site in it. And such a site is called an ecological niche, and it is always an ecological niche for a certain well-defined animal species. Only individuals of this species can occupy that particular ecological niche.
( End of Intermezzo )
And thus in this way (as expounded above) we see the evolution of animals (but the same goes for plants) as driven, not by random genetic mutations and the natural selection of those which increase the survival value of the animals (this process does occur in the organic world, it is true, but is not the overall driving force of organic evolution), but by ecological expansion of (higher) taxa. Of course a new environment (or changed environment) will change selection pressures, but it cannot be held that it is necessarily accompanied by corresponding advantageous genetic mutations occurring at the right time and in the right order, because most genetic mutations are either not phenotypically expressed or are disadvantageous or even lethal. The fact that complex 'smart' adaptations of organisms to new environments or new ecological niches have occurred abundantly and seem to defy reductionistic explanation even in principle, has led us to theorize about the noëtic background of material reality and especially of biological reality.
Any biologically correctly conceived taxon, also any of the higher-ranking ones, is, by our theory, considered to be a substance in the metaphysical sense, in addition to already acknowledged substances such as individual crystals and individual organisms. But the taxon is a substance of an altogether different order than individual crystals or organisms are. It does not represent itself in the Explicate Order as a single uniform individual thing -- as crystals and organisms do -- but as a collection of related, but spatially separated, individual organisms. And because the latter are substances themselves our taxon is a supra-individual substance (high-level substance), and can contain a number of lower-level, but still supra-individual, substances -- subtaxa. And, as has been said, while the one taxon is in the Explicate Order present in the form of a smaller or larger number of physically separated individuals, in the Implicate Order it is one single noëtic entity. Further, while the constituent species of the taxon split up into more and more different species, occupying more and more different ecological niches, the taxon complexifies, that is, the taxon evolves. Indeed, the taxon is a substance that can, and will, evolve. The historically initial species of the taxon (always considered as a monophyletic group), that is, the one species that is ancestral to all the other species of the taxon, sets the limits for the taxon's future evolutionary development. Indeed, the ancestral species, as containing the central intrinsic nature of the taxon, determines the general direction of the taxon's evolutionary development, which here means its ecological diversification along certain more or less definite pathways. But, as it is important to note, the nature of the ancestral species does not contain, or determine, the complete evolutionary development of the taxon, but only sets certain limits under which this development is allowed to take place. It does not abolish the aspect of genuine creativity in evolutionary development.
Further characterization of a biological (higher-level) taxon as a s u b s t a n c e in the metaphysical sense.
A genuine substance is a true 'something', that is, a something that in a way possesses some kind of 'self ' (in a broad general sense). This implies that it allows to distinguish between intrinsic and extrinsic features with respect to it. If we consider, to begin with, a given inorganic substance, for example an individual snow crystal, we can distinguish features intrinsic to it, for instance its total symmetry, that is, its space group, and its chemical composition. When the snow crystal has grown under uniform and mild conditions (of temperature and humidity of the air) and has been allowed to grow feely, that is, not in close material contact with other snow crystals or other solid objects, it develops into a neat hexagonal column. This repeatable and stable shape of the crystal is a direct consequence of its internal structure and symmetry and thus is an intrinsic feature of ice crystals in general (crystals of the compound H2O), while the shapes of all the many 'distorted' unsymmetrical snow crystals which we often encounter in a given snowfall are extrinsic features of ice crystals in general. They are caused, not by internal factors such as the structure of the ice-crystal lattice, but by external factors, external with respect to the snow crystal. Of course the very way the snow crystal reacts to those external influences is intrinsic, but the resulting features such as its shape (expressed for example in the irregular branching of the crystal) must be evaluated as extrinsic because they are caused by factors external to the crystal and its intrinsic structure.
The same can be said of a given organic individual. Also such an individual organism is a genuine substance in the metaphysical sense. A human being for example can become tanned. But this is caused by having exposed himself for a while by strong solar radiation, which contains ultraviolet rays. Also here the way that his skin reacts to this radiation, namely by the formation of a brown pigment, is intrinsic for at least the white man, but, according to the way it is caused the brown color of parts of his skin is an extrinsic feature. And, as another example, when a given man is fat, his fatness is an extrinsic feature, extrinsic that is to say, to the human species, which is clear from the fact that not all humans are fat.
