e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Most expressed example of the type :
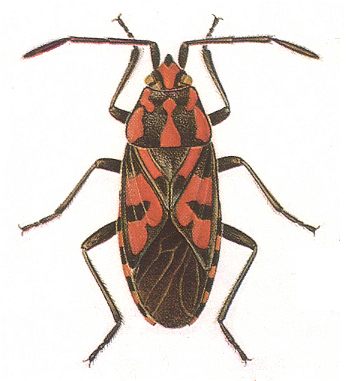
Figure 1 : Lygaeus saxatilis. Family Lygaeidae, Order Heteroptera. 10-11 mm. Recent.
(After SEVERA, in Thieme's insektengids voor West- en Midden-Europa, 1977.)
Description of the type
Morphological features.
Before we start with Rohdendorf's text on the matter we introduce figures and statements of other authors :
Heteropterygia is expressed in wings of which the forewings are themselves subdivided into a clearly stiffened proximal half and a membranous distal half. The hindwings are completely membranous with a moderately expanded anal area.
The very beginnings of the subdivision of the forewings can already be seen in the permian Prosbolidae :
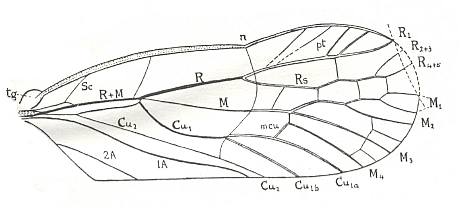
Figure 2 : Permoglyphis belmontensis Till. Family Prosbolidae, Order Homoptera. Upper Permian of Belmont, New South Wales, Australia. Tegmen, length 9.4 mm., greatest breadth 3.6 mm. n - nodus, tg - tegula.
From the nodus (n), a faint, curved, transverse line divides the tegmen as far as Cu2 into a somewhat roughened, granulate basal portion and a very smooth distal portion.
(After TILLYARD, 1926)
In the next Figure we show the definitive subdivision of the tegmen (the hemelytron, the partly sclerotized forewing) of recent bugs (Heteroptera) :
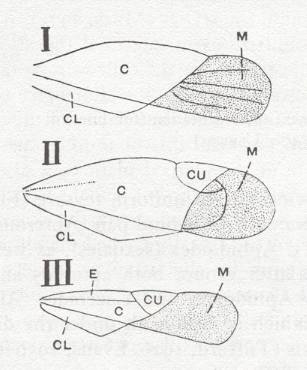
Figure 3 : Diagrams of hemelytra (in the Order Heteroptera, bugs) of :
I - a Lygaeid. II - a Mirid. III - an Anthocorid.
C - corium, CL - clavus, CU - cuneus, E - embolium, M - membrane.
(After RICHARDS and DAVIES, in Imms' General Textbook of Entomology, 1977.)
Most important of these differentiated regions of the forewings in bugs are the corium (sclerotized basal half of wing up to CuP [= vena dividens] ), the clavus (sclerotized anal area of wing), and the membrane (distal, membranous, part of wing). And, of course, especially important is the dividing line between corium and membrane. And all of these structures and regions are also visible in fossil tegmina of mesozoic bugs (Heteroptera), and already in those of certain permian Homoptera.
In Heteroptera the fore- and hindwings are coupled in flight by a simple apparatus, meaning that the forewings do not only have a shielding function, but participate, together with the hindwings, in flight- (wingbeat-) movements.
In order to have an idea of how these subdivisions (differentiations) in the forewings did originate, and especially how to see these subdivisions functionally, that is, to see the mixture and interaction of functional with non-functional structures in the tegmina of certain Homoptera and especially in Heteroptera, it is instructive to quote a page or two from TILLYARD, in "Mesozoic insects of Queensland, No, 4, Hemiptera Heteroptera : The family Dunstaniidae. With a note on the origin of the Heteroptera", published in The Proceedings of the Linnean Society of New South Wales, 1918, Vol. XLIII.
