
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Most expressing example of the type :

Figure 1 : Asilus crabroniformis. Family Asilidae (robberflies), Order Diptera. 15-30 mm.
(After SEVERA, in Thieme's Insektengids, 1977).
Description of the type
Morphological features
Forewings significantly stronger developed than the hindwings. The latter are short, sometimes even completely absent. As a rule there is the formation of various coupling devices, unifying the hindwing with the forewing during wing-beat.
Mesothorax always significantly stronger developed than the metathorax. Muscular apparatus of metathorax strongly reduced as compared with that of mesothorax. The overall number of muscles of the thorax has decreased whereby in the most specialized forms steering muscles are explicitly present : various pleural muscles (pm) in 3-4 pairs. The chief mesothoracic muscles of indirect action [i.e. not directly attached to the wings themselves, but contracting and relaxing the thorax, resulting in the motion of the wings] are very powerfully developed in spite of their low number : there are only a few, but strong, such muscles.
In the great majority of forms, the wings have well expressed costalization, often reaching extreme limits. The known cases where the anterior part of the wing is not thus strenghened are, undoubtedly, only of a secondary nature. The hindwings [where present] possess a fan-like venation, often very reduced, almost absent. Their straight anterior margin lies closely against the posterior margin of the forewings and produces various hooks, folds, or outgrowths. Mechanically the hindwings form one whole with the forewings, playing the role of posterior lobe of them.
The shape of the body varies a lot, and in some most specialized forms indicates very progressed mechanical -- aerodynamic -- specializations. In such cases the body is shaped as an elongate-spindle, or is drop-like or spherically formed.
The absolute size of the insect varies strongly. The relative size of the wings in the representatives of the various subtypes differs. In little specialized forms the wings are, with respect to the body, very large, whereas in more specialized forms often reaching the smallest relative size among all flying insects.
Functional features
The representatives of dipterygia are characterized by the development of principally the mesothoracic wings, being the main, and sometimes even the sole, flight-organ. This feature determines all other structural traits. When the hindwings are developed, they, in the process of wing-beat, play the role of often very small appendages of the forewings. The beat of both pairs is strictly synchronous.
Speed of flight is best investigated precisely in this type. It is certain that within this very type the highest speeds are observed. Such speeds, up to 14-15 m/sec, are seen in the horse fly Tabanus bovinus and the hawk moths by Demoll, 1918. The flight of other members of the type is slower, bumble bees and honey bees make for 3-5 m/sec, various flies (Eristalis tenax [hoverfly], Calliphora sp. [blue-bottle], and Musca domestica [housefly] ) make from 2 to 2.7 m/sec.
Wing-beat frequency fluctuates on average from 110 up to 200 -- aculeate Hymenoptera and flies. Usually the wing-beat frequency reaches such values [in the present type], and only the daddy-long-legs Tipulidae have a lower frequency, from 44 up to 73 beats per second. The highest values of wing-beat frequency are found in mosquitos Culicidae, from 248 up to 307.
Maneuverability in flight is very perfect. Together with dragonflies, representatives of this type are best investigated.
Load per unit of surface area of wings reaches high values, fluctuating from 0.039 up to 0.103 gr/cm2 in the mosquito Culex, the flies Musca, Volucella, in the hawk moths Sphinx, Smerinthus, Hyloicus, and reaches high values in the Hymenoptera Bombus [bumble bee], Apis [honey bee], Vespa [common wasp], and in the robber fly Laphria -- 0.190-0.260 gr/cm2.
Persistence [and range] of flight is not especially investigated, but is undoubtedly significant.
The biological significance of flight is great for all representatives of the [present] type. The adult insects in this type spend most of their active life in the air. Undoubtedly, for certain hawk moths the range [distance] of flight is great -- there are observations about over-flight of the oleander hawk moth (Daphnis nerii) of 1200 km from the site of its emergence from the pupa. Securing food and reproduction is totally conditioned by a perfect flight in the most specialized flyers -- Hymenoptera and Diptera.
It is necessary to say that the features of flight -- wing-beat frequency, speed, maneuverability, and load -- are far from being investigated in all principal groups of this type. Thus, not investigated is the flight of sing cicadas, being undoubtedly perfect flyers comparable to the hawk moths, further, not investigated is the flight of various Diptera belonging to the subtype metadipterygia, aphids, the bugs Plataspididae, and a series of other groups. Knowledge of the flight features of these forms may profoundly change existing data after having significantly increased the extreme indexes of the qualities of flight.
Differentiation, connections, and representatives of the type
Two-wingedness has a special position among all the types. This fact consists, on the one hand, in the extreme "polyphyly" of dipterygia, having originated from the most diverse sources, and on the other hand, in the very nature of this type, this nature determining the working-out of those forms of flight which have the highest values of speed, maneuverability, etc. This legitimates to take dipterygia as the most perfect type of structure of the flight apparatus of an insect determining "the best" qualities of flight. The diversity of sources, and especially the widest development of two-wingedness results in great differentiation within this type, which can be divided up into a whole series of subtypes. See next Figure.
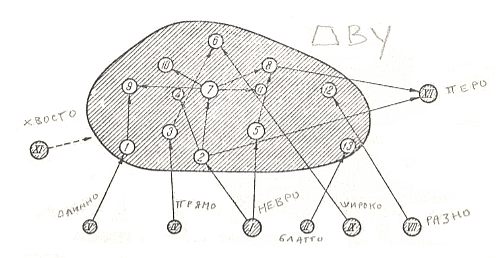
Figure 2 :
Interrelationships with other types, and structure of the type of two-wingedness (dipterygia).
Arabic numbers - subtypes of dipterygia.
From left to right other types :
XI - uropterygia, V - dolichopterygia, IV - orthopterygia, I - neuropterygia, II - blattopterygia, IX - platypterygia, VII - heteropterygia, XII - ptilopterygia.
(After ROHDENDORF, 1949, with additions)
1. Archidipterygia (ancient two-wingedness)
This subtype is the earliest of all. Its source were forms of long-wingedness (dolichopterygia) of certain palaeopterous insects. Characteristic of the subtype is the low degree of reduction of the wing-venation, moderate development of costalization, as a rule the wings preserve a rich little-specialized venation. Processes of specialization of the venation, where they have taken place, consist either in the cancelling of certain longitudinal veins or sections of them. No displacement or thickening of veins is observed. Hindwings in various sizes, significantly reduced. Sometimes the processes of reduction go very far and the hindwings then totally disappear.
The body at its rear end carries long threads (cerci and sometimes a paracercus) which have a determined, but not further investigated, aerodynamic significance. These appendages of the abdomen are inherited from ancestral forms of representatives of dolichopterygia. As in the latter, archidipterygia carries an ancient feature, namely the absence of devices for folding the wings back onto the body : the wings are always held sideways (spread-out) or held upwards.
The flight doesn't show up high flight-parameters.