By the way, when we speak, in the present context, about the possession by a given substance of a 'self ', this should not be confused with the 'self ' of the human (or animal) individual (the presence of such a 'self ' we will not deny). In our new conception of substance (in the metaphysical sense) the individual only represents a particular substance (the human species [different from the other animal species], the species of ice crystal [different from other species of crystals, say from salt crystals). And when we speak about posssessing a 'self ' we here mean that the substance (represented by individuals) possesses a kind of 'self '.
And, as has been said, possession of a 'self ' implies the distiction between intrinsic and extrinsic features of the substance in question.
However, if we want to apply this distinction to the case of a biological higher-level t a x o n -- which we now also ontologically assess to be a substance -- things become more difficult. As has been said, a biological higher-level taxon (such as a family or infraorder of animals) is a substance (which, as every substance does, stands with one foot in the Explicate Order and with the other in the Implicate Order) that, in contrast to inorganic substances, has evolutionarily evolved and will, if conditions permit, evolve still further. The evolution of the taxon consists, as has been said, in its ecological diversification as a result of the evolutionarily generation of new species of it which occupy, as a result of the development of special adaptations, new ecological niches. And because ecological niches are aspects of concrete environments such as woodland, lake, desert, etc., they are as such external to the one animal species of which we discuss such an environment as being its habitat and of which we discuss the special ecological niche in that environment that is occupied by that particular animal species. Indeed, that part or aspect of the environment which, earlier in time, was to become the ecological niche of the new animal species, has appeared as a result of certain changes of the overall environment, and these latter changes are clearly extrinsic to that given (new) animal species. Consequently, the special adaptations (morphological, physiological, behavioral) acquired by (individuals of) that species in order to be able to occupy that particular niche have an external cause and should consequently be assessed as extrinsic features of that -- new -- species, and by implication assessed as extrinsic features of the higher-level taxon to which this new species (and its direct ancestral species) belong. However, it is problematic to assess members (species or subtaxa) of a given taxon as mere extrinsic features of that taxon, because it truly seems that they intrinsically belong to the formal content of the taxon so far evolved, despite the fact that the composition of the taxon (i.e. its being composed of species and subtaxa) in a way reflects the environmental history of the geographical regions where the individuals of the taxon live, which history seems to be extrinsic to the taxon. And indeed, this history is not determined by it, but by factors external to it. So this is the problem we must solve. And it can be solved. But the solution consists of no more than drawing the full consequence of the assumption we have made, namely the assumption that every biological taxon -- as presumed to be a substance -- is intrinsically evolutionary, that is, the higher-levelness, if present, of any given biological taxon -- i.e. the taxon consisting of a number of subtaxa -- is the result of an evolutionary process. Biologically seen there is little new in this assumption. In its form of the taxon being enriched by new subtaxa as a result of evolutionary processes it is not questioned by anybody. But in its metaphysical form, namely that the taxon is a substance, and moreover a substance that can evolve, here meaning that although its intrinsic content changes -- evolutionarily develops -- it nevertheless remains the same substance, it is a very hypothetical assumption, but, we would like to stress, compelled to make as a result of evaluating (or trying to understand) observed phenomena in organic evolution. And every taxon, now assumed to be an evolutionary substance, will, so long as conditions permit, continue to evolve. It will acquire more and more subtaxa as a result of the formation of new species in it, species adapted to new ecological niches.
The following statement will now express how we should interpret the status of the new content being added to that of the taxon, that is, whether this newly added content is, like the original content of the taxon, intrinsic to it, or (whether it is) not.
Every new adaptation to a new ecological niche (a new adaptation) that is developing in one of the taxon's species, eventually leading to a new species being added to the taxon, boils down to a little piece of new formal content being added to the formal content of the taxon, being added in such a way that it now has become an integral part of the (new) formal content or intrinsic nature of the taxon, regardless of the extrinsic (with respect to the taxon's initial formal essence) nature of the ultimate cause of the addition of new content.
The ultimate cause of the content of a taxon getting expanded by a piece of new content often can be traced back to a change of the overall terrain (inhabited by the species in question of the taxon) as a result of, say, climatic changes or due to the advent of other animal species from elsewhere which either constitute a new type of predators of the individuals of one of the taxon's species or themselves happen to constitute new sources of food for these individuals. And it is clear that such (ultimate) causes are totally extrinsic to the essence or nature of the taxon which experiences such an extension of its content. So independently of the status -- intrinsic or extrinsic -- of its ultimate cause, every subtaxon of the taxon is an intrinsic, that is, per se, part of it.
The formation of a new species generally gives rise to a new subtaxon, because it is the result of one original species having divided into generally two species (where one of them is often not significantly changed with repect to the original species, while the other is), and these two species together constitute the new subtaxon. Further subdivisions of these species will lead to a higher-ranking subtaxon.