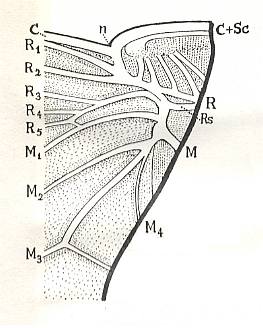
Figure 4 : Region of node (n) and dividing line in hemelytron of Dunstania pulchra Till. (Upper Trias of Ipswich, Queensland, Australia), showing the small anterior portion of the corium preserved in the fossil (hight of shown part of fragment, a little less than a centimetre). The thick black line bordering the right side of the drawing is not the dividing line (between corium and membrane) but the edge of the fossil. The true dividing line starts at the nodus. The apex of the wing (not shown) is to the left. (After TILLYARD, 1918)
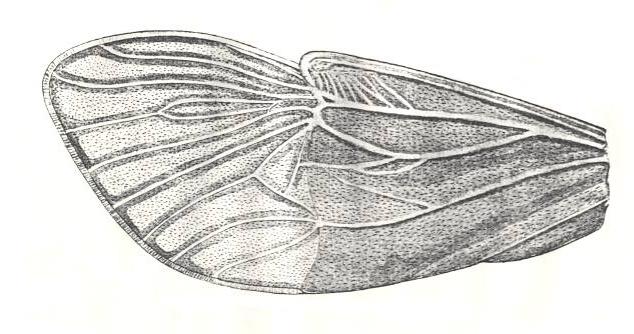
Figure 5 : Restoration of right hemelytron (forewing) [drawn with apex to the left in order to facilitate comparison with previous Figure] of Dunstania pulchra Till. (Upper Trias of Ipswich, Queensland, Australia), based upon the preserved portions of this fossil and of the allied Dunstaniopsis triassica Till. Length of hemelytrom about 2.8 cm. (After TILLYARD, 1918)
(pp. 590)
Let us now consider the very interesting question of the evolution of the dividing line between corium and membrane in the Suborder [or Order for that matter] Heteroptera. Dunstania shows us this line in an exceedingly primitive condition. From it, we see that its anterior portion was originally composed of either branches of main veins, or cross-veins. But, as the main veins are all proceeding more or less distad, it is, therefore, inevitable that these various parts should meet at different angles, as we see so definitely shown in Dunstania. (see Figures 4 and 5 ).
The parts of the dividing line (see same Figures) may be shown as follows, starting from the node or costal end [of that line] :A. Anterior Division, formed from cross-veins and branches of main veins :
- The node, formed from the fused C and Sc.
- Radial portion, formed from short parts of R1 and R2, partially fused together.
- A short cross-vein connecting the radial portion with the median portion.
- Median portion, consisting of a swollen root on M, from which the three branches of M are given off.
B. Posterior Division, consisting of a sinuous line crossing the wing more or less transversely, and not carrying either cross-veins or parts of main veins :
- Medio-cubital portion, from the swollen root of M to Cu. The curvature of this part is concave to the corium.
- Cubito-anal portion, from Cu to the antinode [opposite of the node on the other side of the wing]. The curvature of this part is slightly convex to the corium.
In the evolution of such a type as a recent Lygaeid (see Figure 3 I ) or Pentatomid bug, all these diverse parts have to become aligned and fused into one harmonious whole. Probably a careful study of existing types amongst the Heteroptera would reveal many traces of the method by which this change has been brought about.
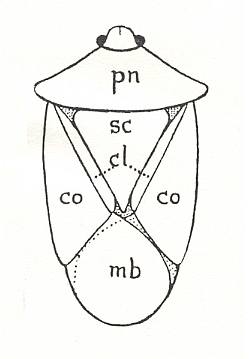
The other point of outstanding interest in the evolution of the heteropterous hemelytron is the development of the clavus, which is correlated with the growth of the scutellum [a small triangular dorsal shield of the thorax, sc in next Figure].
Figure 6 : Diagrams of a shield-bug, family Pentatomidae, Order Heteroptera, to show the five parts of the shield, viz., 1. the pronotum, pm. 2. the scutellum, sc. 3 and 4, the right and left coria, co. 5. the two completely overlapping membranes, mb (see also Figure 1 ). The clavus, cl, borders the scutellum on either side.
(After TILLYARD, 1918)We are bound to assume that, in the original Hemiptera [as a taxon representing the Homoptera + Heteroptera], the scutellum was of more normal size than it now is, and, consequently, the wings were not so completely folded over one another distally as they are now (see Figure 1 ). In fact, we have to start from the normal type of scutellum and the roof-like position of holding the wings [at rest], still to be found in many Homoptera of the present day. From this, a gradual enlargement of the scutellum, together with a close folding-down of the wings upon one another, leads us to the condition found in recent Heteroptera. Here the dividing line marks the limit of the portions of the wing that are folded upon one another, while the anal area, or clavus, borders the two posterior sides of the large triangular scutellum. Thus we have to correlate the length of the clavus with the size of the scutellum, the tendency being towards enlargement as evolution progresses. Since, in Dunstaniopsis [another fossil dunstaniid], there is no trace of a clavus in the preserved part of the wing, and the same is true of Prosbole [however, this genus turned out to have a clavus in the forewings, as does Permoglyphis of the Prosbolidae], we have to conclude that both these forms [Dunstaniidae, Prosbole] had only a small or moderate-sized scutellum, bordered by a correspondingly short clavus, or even, perhaps, by scarcely any claval area at all [but as our knowledge today says, this was the case in the (triassic) Dunstaniidae only, while the true heteropterous configuration of scutellum and clavus was already well developed in the permian Prosbolidae] .