The structure of the thorax is weak -- pleural sclerites weakly connected to one another, not constituting a firm thoracic skeleton necessary for the functioning of strong thoracic muscles. Size of the wings relative to the body big, load small.
To this subtype do belong recent and mesozoic mayflies, Ephemeroptera, from which in certain groups, like for instance Heptageniidae, Ephemeridae, Baetiscidae, and many others, we see the most primitive expression of the type. Other families of the Order show far-reaching processes of reduction of the hindwings -- such Lentophlebiidae, Baëtididae. Another evolutionary direction is realized in the families Palingeniidae, Behninggiidae, Oligoneuriidae. In these groups the hindwings undergo insignificant reduction, but with nevertheless the cancelling of many longitudinal and almost all cross-veins, and, as a result of this process, a marked rarification of the whole system of the wing-venation.
On the basis of this subtype another one originated, fan-winged dipterygia or strepsidipterygia. Ancient two-wingedness is the geologically earliest appearance of dipterygia. This is documented by the Permian Archodonata, having a structure similar to recent youngest groups of mayflies. They had totally reduced hindwings. See next Figures.
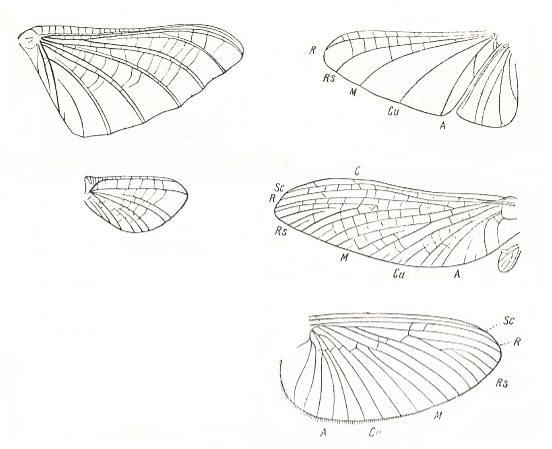
Figure 3 :
Left : Wings of Behningia ulmeri Lest. Family Behningiidae, Order Ephemeroptera.
(After TSJERNOV, in ROHDENDORF, 1949.)
Right :
Top : Wings of Oligoneuriella rhenana Imh. Family Oligoneuriidae, Order Ephemeroptera.
Center : Wings of Teloganodes tristis Eat. Family Leptophlebiidae, Order Ephemeroptera.
Bottom : Wings of Ordella horaria L. Family Brachycercidae, Order Ephemeroptera.
(All three, after HANDLIRSCH, in ROHDENDORF, 1949)
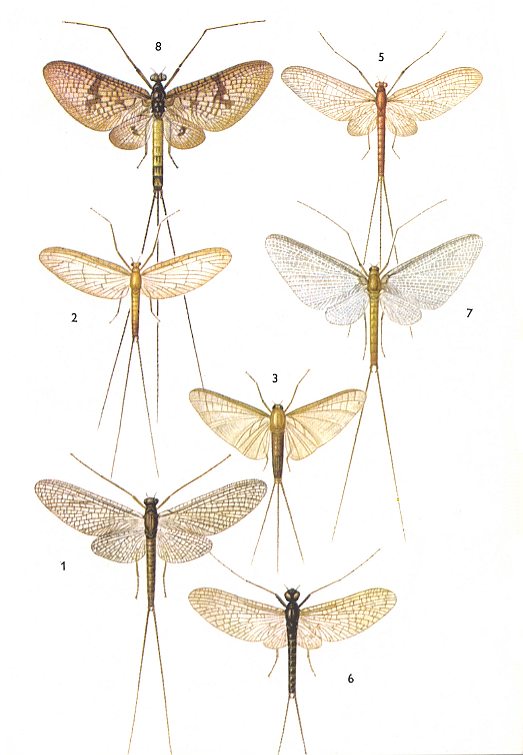
Figure 4 : Ephemeroptera (mayflies).
1. Siphlonurus lacustris. 11-14 mm, + cerci 16-22 mm. Family Siphlonuridae.
2. Cloëon dipterum. 6-8 mm, + cerci 8-17 mm. Family Baetidae.
3. Oligoneuriella rhenana. [?]3-13 mm + cerci 9-16 mm. Family Oligoneuridae.
5. Ephemerella ignita. 6-9 mm, + cerci 12-18 mm. Family Ephemerellidae.
6. Habrophlebia lauta. 5-7 mm, + cerci 6-9 mm. Family Leptophlebiidae.
7. Polymitarcis virgo. 10-18 mm, + cerci 15-35 mm. Family Polymitarcidae.
8. Ephemera danica. 15-24 mm, + cerci [?]14-40 mm. Family Ephemeridae.
(After SEVERA, in Thieme's Insektengids, 1977)
2. Neurodipterygia (primitive two-wingedness)
This subtype originated from the basis of various forms of neuropterygia one of the most ancient types of flight devices. As a rule characteristic of this subtype is an advanced reduction of the venation, in the forewing consisting in the shortening and disappearance of many branches of the main trunks of the veins, [disappearance] of the majority of the cross-veins, and in the presence of a certain degree of costalization -- strenghening of the veins at the anterior margin and the latter's straightening. The hindwings undergo these changes in an even greater degree. In a number of cases they have a quite impoverished venation. The size of the hindwings is relatively variable. Sometimes they are even almost totally reduced. The size of the wings relative to the body is large.
Skeleton of the thorax little strenghened. The flight, apparently does not show high parameters. At rest the wings are always folded back onto the body.
To this subtype do belong diverse insects : Neuroptera of the family Coniopterygidae, extinct mesozoic Paratrichoptera, Homoptera of the superfamily Psylloidea (jumping plant lice), book-lice (Psocoptera) of the families Myopsocidae and Psocidae.
Maybe also belonging to this subtype of dipterygia are certain peculiar Neuroptera from the families Hemerobiidae (Psectra) and caddis flies (Trichoptera) from the tribe Bajcalinini (Thamastes).
A series of features of these insects (insignificance of mechanical specialization of the forewings, their shortening) calls for caution in evaluating these, probable, regressive changes. See next Figures.