We have given to the biological taxon the ontological status of being a substance. And the question of the per se has now been settled. But if certain features are definitely distinguished as to belong per se (that is, intrinsically) to a given taxon (seen as a substance), there must be other features that belong merely per accidens ( = accidentally), that is, extrinsically, to that taxon (still seen as a substance).
Do such features really exist? And if so, where? Well, we can confidently maintain that all of the accidental can only be found at the level of individuals. Indeed, in individuals (representing a substance) we can recognize per accidens features, that is features not belonging intrinsically to the substance that is represented by the individuals. Above we have given some examples (distorted shapes of crystals, being-tanned in humans, being-fat in humans). So per accidens features of a given taxon can be found, and can only be found, in the individuals representing that taxon. And from this it follows that such per accidens features cannot -- precisely in the form of per accidens features -- exist in the Implicate Order, because there individuals (spatially separated things) cannot exist.
The reason that a taxon is, in contrast to any inorganic substance, intrinsically an evolutionary substance must be due to the fact that any organism and with it its species (which is the starting point for the evolutionary development of a higher-level taxon) is intricately connected with its environment or habitat and especially with its ecological niche in that environment, while inorganic substances, such as crystals are not. A crystal, it is true, also needs a particular environment in order for it to emerge and be substained, but this environment is in fact just a (1) chemical condition, that is, the presence of a certain chemical element or compound saturating a solution or just being present in sufficient quantity, and (2) certain physical parameters (like temperature and pressure) lying within a certain range. There are no ecological niches in such an environment for crystals, that is, for special sorts of crystal to exist. The connection of the crystal with its environment is rather loose, that is to say, the crystal barely internalizes parts of its environment. The latter is only its conditio sine qua non, that is, an absolutely necessary condional field (comprising a determined range of concentrations, temperatures and pressures) for the crystal to emerge. But despite its being necessary this conditional field cannot be interpreted as some sort of qualitative extension of the crystal's intrinsic nature or formal content. It does not add to this formal content. The crystal will never venture into another environment, and when it is subjected to a change in the environment that brings the latter beyond the mentioned ranges of the chemical and physical parameters, the crystal will become unstable and will disintegrate, or at most metastable (that is, there is a certain smaller or larger energy barrier that has to be taken before the crystal disintegrates). The crystal will never adapt to such a new environment. And consequently the crystal will not evolve.
Organisms, on the other hand, internalize parts or aspects of their present environment, and can, within limits, adapt to a new environment or to a new (later to be called) ecological niche by internalizing certain aspects of this new environment. Possessing this ability to adapt even to significanly different environments (harboring different potential ecological niches) it can, as a life form, that is as an initial species, evolve, eventually resulting in the course of its evolution in a large and internally differentiated biological taxon.
The evolving taxon.
As has been said earlier, we consider every biological taxon to be monophyletic, which means that the taxon has one ancestral species from which all of its subsequent species have descended : the ancestral species divides into two or more species each one of them in turn divides into two or more species. And this can go on indefinitely, resulting in many subtaxa of the taxon. The formation of new species implied by these subdivisions is caused by, so to say, the 'exploration' of the taxon of different environments and the occupation of more and more different ecological niches by newly developed species of the taxon, where this occupation of new ecological niches is made possible by the evolutionary development of special adaptations. The formal content of the taxon's ancestral species sets limits as to what ecological niches can be occupied, and this is visible in the 'general evolutionary trend' of the taxon. The occupation of a new ecological niche by one of the taxon's new species is in fact the internalization of this niche (or of parts of it) by the organism, that is, by the members of the species. This internalization process consists of the evolutionary formation of special adaptations of the organism to that new ecological niche, which (niche) in this way is, morphologically, physiologically, or behaviorally, reflected -- internalized -- in the organism. In the Explicate Order we do not actually see the taxon-as-a-whole exploring new environments. The only thing that we see is individual organisms interacting with individual elements of their environment. Only in the Implicate Order we 'see' this same taxon as one noëtic unit noëtically reacting with noëtic formal contents representing aspects of some environment, and the projection of the reaction products is then seen in the Explicate Order as the evolutionary development of adaptations and the origin of new species occupying new ecological niches. All this has been expounded already earlier.
Let us try to graphically and schematically express what we see in the Explicate Order when a given taxon is evolving in geological time.