As soon as the overlapping of the two distal parts of the wing, or membranes, has been brought about, it will be clear that these two parts will in future act (while the insect is at rest) as one [part] only, whereas the coria of the two wings remain separate (see Figure 6 ). There will, therefore, be little tendency towards the thickening of the membrane at all, and the "heteroptery" might be expected to advance more quickly than ever. The line of evolution would then culminate in forms having a thick corium, from which all traces of venation had been eliminated, and a thin membrane, in which the veins were arranged more or less parallel to one another and to the wing-border. Probably the highest point reached by this line of evolution to-day is to be seen in the dominant family Pentatomidae, in which the shield-shaped or "cut-into-five" appearance of the insect becomes perfected, and is often enhanced by bizarre sculpture and brilliant colouration. Figure 6 shows the outline of such an insect, with the five separate areas of the shield-design named.
( Now continuing with Rohdendorf's text -- but adding statements, clarifications, and Figures -- concerning the morphological features in the description of the heteropterygian type of wing-apparatus)
There exists a sharp heteronomy in the pairs of wings. Mesothorax more strongly developed than metathorax. Musculature also heteronomous. Forewings having quite a peculiar structure and divide into clearly separated parts of different consistency -- a firm leathery part, lying at the proximal half of the wing, and a thin membranous part making up the distal part of it. The venation of the forewings plays a markedly subordinated role as supporting element. It is rather variable and best developed in the distal, membranous, part. The forewings are named half-shields, hemelytra. The hindwings are membranous, fairly broad, but shorter than the forewings, provided with a strong rather reduced venation. The anterior margin is straight, connected with the posterior margin of the forewing by way of special adaptational structures, hooks. Both pairs of wings peculiarly specialized mechanically. Body usually short, not possessing flexible parts. Size of the insects small or medium [except certain larger Prosbolidae]. Wings shorter than, more rarely equal to body.
Functional features.
Both pairs of wings participate in flight-movements as organs of supporting the body and of traction. Wing-beats of both pairs strictly synchronous -- hind pair connected with the front pair by means of special adaptational structures and in wing-beat movements follwing it [i.e. following those of the front pair], functionally being its mere appendage. Speed of flight not studied. Known is only the wing-beat frequency reaching in the bug Deraeocoris schach (L.) 100-109 beats per second. Governability, weight, load and other properties of flight not especially investigated. The biological features of the representatives of the type are relatively well known. Significance of flight for the biology fairly great. We may see a certain parallel between beetles (Coleoptera) and bugs (Heteroptera) : As the majority of the former -- representatives of the type of elytropterygia (shield-wingedness) -- uses the wings only in certain periods of life, often leading a concealed way of life, also the bugs, representatives of heteropterygia (partitive wingedness), only take flight in certain episodes in their life. We must, however, indicate that heteropterygia is a markedly more sophisticated type aerodynamically than is elytropterygia.
Differentiations, connections, and representatives of the type.
Heteropterygia is a derivative of special forms of orthopterygia (straight-wingedness), expressed in Homoptera (cicadas), in which originated a subdivision of the forewing into two parts by means of a transverse seam. Some representatives of heteropterygia secondarily simplify the structure of the hemelytra, in which the apical membranous part is reduced. These forms connect heteropterygia with elytropterygia or even once again with the original type, orthopterygia [it is, perhaps, reasonable also to count neuropterygia as to represent one of the types from which heteropterygia has evolved].
The with respect to flight most progressive forms of heteropterygia are characterized by the improvement of the mechanical qualities of the hemelytra, which become narrowed and acquire the nature of costalized wings. These forms are a clear transition to dipterygia (functional two-wingedness). The various cases of differentiation in heteropterygia naturally split up into three subtypes.