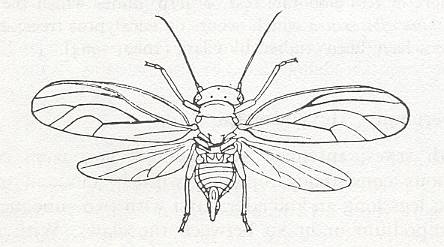

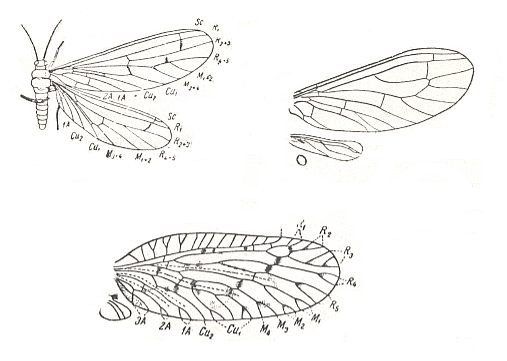
Figure 7 :
Top left : Spiloconis maculata End. Family Coniopterygidae, Order Neuroptera. Australia. Length of forewing 4.5 mm. (After TILLYARD, in ROHDENDORF, 1949)
Top right : Wings of Conwentzia pineticola End. Family Coniopterygidae, Order Neuroptera. Europe. (After ENDERLEIN, in ROHDENDORF, 1949)
Bottom : Wings of Psectra diptera L. Family Hemerobiidae, Order Neuroptera. Europe. (After KIMMINS in ROHDENDORF, 1949)
3. Homodipterygia (dipterygia of minute cicadas)
This subtype is a derivative from the orthopterygia (straight-wingedness) of the Homoptera, and characterized, on the one hand, by the preservation of traits of the mentioned type, but, on the other hand, by the presence of features of its own (i.e. special features). The venation of the forewings is extraordinary strong. Veins small in number but thick, forming a characteristic closed net, leaving free membrane along the outer wing-margin. Costalization well expressed. Hindwings relatively large, with rarefied but strong venation. Skeleton of the thorax firm, musculature and flight itself almost not investigated. Size of wings with respect to body variable, usually not large, more rarely the wings are long (Derbidae).
To this subtype do belong certain cicadas -- Machaerotidae and many Membracidae. Perhaps also be included should be the peculiar group of cicadas, the Derbidae (genus Zoraida and relatives), distinguishing themselves by rather long costalized forewings. See Figures.
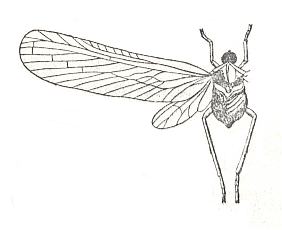
Figure 8 : Zoraida egregia Mel. Family Derbidae, Order Homoptera. Wingsspan 16 mm. Sri Lanka.
(After MELIGAR, in ROHDENDORF, 1949)
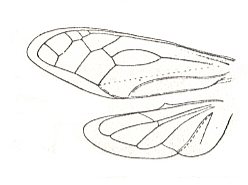
Figure 9 : Wings of Machaerota spangbergi Sign. Family Machaerotidae, Order Homoptera. Southern Asia.
(After HANDLIRSCH, in ROHDENDORF, 1949)
4. Aphidodipterygia (two-wingedness of aphids)
The characteristic insects of the temporate Zone -- aphids, Aphidoidea, possess highly specialized wings. The whole construction of the body as a flight device forces us to erect for them a special subtype. The origin of aphidodipterygia is evident : the source is to be found in the primitive dipterygia of the ancestors of aphids. An idea of such early forms might be shown by the group Psylloidea (jumping plant lice) and, perhaps, Psocoptera (book lice). The dipterygia of the aphids is characterized by a high stage of costalization of the forewing having thickened veins at the anterior margin. The latter is straight or a little convex. Also characteristic is the presence of a well developed large pterostigma, a strongly reduced venation of the remaning part of the wing, and, finally, an elongate triangular shape of the wing. Hindwings not large, firmly connected with the forewings. They have a fairly constant size, seldom reduced to especially minute size. The size of the wings with respect to the body is large. In resting position they are folded backwards.
Thorax moderately sclerotized. Very small insects. Their flight is active, but little studied. To this subtype, which isn't similar to any other, only aphids do belong. See Figures.
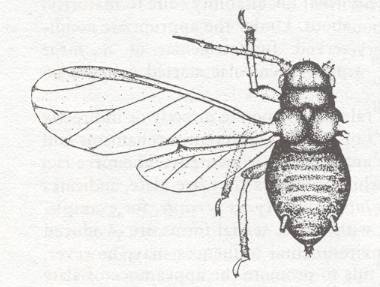

Figure 11 :
Left image : Wings of Phylloxera coccinea Heyd. Superfamily Aphidoidea, Order Homoptera. Europe.
ha - coupling bristles. pt - ptererostigma.
Right image : Brasilaphis bondari Mord. Superfamily Aphidoidea, Order Homoptera. South America.
siph - secretional tubes on abdomen.
(After MORDVILKO, in ROHDENDORF, 1949)
5. Hymenodipterygia (two-wingedness in Hymenoptera
This subtype is very peculiar and can only, through its most primitive forms, with difficulty be connected to some other more primitive form. The next Figure depicts a typical representative of the present subtype.
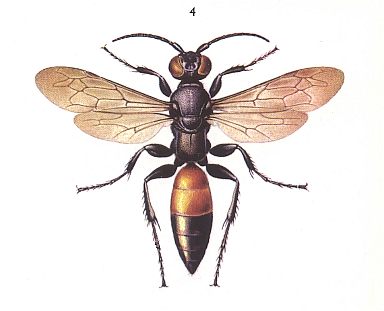
Apparently, we must assume the formation of hymenodipterygia from material having some form of neuropterygia in which traits of dipterygia were developed with very early formation of a strongly sclerotized thorax together with development of powerful musculature. Characteristic to hymenodipterygia is a well expressed costalization of the forewing consisting in the strenghtening of the anterior margin, often the development of a pterostigma, transformation of the post-costal venation into a firm net, consisting of a system of a small number of firm veins, leaving a weakly strenghened posterior margin. Hindwing relatively large (about 2/5-1/3 of the length of the forewing), of constant size, firmly coupled to the forewing, in flight forming one mechanical whole with the latter.
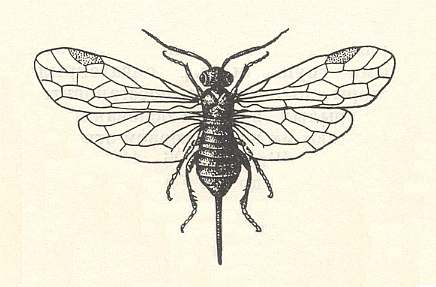
The other end of the series [of certain forms of the present subtype] contains, evidently, the aculeate Hymenoptera, for instance Vespidae (wasps). Another kind of change is expressed in the groups Ichneumonoidea and Braconoidea and characterized by the large size of the wings with relatively weak venation, but nevertheless [advanced in the sense of] fairly strongly reduced. Together with this, characteristic for this change is a progressed "lightening" of the body, having a small head, thorax, slender abdomen, and long appendages of the body -- antennae, legs, and often a long ovipositor (certain Pimplidae and Braconidae) being comparable with the cerci of other insects as to its aerodynamic effect in flight.

For a typical (solitary) (true) bee, see next Figure.