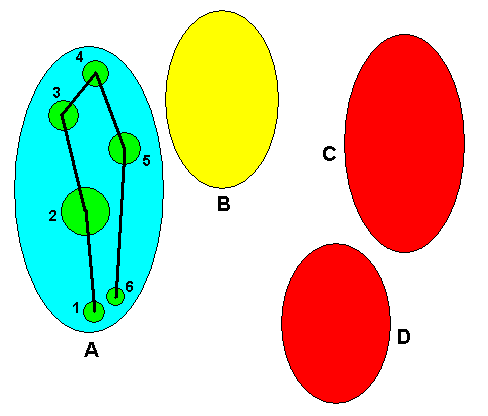
Figure 6 : A given taxon has originated in environment A, that is, its ancestral species occupies ecological niche 1 in this environment. After this, the environment A is further colonized by newly developed species of the taxon, resulting in the successive occupation of ecological niches in this environment : 2 ==> 3==> 4 ==> 5 ==> 6.
Environment B is not yet colonized by species of the taxon, while the environments C and D are supposed to be such that they can never be colonized by any (present or future) species of the taxon because the nature of this taxon is incompatible with these environments.
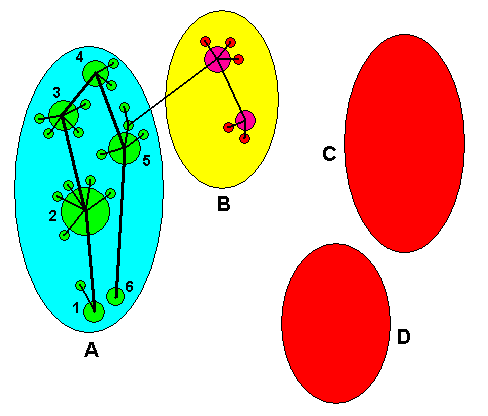
Figure 7 : This diagam depicts the splitting up of some of the new species of the previous diagram into yet more new species each one of them occupying an ecological niche slightly different from the original ecological niche, but still part of environment A. In this way real subtaxa of the taxon are developed. One of the descendant species has split up into two species, one of which -- a new species -- has adapted to an ecological niche in a new environment : this ecological niche being a part or aspect of environment B . It then has split up further as a result of adaptations to several new ecological niches in that same environment. Those species of our taxon now inhabiting environment B together form a higher-ranking subtaxon.
As it is clear that not all environments can be colonized by a given taxon, it should also be clear that not all parts, places or aspects of one and the same environment can be occupied by (present or future) members of the taxon, that is, not all such parts can become ecological niches of (present or future) species of this taxon.
The above two diagrams symbolically depict several different environments, that is, different types of environment, such as woodland, seaboards, rivers, open grasslands, sea, mountain range, etc. These should not be confused with geographical regions. Most geographical regions contain several types of environment. Also the indicated ecological niches are not equivalent to certain geographical regions. So a given ecological niche could be simultaneously present in several geographical regions. Geographical regions are spatial entities, while environments and ecological niches are purely biologically determined. An ecological niche of a given animal species is a definite aspect of an environment, which (niche) is, by way of adaptations, internalized into the individual organisms that have come to possess these adaptations.
The evolutionary history of a given taxon is, even in its symbolic and general form as in the diagrams, not completely depicted or symbolized. This history is also reflected by the systematic content of the taxon in its present condition : the total number of its species, the way how these species are distributed over the different subtaxa, and the number of species of each subtaxon.
And whether the taxon, or one or more of its subtaxa, is a relict group, that is a group (taxon, subtaxon, or species) that is presently on the decline, but which was florishing during earlier times, can be seen (1) by the mixture of (utterly) specialized and primitive (i.e. original) features of the body, the physiology, the individual development, etc., and (2) by the occupation by such a group of an ecological niche in a very extreme and sparsely populated environment such as high mountains or deep sea, and, (3) by the extraordinary abundance and diversity of fossil members of the group (if we are so lucky to find these fossils), and, finally, (4) by the small number and little diversity of species of the group living at present.
On the other hand, a presently (still) flourishing taxon can be recognized by its large number of recent species, its spread over many types of environments, and its global or almost global geographical distribution. And, of course, every relict group once was such a flourishing taxon.
We have described the evolution of a given taxon as consisting of the increase of the number of its species as a result of more and more completely colonizing a certain type of environment and even the colonizing of one or more other types of environment. Each species of the taxon then occupies a specific ecological niche in one of these environments.