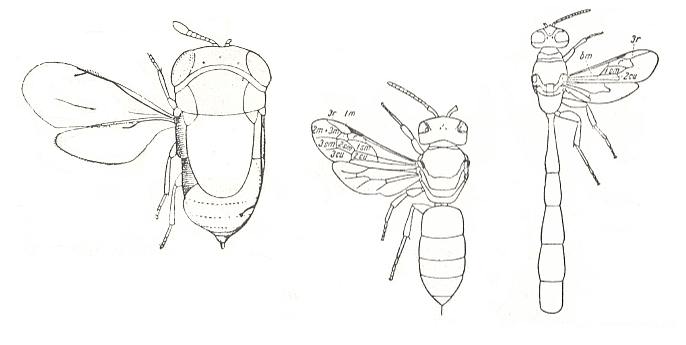
Figure 16 :
Left image : Scutellista cyanea Mot. Family Pteromalidae, Order Hymenoptera. Length of body 1.3 mm.
Middle image : Hyaleus capitosus Sm. Superfamily Apoidea (bees), Order Hymenoptera. Length of body 10 mm.
Right image : Trypoxylon connexum. Turn. Superfamily Sphecoidea, Order Hymenoptera. Australia. Length of body 13 mm.
(All Figures after TILLYARD, in ROHDENDORF, 1949)
6. Megadipterygia (dipterygia of large insects)
To this subtype [not to be confused with metadipterygia, which is being dealt with further below] do belong different insects, themselves belonging to two Orders phylogenetically far apart from one another : Lepidoptera (butterflies and moths) and Homoptera (cicadas, etc). These, apparently, totally disparate insects are unified through a number of features. Such are the large or medium size of the body and the strongly elongate wings, possessing moderate costalization of the venation together with the reduction of a large part of the cross-veins. Hindwings relatively large, usually about 3/5 of the length of the forewings, more seldom shorter. Wing-venation fairly rich. In resting position the insect folds the wings back onto or along the abdomen. Thorax, as a rule, very massive and strongly built, possessing a strong muscular apparatus. The shape of the body is characteristic : The front part is more or less thickened, little pointed. Towards the end the body is definitely thinning out, pointed. Legs short. All these features are definite features of aerodynamic specialization of the body. Size of the wing-surface as compared to the weight of the body small. Flight is investigated in one group only, namely in hawk moths, Sphingidae (Oder Lepidoptera), distinguishing themselves by the highest indexes of range [of flight] (many hundreds of kilometers) of all insects.
Megadipterygia originated, on the one hand, from forms of broad-wingedness (platypterygia) of certain Lepidoptera (in particular of the subtype diplatypterygia), and, on the other, is based on the homodipterygia of cicadas. The known forms of megadipterygia point to the diversity of the subtype.
Together with the typical representatives [of the subtype], characterized by large size, massive aerodynamic and specialized body, well developed hindwings, such as the Sphingidae (Lepidoptera), many cicadas (Homoptera), and the fossil Palaeontinidae, there exist smaller Lepidoptera -- Cuculiidae, Psychidae (Oiketicus), and others characterized by a more elongate body. Another change within the subtype is illustrated also in Lepidoptera : certain Zygaenidae and Syntomididae (Trichura). In these insects the hindwings show a more advanced stage of reduction, together with relatively insignificant elongation of the forewings, and weak development of the thorax. It may even be that the forms of the latter example should be placed in a special group after having studied things in more details.
Biological significance of flight is very diverse. On the one hand, for particular groups of Lepidoptera, for example the Sphingidae, flight realizes feeding and distribution (the insects feed, in flight, on nectar from flowers, and fly long distances), other groups, also Lepidoptera, are, apparently aphagous [they do not feed] (Psychidae, Oiketicus), and flight for them has only significance in reproduction. The significance of flight in sing-cicadas is not known to me [Rohdendorf] (is it for protection of for distribution?).
See next Figures.

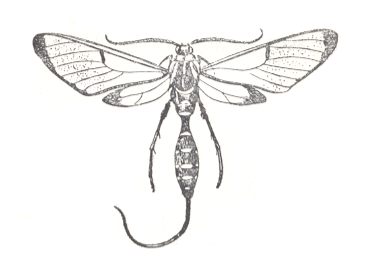
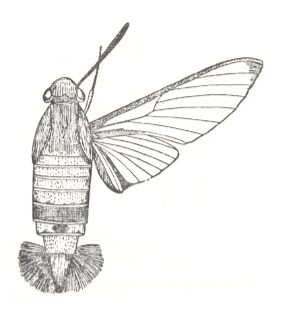

7. Eudipterygia (true two-wingedness)
We begin with some Figures illustrating the subtype (in the Order Diptera) :
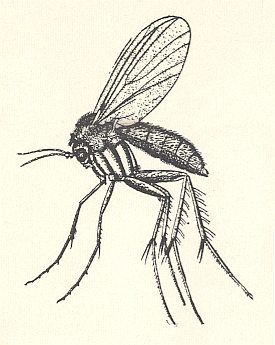

Figure 22 : A male adult of the New Zealand glow-worm, Arachnocampa luminosa (Fungivoridea-Ceroplatidae). This is a rare photograph of an elusive fly, but its appearance is typical of land-midges all over the world.
(After OLDROYD, 1964)
See also Figure 1, above.
And further :

Figure 23 : Some representatives of the Lifting Lightly-veined (muscoid) functional wing-type [in ROHDENDORF's classification (1951) of the wings of Diptera].
1 - Chlorops pumilionis, Yellow stem-fly, 2.5-3.5 mm. Family Chloropidae (Acalyptratae).
2- Oscinella frit, Frit-fly, 1.5-2 mm. Family Chloropidae (Acalyptratae).
3- Delia brassicae, Cabbage-fly, 5.5-7.5 mm. Family Anthomyiidae, flower-flies (Calyptratae).
6- Musca domestica, House-fly, 7-9 mm. Family Muscidae (Calyptratae).
(After SEVERA, in Thieme's insektengids voor West- en Midden-Europa, 1977)
To this subtype do belong the majority of the Order Diptera and certain other insects. They are, morphologically, the most specialized forms of dipterygia. Most characteristic is the complete reduction of the hindwings with strong development of the mesothorax, its skeleton and musculature, together with various mechanical, often very advanced adaptations of the wing-venation, shape of the body and other features. The complete concentration of the flight-function in the forewings, being appendages of the metathorax, caused the reduction of the other divisions of the thorax, being transformed into insignificant appendages of the mesothoracic skeleton, carrying merely the corresponding pairs of legs with their musculature. In the structure of the whole body first of all as to its shape, one observes various specializations of which the shortening of the body is the clearest expression directly related to the enlargement of the thorax. But the most characteristic trait of this subtype are the structural features of the wings which acquire a special form. Anterior margin of wing straight, apex quite distinct, termen very large (See Part I Figure 1) and reaches far along the posterior wing-margin. The latter in its basal half gives off various membranous appendages, lobes and blades, often rather large.
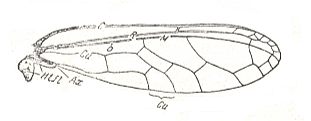
Figure 24 : Wing of Muiria sp. Family Derbidae, Order Homoptera. Htfl -- rudiment of hindwing.
(After HAUPT, in ROHDENDORF, 1949)
characterized by elongate costalized wings with a remarkably structured wingbase.
8. Metadipterygia (costalized two-wingedness)
This subtype is characterized by the extreme stage of specialization of the wing-venation, consisting of a small number of very strong veins at the anterior margin of the wing. The remaining venation is weakly expressed, reduced, sometimes even totally absent. Hindwings without venation, more seldom with a vein carrying the coupling apparatus. Sometimes the hindwings are absent. [So here, in this subtype, the shift of concentration of the flight-function still further to the front is driven to its extreme.]
This subtype is the derivative of two sources : eudipterygia and hymenodipterygia. The representatives of these subtypes independently acquired complex features while developing and strenghtening costalization of the wing. The formation of such highly costalized wings, testifying to a high wing-beat frequency, goes together with an enlargement of the thorax -- container of powerful muscles.
To this subtype do belong various Diptera, namely midges and gnats of the families Tendipedidae (especially of the group Corynoneurinae) and Heleidae, flies of the family Phoridae, Stratiomyidae, and certain Sarcophagidae (for instance Nyctella), and certain Hymenoptera, namely various Chalcidoidea (small parasitic wasps).
In this subtype, as well as in the previous one, also further subdivisions can be seen, characterized by structural features of the basiala, and the shape of the wings and body. The subtype is the limiting phase of transformation of two-wingedness, and in essence its further evolution will merely boil down to regressive processes, namely to the formation of special forms of ptilopterygia (feather-wingedness). See next Figures, illustrating the present subtype.
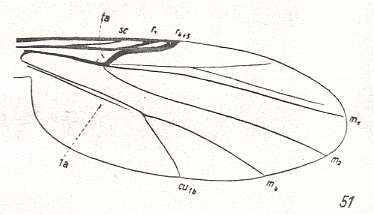
Figure 25 : Wing-venation of Forcipomyia ciliata WINN. Order Diptera, Family Ceratopogonidae ( = Heleidae), Superfamily Chironomidea. (After HENNIG, 1954)
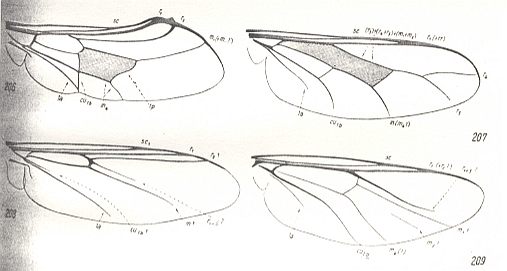
Figure 26 : Some more representatives of the Lifting Costalized (stratiomyoid) functional type of wings [in ROHDENDORF's classification (1951) of the wings of Diptera].
Cyrtidae [= Acroceridae = Oncodidae], Order Diptera.
206 : Pterodontia misella O.S. 207 : Acrocera convexa Cole. 208 : Philopota aenea Meig. 209 : Oncodes zonatus Erichs. (After HENNIG, 1954)
Figure 27 : Wings of representatives of the Lifting Ultra-costalized (phoroid) type [in ROHDENDORF's classification (1951) of the wings of Diptera].
Top image : Aphiochaeta sp., Phoridae. Length 3.5 mm. (After Rohdendorf, 1951).
Bottom image : Palaeophora ancestrix Rohd., Palaeophoridae. Jurassic of Karatau (Southern Kazachstan). Length a little less than 2 mm. (After Rohdendorf, 1938) [ In 1938, in Trudy Paleontologicheskovo Instituta, Tom VII, Issue 3, Rohdendorf had described this fossil -- nr 2452/329 -- as Archiphora ancestrix . The name had to be changed because of the priority of the name Archiphora (see next Figure) ].
(From ROHDENDORF, 1951)
The next Figure depicts wings of Phoridae, belonging to the present subtype. Their derivation from clythioid wings is shown.
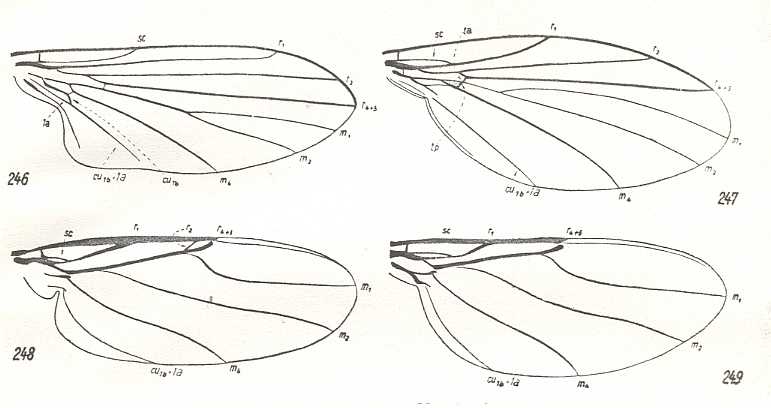
Figure 28 : Venation of Phoridea, compared with that of Clythiidae.
246 - Opetia nigra Meig., Clythiidae. (After Schmitz, 1929). Clythioid subtype of muscoid type [in ROHDENDORF's classification (1951) of the wings of Diptera].
247 - Sciadocera (Archiphora) patagonica Schmitz, Sciadoceridae. (After Schmitz, 1929). Sciadoceratoid subtype of empidoid type [in ROHDENDORF's classification (1951) of the wings of Diptera].
248 - Aneurina thoracica Meig., Phoridae. (After Hennig, 1954). Phoroid type [in ROHDENDORF's classification (1951) of the wings of Diptera].
249 - Phora stictica Meig., Phoridae. (After Hennig, 1954). Phoroid type.
246 ==> 247 ==> 248 ==> 249 forms a neat derivational line.
(From HENNIG, 1954)
Further, a member of the Chalcidoidea (Order Hymenoptera) also belonging to the present subtype :
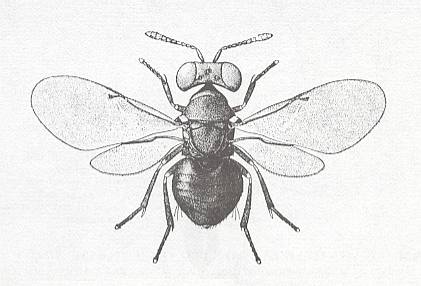
9. Strepsidipterygia (fan-winged two-wingedness)
This subtype is characterized by the broadening of the wing-blade turning the wing into a broad fan-like lobe.
The subtype originated from two not much similar to one another sources -- the archidipterygia of mayflies (Order Ephemeroptera) and the eudipterygia of certain Nematocera (midges, gnats, mosquitos) (Order Diptera). Difference of origin entails a fairly great difference between the two groups composing the subtype -- the mayflies Brachycercidae and the blackflies Simuliidae. The broad wings of the Brachycercidae are little costalized and have a radially structured venation. Simuliidae, on the other hand, possess strongly costalized wings. The body of the representatives of strepsidipterygia is short with a large thorax. The mayflies, moreover, possess long cerci at the end of the body, with their presence, undoubtedly, producing special aerodynamic conditions during flight. This subtype, as does the previous one, shows it to be a peculiar extreme transformation of eudipterygia, namely into the direction of increase of the surface area of the wings, their posterior half. See next Figures.