But apart from the case of the biological establisment (in Nature) of a taxon in very ancient times (that is, in times in which lived its ancestral species and the latter's immediate descendants), times so ancient that many types or subtypes of terrestrial environments, such as environments where angiosperms (higher plants, that is, plants higher than conifers, ferns, and the like) are dominating the scene, did not yet even exist on the Earth's surface, it is hard to see why the expansion of the taxon's habitat, either within one and the same environmental type or including other such types, should be the sole constructional force generating new and different species -- new and different ecological types -- of the taxon. We can confidently assume that from Jurassic times onward most, if not all, types of terrestrial environments were already existing, not of course necessarily at the same geographic locations, but existing (somewhere) nontheless ( The whole set of types of marine environments was, as we can be sure, existing already much longer). So when, reckoned from this time onward, some given taxon enters, with one of its species, a 'new' environment, it is an environment that is new for the taxon, not for the Earth's surface itself. But the fact of an environment being new for the taxon can still result in special adaptations, new species, and consequently, new ecological types, in the taxon. Nevertheless there must soon come a time (somewhere in the geological past) in which all aspects of the environment that could potentially become ecological niches for species of the taxon have actually become such niches. All remaining aspects of that environment are then incompatible with the intrinsic nature of the taxon -- they can never become ecological niches of present or future species of the taxon, and that means that the environment cannot be further colonized by it. And when it was the case that other types of environments were equally incompatible with the taxon's intrinsic essence, we will expect that no further evolution of the taxon will take place anymore, that is, we, in the present time, still encounter this taxon precisely as it was a long time ago, which means that it still consists of the same species as it did at that time. However, the study of fossil material often shows otherwise. With respect to insects, for example, we can say that -- judging from the fossil record -- while Tertiary faunas (dating from some 40 million years ago) look quite the same as the recent ones, Jurrassic insect faunas already are different, not to mention the Triassic, Permian, and Carboniferous insect faunas.
So, in addition to the adaptation and further specialization of animals to live in special aspects of some given environment, there must be other driving forces of evolution, resulting in recent animal forms that differ from their ancestors. This conclusion is also corroborated by the remarkable fact that many fossil animals, and in this respect certainly many fossil insects, even dating from very ancient times such as the Jurassic, show many specialized features already, and, moreover, sometimes possess features that are even in a more advanced specialized state than those of comparable recent animals are. Because this is so often the case one is forced to trace the ancestors of many recent animal groups further and further back into the geological past. Further it is well known that many recent animal forms that have remained relatively primitive with respect to their organization (morphology, physiology, behavior) still seem to do well at present times : All the lower orders of insects, even the original wingless ones (among which the so-called silver-fishes) are still important elements of the recent fauna, and the oceans abound with primitive animals, like polyps. So while we see in evolution the continuous sophistication of adaptations (bringing with it for example the so-called evolutionary 'arms races' between preditors and their preys), animals with more or less primitive adaptations seen to do just as well.
On the other hand, the occupation of newer and newer ecological niches by species of a given taxon could still be the sole factor of evolution if the number of possible ecological niches in a given environment turns out to be virtually infinite (this because any ecological niche is so subtly defined [by nature] ). And we must reckon that this could indeed be the case. Most terrestrial environments are very rich in small-scale physical and chemical gradations and patternings (abiotic components), such as lighting, moisture, temperatures. They are also rich in the more subtle biochemical gradations and patternings resulting from the presence in such an environment of a diverse biotic component which itself consists first of all of the plantcover and its associated residues, further of the microbiological content of the environment, and, finally, of the animal life that is contained in it. An animal living in a particular environment must, first of all, have a physiology that allows it to exist there at all, that is, a physiology that is compatible with the global abiotic conditions of that environment (average temperature, moisture, seasonal change, etc.). The animal must also be allowed to interact with this environment, that is, to practise its function of Exchange consisting of : feeding, excretion, and respiration. Feeding and respiration demand a chemical and physical compatibility of the animal with its environment, and especially feeding demands a subtle relationship, namely a biochemical compatibility between animal and environment.
And this is not all. The environment must allow individual development and reproduction of the animal.