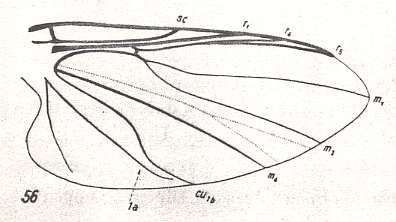
Figure 30 : Wing-venation of Prosimulium rufipes MEIG. Order Diptera, Family Simuliidae ( = Melusinidae), Superfamily Chironomidea. Basal cell not drawn. (After HENNIG, 1954)
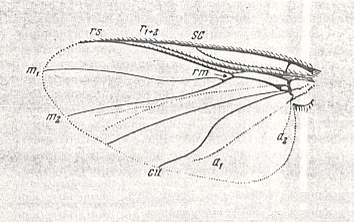
Figure 31 : Wing of Simulium sericatum MEIG. Order Diptera, Family Simuliidae ( = Melusinidae), Superfamily Chironomidea.
(After HENDEL, from ROHDENDORF, 1951)
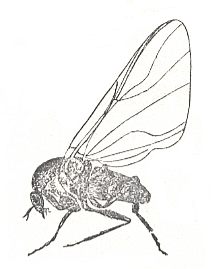
Figure 32 : Simulium hirtipes Fries. Family Simuliidae, Order Diptera. Europe.
(After LINDNER, in ROHDENDORF, 1949)
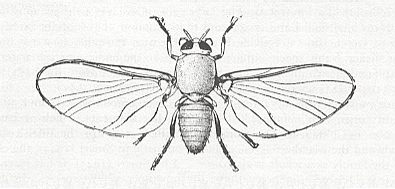
Figure 33 : Simulium venustum, female. Family Simuliidae, Order Diptera. Wingspan about 6.7 mm. N. America.
(Trustees of the British Museum, in RICHARDS and DAVIES, 1977)
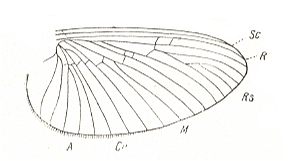
Figure 34 : Wing of Ordella horaria L. Family Brachycercidae, Order Ephemeroptera.
(After HANDLIRSCH, in ROHDENDORF, 1949)
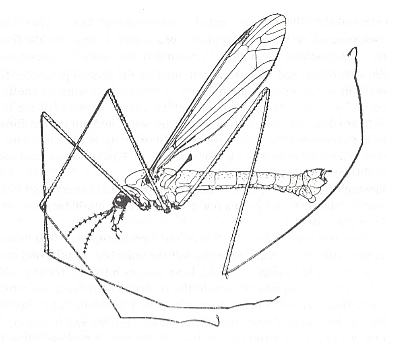
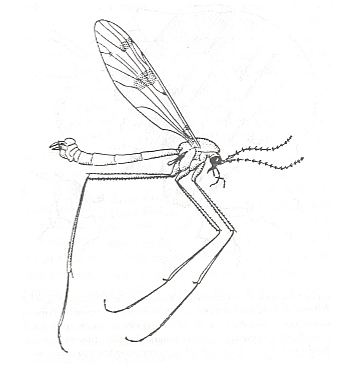
10. Kopedipterygia (paddle-winged two-wingedness)
This subtype of dipterygia, namely kopedipterygia, only includes the characteristic craneflies, Tipulidae, and partly Limoniidae. These insects have worked out strongly elongate moderately costalized wings with a comparably little reduced venation, and, what is especially characteristic, with a strongly narrowed base, basiala, which has obtained a peculiar venation. In these Diptera the basiala is formed by strong closely to one another running veins, which [sets of closely running veins], after having in turn moved to one another, form a narrow handle of the wing. A section through the basiala shows the peculiar tube-like structure of this organ consisting of concave and convex veins. Such a structure of the basiala guarantees its strength and elasticity to be greater. The venation of the wing-blade is characteristic [rendering craneflies directly as such recognizable] also by the shift towards the distal end of the wing of cross-veins and of the forks of the majority of longitudinal veins, determining the special nature of the venation as a whole and of the aerodynamics of the wing.
The body doesn't show any adaptation to fast flight, it doesn't have a streamlined shape. The appendages of the body -- legs and antennae -- long and narrow, abdomen long, heavy in females, and swollen at the end in males.
Kopedipterygia is a very ancient form of two-wingedness already present in Jurassic times. In this subtype we see a certain similarity with the kopepterygia of damselflies (Odonata, Zygoptera) and that of the scorpionflies Bittacidae (Mecoptera). Such analogous specialization, having originated wholly independently from different sources (in the one case [Zygoptera and Bittacidae] from respectively long-wingedness (dolichopterygia) and neuropterygia, in the other case (Tipulidae) from two-wingedness (dipterygia)), undoubtedly was an effect of similar environments determining the working-out of a certain flight-regime. Today it is not yet possible to precisely characterize the features of "paddle-winged" flight. Apparently for such a flight-regime it is very useful to obtain the greatest possible length of the wing with least load onto its basal part. See Figures.
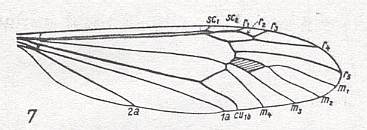
Also {not specifically mentioned by Rohdendorf) having paddle-shaped wings are midges of the Family Ptychopteridae (Order Diptera, Nematocera) :
The wings of these forms have a venation different from that of Tipulidae :
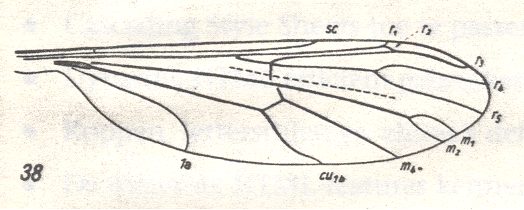
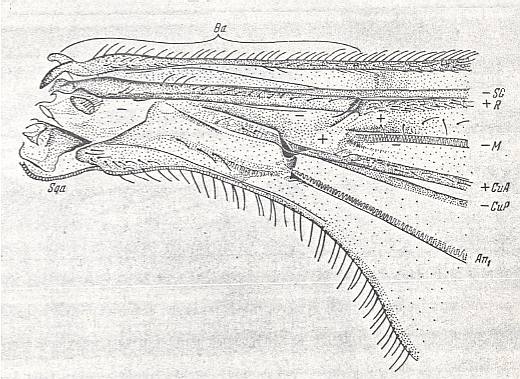
Figure 39 : Liriope contaminata MEIG. ( Liriopeidae [ = Ptychopteridae] ).