In insects -- in fact the main inhabitants of terrestrial environments -- this is particularly significant because in holometabolic insects (that is, insects with a complete metamorphosis, such as butterfies, beetles, mosquitos, bees, etc.) each individual, from the egg-stage onward, passes through three consecutive developmental phases -- larva, pupa, adult -- that, generally, freely live in the environment. All three stages must be able to breathe. The pupa does not feed, and in most cases does not move about. Generally, both the larval stage(s) and the adult stage feed, but in many species the adults do not take food (aphagia). Further, where in holometabolic insects both the larva and the adult feed they take very different sorts of food, for example a larva of a mosquito feeds on microorganisms in fresh water, while the adults feed on blood of vertebrates. Larvae of other insects are feeding on rotting organic material while the adults feed on nectar which they extract from flowers. So in these cases (which are extremely abundant) the 'subenvironment' of the larva is very different from that of the corresponding adult insect. Also their morphology is totally different : Larvae of holometabolic insects (like all larvae) are wingless and generally are adapted to movement and feeding activity in certain liquid, semiliquid, or solid substrates which are or contain their food. The adults generally are winged. They fly around to seek either food (as predators or nectar feeders) or (they seek) appropriate substrates to lay their eggs on or in. But although the animal species' trophic relations (i.e. relations with respect to feeding) with elements of a given environment can be very different indeed in the different stages of individual development (larva and adult), these stages are nevertheless closely tied up to each other : whether the adult insect of a given species (always) feeds, and if it does what food it (always) takes, is dependent on the evolutionarily determined way of feeding of the larva. That is to say, there are species the larvae of which are adapted to some subenvironment in which there is plenty of food rich in nutritive substances. The adults of many such species do never take food : their mouth parts are reduced or absent. These adults only serve to reproduce and to geographically spread the species. In every respect the larval and adult stages are fully adjusted to each other. We should therefore -- in the context of theoretical discussions -- never speak about the "ecological niche of the larvae of a given animal species", or about that of the imagines ( = adult insects) for that matter, but only speak about the ecological niche of the species as a whole, that is, including all the stages of the individual development of its members.
As should be clear from the foregoing, an ecological niche of a given animal species is in fact defined biochemically, that is, as the presence of a biochemical connection or compatibility between the relevant biochemistry of the animal species, i.e. its biochemistry insofar as it 'reaches out' into the environment, and the biochemical nature of that environment insofar as it connects or interacts with that of the animal species. Such an ecological niche is always a part -- in the sense of an aspect -- of some environment, and as such it is uniquely defined by the nature of the animal species occupying that niche.
The mentioned biochemical connection between a given animal species and its environment, defining the species' ecological niche, primarily concerns food relationships (and with respect to insects : those of the larval stage(s) as well as of the adult stage, taken together). The 'penultimate' nutritive materials (i.e. materials that simply and solely need to be digested to yield the ultimate nutritive materials) needed by the individuals of the given animal species will, in many cases, be present in its ecological niche in a more or less pure state, which means that it is as such directly accessible, for example in the form of a prey, namely when the mentioned animal species is, during one or all of its developmental stages, a predator, or (is directly accessible) when the animal feeds on highly specific parts of certain plants such as their spores, pollen or fruits. But this is not always the case, that is, in many cases the penultimate materials needed by the individuals of some given animal species are enclosed within other materials, and it is then primarily these latter materials that are looked for by these individuals. But it might be possible that these latter materials not only contain the penultimate nutritive materials specifically needed by the individuals of our animal species, but also other penultimate nutritive materials specifically needed by another, closely related, animal species. What we want to say with all this is the fact that an ecological niche of a given animal species can partly o v e r l a p with that of another animal species. Of course the overlap is just a partly overlap because there exist lots of other necessary relations with the environment that are different for different animal species. Given this possibility of partly overlap, the possibility of competing for the same food material (the same 'pen-pen-ultimate nutritive material, but not the same pen-ultimate nutritive material) is automatically given. And this could mean that, as a result of such competition, one of the competing animal species, initially living in the condition of absence of the other species, is eventually driven out of its ecological niche, or (it could mean) that the adventive species could not occupy its proper ecological niche because that niche turned out to partly overlap with that of another species already present, and because it has lost the subsequent competitional struggle (which struggle, might, like in the former case, be evolutionary in nature, that is, involving the evolutionary development of special adaptations by the opponent species, concerning the ability of the latter's individuals better to find and approach their appropriate food resources, or the ability to better reproduce resulting in the presence of larger numbers of them). The species that has lost this struggle will quickly become extinct or perhaps manages to adapt to another aspect of the same environment or to an aspect of another environment.
So, with respect to the evolving taxon, what we see in the Explicate Order is the continued subdivision of its ancestral species and of the latter's descendants as a result of these species adapting to, and thus the establishment of, more and more different ecological niches in a given environment or in different environments. In this way an environment -- that is, a certain type of environment -- or even several different types of environment, is (are) colonized by the taxon. But this colonization is restricted by the competition between closely related species of that taxon or even by the competition with species of another taxon. And the latter case is expected to be much more common than the former because in it the competing species have developed phylogenetically independently of each other, that is, the one was already established elsewhere and residing in its own proper ecological niche, but later venturing out to other aspects of the environment or to a different environment altogether. When, on the other hand, two species are the direct result of the splitting-up of one ancestral species (ancestral to them) -- and in this way forming no more than a subtaxon of the taxon to which the ancestral species belonged -- they will not as a rule compete, otherwise the splitting-up of their ancestral species had not occurred at all. So we can savely maintain that one of the restrictions to which the evolutionary development of a taxon is subjected, and thus one of the factors shaping it, are ecologically similar species of other (more or less related) taxa (this in addition to all the other restrictions). So apart from predators or food resources (food plants, preys, hosts) the biotic aspects of the environment of a given animal taxon include ecologically similar species of other animal taxa. Some new ecological niches for certain species of the taxon can be established successfully, while other such niches cannot, either because they are aspects of environments totally cut off for the taxon, or because of the established presence of ecologically similar species. So the taxon must evolutionarily find its way through the existing ecological patterning of the environment or environments, and it is clear that its evolutionary development and fate is dependent on external abiotic and biotic factors, but, of course, also dependent on the typical way along which the nature of taxon -- as presently developed -- responds to these factors, which here means that the newly added contents to that of the taxon, as a result of the development of new adaptational structures in some of the taxon's species, in spite of having external causes, become intrinsic parts or aspects of the central content of the taxon. This is the way we should see the taxon 'acting as a whole' resulting in its evolution.
But this is still not the whole story. Until now we had imagined ourselves some given animal species -- in the form of one or more populations of living individuals and belonging to some higher-ranking taxon -- being adapted to a particular environment and occupying a specific ecological niche there. Then, as we have imagined further, at some time, one of its populations or a part thereof, happens to find itself in qualitatively another part of the environment or in another environment altogether, or has migrated to it. This population can or cannot adapt to the new conditions it finds itself in. And if it does indeed adapt successfully, then a new ecological type is added to the taxon, where this new ecological type generally is embodied in a new species. And so we concluded above that "the taxon must evolutionarily find its way through the existing ecological patterning of the environment or environments".
However, we cannot legitimately maintain that in the history of animals we always, that is, in every particular case, have to do at first with a more or less irregular pattern of possible ecological niches already existing at the time within some existing environmental type, and then followed by the colonization and subsequent adaptation of certain animal species. On the contrary, very often a new ecological niche for some animal species (and for the corresponding higher-ranking taxon if this species is its ancestral species) is not actually discovered by this species (or by any species for that matter) but in fact created by it. This we can, for example see at the origin of the dipterous insect family of Gall Midges (Cecidomyidae, also known as Itonididae). These are two-winged insects more or less distantly related to fungus gnats. Their larvae live in the tissues of living plants where their presence (or already the act of laying the eggs) causes the plant to develop galls in which then the larva lives. Indeed, living inside self-induced plant galls is a part of a totally new ecological niche which did not exist before. The immediate ancestors of this family (or we can say, the ancestral species of that taxon) did not discover the presence of this possible ecological niche and then afterwards (evolutionarily) adapted to it, that is, occupied it. On the contrary, they created this niche for themselves.
Cases like this are abundant in the evolution of organisms and can only be made a bit more understandable by assuming the existence of a noëtic order (the Implicate Order) : The evolutionary changes taking place (in time) in some ancestral species, ancestral to the gall midges -- which is probably a species of the Jurrassic subfamily Pleciomiminae -- are really the projection ( = unfolding) into the Explicate Order of a product of some noëtic reaction between noëtic entities in the Implicate Order, entities involving the formal content of one of the species of the mentioned Jurrasic family and the formal content of a particular plant species. While the mentioned noëtic reaction and its product are in a state of enfoldment and thus space- and time-less entities, the corresponding evolutionary changes as they are seen (that is, deduced from observations) in the Explicate Order are the result of the unfolding along the space and time dimensions of the noëtic reaction product. These evolutionary changes, among other things, are : Decrease of body size (in adult and larval phases), complexification of the antennae of the adult midge, and development in the larva of external digestion of food. The family of gall midges as a whole has then extensively developed and diversified and still flourishes today.