Base of right wing of male. Topview.
Specimen Nr.3684. Length of wing 7.50 mm. Length of part starting from the level of the base of the costal vein to that of the humeral cross-vein [the cross-vein between Costa and Subcosta], supposedly delineating the basiala, 1.35 mm.
For the abbreviations, see Figure 1a in Fifth Part of Website, Organic evolution part II . The brace (in upper part of figure) with the sign Ba indicates the extent of the supposed basiala.
(After ROHDENDORF, 1951)
The reader will find more about this type of wings in the documents on evolution in Fifth Part of Website.
11. Platydipterygia (not-costalized two-wingedness)
This subtype is characterized by the development of very broad wings together with the almost complete absence of features of costalization. The anterior as well as the posterior wing margin almost uniformly strengthened. Wing-venation rich, evenly distributed over the whole wing-blade. The basiala broad, carrying many interruptions and various bugings of the veins. The wings, as well as the whole body, covered with long hairs and scales.
The subtype is best expressed by the representatives of a peculiar family of Diptera, the Psychodidae (moth-flies). With a certain degree of qualification, apparently also do belong to the present subtype the flies of the family Musidoridae in which also we see the almost total absence of costalization of the venation. But the wing of Musidora is narrower [than in Psychodidae] and almost naked without hairs or scales which sharply distinguish the wing from that of the Psychodidae.
The flight-features are not investigated. The origin of the subtype is more or less clear : It, apparently, is a derivative of such forms of eudipterygia that we saw in Culicidae (mosquitoes) and relatives. See next Figures.
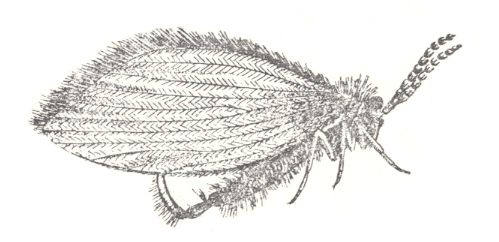
Figure 40 : Psychoda phalaenoides L. Family Psychodidae, Order Diptera. Europe.
(After LINDNER, in ROHDENDORF, 1949)
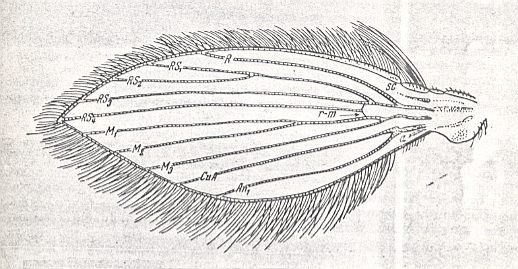
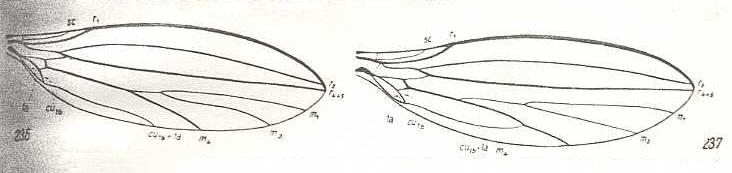
Figure 42 : Wing-venation of representatives of the lifting not-costalized (musidoroid) type [as in ROHDENDORF's classification (1951) of functional wing-types in Diptera].
Left image : Musidora strobli de Meij., Musidoridae. Male.
Right image : Musidora strobli de Meij., Musidoridae. Female.
(After HENNIG, 1954)
12. Heterodipterygia (folding-shield two-wingedness)
This subtype is represented by only a single group, namely of the most peculiar bugs (Order Heteroptera) of the Family Plataspididae possessing large tranversely folded forewings with a significantly strengthened anterior margin and a pectinate venation in the distal folded part. Hindwings as to size less than half of the forewings with a fairly simple radial venation. The body doesn't show aerodynamic adaptations.
This subtype is a direct product of certain forms of heteropterygia, and apparently illustrates a higher stage of development of flight device in the Order Heteroptera. The ability of transverse folding of the forewings is unique in the whole class of insects. Flight of these insects not studied. See next Figure.
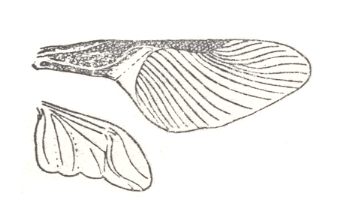
Figure 43 : Wings of Brachyplatys sp. Family Plataspididae, Order Heteroptera. Tropics.
(After HANDLIRSCH, in ROHDENDORF, 1949)
13. Blattodipterygia (two-wingedness of cockroaches)
To this subtype do belong little known insects, the cockroaches (Order Dictyoptera) of the genus Hypercompsa, in which clear traits of dipterygia are developed. Forewings longer than the hindwings, showing well expressed features of costalization. Hindwings fairly broad, notably costalized with a broad ano-jugal area and radial venation. Flight not investigated. The origin of the subtype is clear -- it is a progressive derivative of blattopterygia one of the most ancient types of flight-devices in insects. See next Figure.
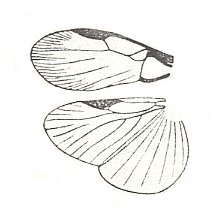
Figure 44 : Wings of Hypercompsa fieberi Brun. Family Euthyrrhaphididae, Order Dictyoptera. South America.
Length of wings 6-7 mm.
(After HANDLIRSCH, in ROHDENDORF, 1949)
It is possible to add another subtype of two-wingedness represented by the flight-apparatus of the peculiar and little known tropical Zoraptera. The general look of these insects partly reminds one of Hymenoptera, and partly of Psocoptera (book-lice). The still insufficient study of the Zoraptera doesn't make possible to clarify flight-features. See next Figure.
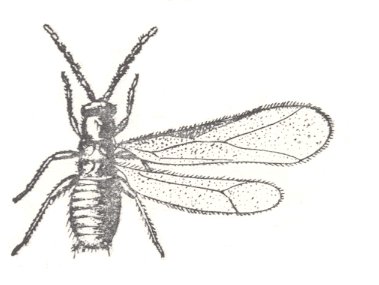
Figure 45 : Zorotypus hubbardi Coud., Order Zoraptera. North America.
Length of body 1.5 mm.
(After COUDEL, in ROHDENDORF, 1949)
As we've seen, two-wingedness (dipterygia) underwent the most diverse differentiation as compared with any other type (See Figure 2 above). The diversity of sources, the very nature of two-wingedness, apparently, being the most progressive and perfect type of flight-devices in insects, all are the natural and evident clarification of this fact [of being the most progressive type].
End of our exposition of ROHDENDORF's classification (1949) of the flight-devices in insects.
We have now concluded our exposition of ROHDENDORF's (1949) classification of the flight devices (mainly wings) in insects from a functional perspective. ( In Fifth Part of Website we have treated Rohdendorf's subsequent (1951) classification of functional wing-types in the Order Diptera).
All authors writing on the subject of insect wings (and all other structures for that matter) try to establish "homologies" of the various morphological parts, here especially the parts of the venation (for example to recognize the "radial sector" in the wings of diverse insects). The concept of homology, originally (and that was before the establishment of evolutionary theory) conceived as formal morphological relationship between corresponding material features in different organisms, "rational morphology", is now (after acceptance of evolutionary theory) theoretically based on common descent of these features as a result of common descent of animal or plant species. To repeat, today homology of material structures in organisms is not based anymore upon a purely formal relationship between such structures, but upon common descent, i.e. upon history, here meaning material continuity. And common descent is part of evolution as it is taken to be today. And evolution itself is, according to current view, driven by "natural selection" (of genetic mutants, provided for by random (not functional!) mutation of genetic material). So natural selection is the ultimate explanation of the phenomenon of homology of material structures in organisms.
So while "analogy" (in contradistinction to homology) is based exclusively on functional features, such as the analogy between the bird's wing and the insect's wing, homology is based on historical material continuity (of actual structure and its code in DNA) of structures. But within this continuity the appearance of corresponding structures in organisms is nevertheless functionally 'motivated', because evolution is taken to be driven by natural selection of the functionally best alternatives (given by random and spontaneous genetic mutation) (while genetic mutation is not functional, natural selection is). Such functionally best alternatives have the greatest chance to get established because they best support reproduction, and thus the continuance of the species.
And also in our theory of the Explicate and Implicate Orders (i.e. our "noëtic" theory of 'evolution') the development, in the Implicate Order, of structures is driven by functionality in order to produce a "strategy" (expressed by the overall morphology, physiology, and behavior of [individuals of] a [given] species of organism), allowing the possessor of it to materially exist and persist in the Explicate Order. But, in this, our's, view, the development, 'construction', of all the parts of the strategy takes place, not in the Explicate Order, but in the Implicate Order. The space-time nature of explicate reality cannot produce the functional, this can only be accomplished in "noëtic space", the Implicate Order. And there this development is timeless, despite the fact that it is governed by current ecological situations and settings in the Explicate Order. But these situations merely trigger the "projection" of particular strategies from the Implicate Order into the Explicate Order appearing there as organismic species. And thus, because there is no material development and descent of organismic species in and by the Explicate Order, the only historical aspect of transformations of features is the successive order (in time) of projection of such features into the Explicate Order, and thus the successive appearance in this Order. And because the Implicate Order, where strategies are 'constructed' (derived), is completely timeless (and spaceless), there is also no historical aspect of constructions in the Implicate Order. And, moreover, we have found out that the 'development' of strategies, organismic species, in the Implicate Order, is polyphyletic, not monophyletic. So also here there is no historical continuity between organismic species. So in our theory the essence of homology, what it really is, is once again based on "rational morphology" only, but nevertheless still functionally driven. It is rational morphology again, because there is no historical continuity between 'homologous' structures. And because the development takes place in noëtic space, there are, in the development of structures, no spatial or temporal limits, there is no spatial or temporal separation of developing structures and strategies. So in development of wing-venation (and in any other morphological set for that matter) there can be a 'cross-traffic' between the various derivational lines (sequences), and thus between insect taxa, and so even between the various insect Orders, These Orders are, however, established by taxonomists on the assumption of their independent development, evolution. But according to our theory such 'taxa' are not completely independent units, they are merely typologically more or less homogeneous, and have not, each for themselves, monophyletically developed. So in our theory there is much more play for exchange of characters between even widely separated taxa, and that is possible in noëtic space, not in true space. And, indeed, also Rohdendorf's functional classification of flight devices in insects is, despite the fact that each type is maximally represented in a particular insect Order (or smaller taxon), a cross (insect-)Order classification. We see this, for example, in feather-wingedness (ptilopterygia), which type includes members of the insect Orders Thysanoptera (thrips), Coleoptera (beetles), Lepidoptera (butterflies and moths), Trichoptera (caddis flies), and others, while being most widely and typically distributed in the Order Thysanoptera.
Although the 'process' in the Implicate Order of the 'generation' of an organic strategy IS a derivational process, it is not so that one strategy is derived from another. Certain strategies are at most "like one another", formally similar. There is no "evolution" in the sense of a series of successive strategies in which (series) they evolve one from the other. Derivation in the Implicate Order takes place in separate more or less independent "derivational lines" (but always directed by the current ecological situation and setting in the Explicate Order, as a result of the phenomenon of "injection" (alongside projection)). Every such a derivational line ends up in a particular strategy (or part of it), built up, so to say from scratch, but may incorporate morphological or otherwise 'modules' from other such lines. The same is the case in wing-venation : Some modules may be exchanged between various derivational lines, but the venation as a whole of a given species of insect is not completely derived from that of another.
If this conception is correct, with respect to all morphological and other features of the insect body, then the grouping of insects, into taxa, is only a classification and nothing more. Such a classification, then, has no phylogenetic basis, because, according to our polyphyletic and noëtic theory there has been no phylogeny at all. Every morphological feature then has only a functional significance for and in the strategy concerned, that is, in the species concerned (biological species are true entities). That is what makes Rohdendorf's classification of flight devices in insects a cross-(insect-)Order classification. And as to wings in particular, i.e. in considering wings only (together with all the supporting structures), that is how far we can get. It may be detailed only a little bit further as Rohdendorf, 1951, has done with respect to the wings of Diptera, but no further. The differences in wing-venation must be for a large part functional, i.e. as a result of functional and morphological integration into the whole strategy in question. The venational pattern, in one or another way, determines (controls) the local bendings of the wing-blade during wing-beat. These local bendings co-determine the flight-regime of the representatives of a given insect species. That's about how far we can get in judging the differences in wing-venation.
As a result of our theoretical setting, the chief wing-veins cannot in fact be named or as such be identified, at least not so on the basis of a particular ancestral connection of species with earlier species. They can only be identified on the basis of rational morphology, and this is, in our opinion, the true state of affairs discovered by Comstock and Needham (1898-1899), namely the possibility of a uniform terminology of wing-veins in all insects. All this urges a new thinking in judging and interpreting the wing-venation in insects. But, in fact, it isn't a new way of thinking after all : Although one, in setting up or revising a classification of orgsanisms, often speaks of "ancestors", "phylogeny", etc., the majority of authors in fact are led by discovering morphological similarity, and apply it to their classification. It results in the so-called typological systems of classification of organisms (in contrast to "true phylogenetic" systems sensu Hennig and his adherents), in fact largely built upon 'rational morphology'. We ourselves adhere most to the attitude in these matters of the Russian authors, who, in setting up their typological classifications, take much into account the biology of the various species and groups, and that is correct because these biologies represent strategies to exist and persist, and so are constituted and supported by functional structures.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for a study of the general features of Inorganic Nature as the natural context of organisms and organic evolution, Part XV.
Back to first part of theoretic intermezzo
Back to second part of theoretic intermezzo Back to Part IX