So we have seen that a new ecological niche for a given animal species can come about not only as a result of change in an existing environment, for example, as a result of the advent of a new type of plant (say, angiosperms) or animal (say termites and their nests), but (such a new ecological niche) can also be created by the animal species itself of which it is then its proper niche. We cannot do other than to call such a case a biological invention done by the species, that is, by some collective of individuals. We cannot, however, describe it in this way as allegedly observed in the Explicate Order. Here only individuals act and react strictly according to their nature, that is, 'mechanically', be they organisms or the atoms and molecules which constitute them. Nothing is 'searched for', nothing is 'found', nothing is 'invented'. The only aspect of the workings in the Explicate Order that does remind us to these ('searching for', 'finding', 'inventing') is the natural selection of those patterns or things that are able to exist (at all, or in some given context). But this is more or less tautological : Those patterns or things that cannot exist do not exist. It becomes a bit less tautological when it is assumed -- as it is in biological natural selection -- that genetic mutations occur all the time and they take place more or less spontaneously. And only those mutations will survive which will be multiplied at the highest rate (escaping in this way from the Grim Reaper -- starvation, dehydration, etc. -- who is constantly depleting the ranks of the living) which is equivalent to the bearers of them to be more fit than those that harbor other genetic configurations. This is the well-known -- and well-accepted -- theory of natural selection from a set of random mutations as they take place in every population of living beings. But precisely because of this randomness, but also because there is no always-holding (universally valid) one-to-one corrrespondence between a gene (or group of genes) and a feature (or a group of features) as the latter is expressed and visible in the organism carrying the gene, the theory of natural selection of existing genetic mutants cannot pretend to present to us the very mechanism (and the only mechanism) of organic evolution. It is true that in many cases we, so to say, see it -- selection out of random mutations -- happening, but it cannot constitute the main mechanism of organic evolution. If we wished to fully explain the above described origin of gall midges by the model of natural selection of random mutations we would be forced to make a host of ad hoc assumptions about the nature and timing of the random mutations involved in this origin and their interaction with the other genes and their expression. The insects must become smaller in size, their antennae must become more sophisticated, their egg-laying habit and the structure of the ovipositor must be adapted, the anatomy of the anterior part ('head') of the larva and the physiology of certain of its mouth glands must be changed in such a way as to allow external digestion by the larva of tissue of living plants. It is clear that all this cannot be explained by the mutation-selection model or mechanism, although this mechanism can, and often will, play a role in organic evolution as we see it happening in the Explicate Order. Things have their ground in the Implicate Order in which they are interconnected by, what we might call, rational relationships.
There is one other feature that determines the general direction of evolutionary processes : the irreversibility of these processes. This is known as Dollo's Law. It says that when a sufficiently complex feature A, as it initially occurs in some given line of evolution of animals (or plants), has evolutionarily transformed into feature B, it will never happen that at some time this feature B is evolutionarily transformed back into precisely feature A. This law is, insofar as I know, generally accepted by paleontologists. Important for this law to apply is the relative complexity of such a feature and its derivative and the forbidden evolutionary return to exactly the same original feature. So when we find a group of recent organisms that possesses an original, that is, 'primitive', feature of sufficient complexity, we cannot maintain that this group has evolved from a corresponding fossil group of organisms (having existed far back in geological history) that possesses this feature in a strongly derived state, because then we would have to assume that in this respect the evolutionary process had reversed. Whether this law is really universally valid (in the way it is stated above) is not proven, but seems highly likely on the basis of an overview of all fossil material that has been accumulated by paleontologists during the last two centuries or so. This latter also applies to our own -- more general -- theory of the interpretation of the evolutionary process in terms of the Explicate and Implicate Orders. By consulting the data, as accumulated in the existing literature, on the fossil record of groups of organisms, we hope to find out whether our theory is completely compatible with all established facts of organic evolution. If not, the theory must be supplemented, amended, or rejected.
Having concluded the exposition of the general and fundamental aspects of our theory of the evolutionary-process-in-terms-of-the-Implicate / Explicate-Orders, it is now time to illustrate and further work out this theory in the light of expositions concerning the course of evolution as it has actually taken place, where this historical course is expressed (1) in the system of the relevant recent organisms ( this system, consisting of taxa and subtaxa, is the result of that course), (2) in their material structure, that is, their morphology, physiology, and individual development, and (3) in their behavior (instincts), and (4) in the age and morphology of relevant fossils.
Will, when necessary, be supplemented . . .
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the further study of Organic Evolution, Part II, Diptera, Tipulomorpha.
Back to Aristotelian metaphysics Part I
Back to Aristotelian metaphysics Part II
Back to Aristotelian metaphysics Part III
Back to Aristotelian metaphysics Part IIIa
Back to Aristotelian metaphysics Part IV
Back to Aristotelian metaphysics Part V