1. Proheteropterygia
The most simple cases, closely connected with the homorthopterygia of cicadas (which is [one of] the starting form [s] of the present type) consist in the appearance of a clear "dividing" line, partitioning the tegmen into two parts. This subtype, proheteropterygia (pro-hetero-pterygia), we already find in certain mesozoic cicadas (Cicadoprosbole B.-M.). [We definitely also include in this type and subtype the permian genus Prosbole and allies (also Homoptera) in which this dividing line can already be found.].
In the recent fauna splendid examples may be seen in some Cicadidae (for instance Tettigarcta).
This subtype, foreshadowing the wings as they are in the next subtype, euheteropterygia, is very interesting because it is especially represented already in the permian Prosbolidae (Homoptera) (not mentioned as such by Rohdendorf). In that family of ancient cicadas the differentiation in the forewing (tegmen), by a dividing line, into a leathery and a membranous part is clearly present. So it will be instructive to include a number of Figures of tegmina and hindwings, including reconstructions of these ancient insects :
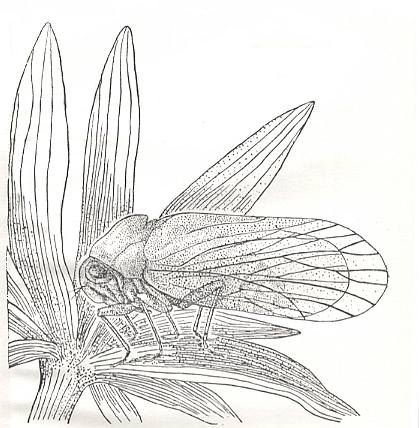
Figure 7 : Permocicadopsis angustata Mart. Family Prosbolidae, Order Homoptera. Reconstruction of the general view of the insect. Permian of the Archangelsk Region, Russia. (After BECKER-MIGDISOVA, 1940)
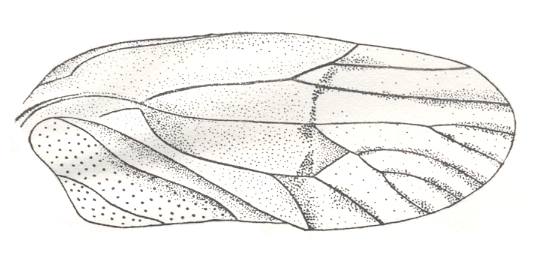
Figure 8 : Prosbole propinqua Mart. Family Prosbolidae, Order Homoptera. Tegmen. Upper Permian of the Archangelsk Region, Letopala, Sojana river, Russia. Length of tegmen 21.0 mm. Width 8.5 mm. (After BECKER-MIGDISOVA, 1940)
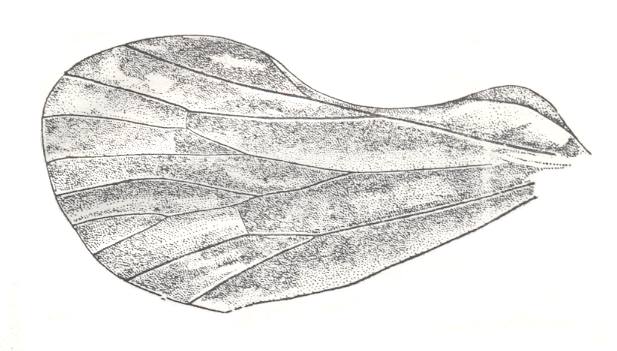
Figure 9 : Prosbole brevialata B.-M. Family Prosbolidae, Order Homoptera. Hindwing. Upper Permian of the Archangelsk Region, Iva-Gora, Sojana river, Russia. Length of wing 20 mm. Supposed length of tegmen about 22 mm. (After BECKER-MIGDISOVA, 1940)
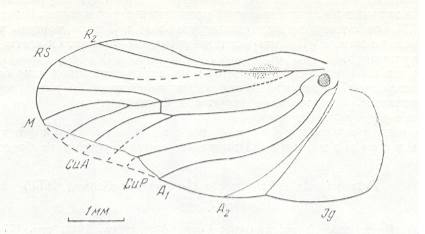
Figure 10 : Permocicada kuznetskiensis B.-M. Family Prosbolidae, Order Homoptera. Hindwing. Upper Lower-Permian of the Basin of Kuznetsk, Kaltan, Siberia, Russia. Length of wing 5 mm. Width 2.7 mm. Wing transparent, with weakly broadened anal and broad jugal areas. (After ROHDENDORF et al., 1961)
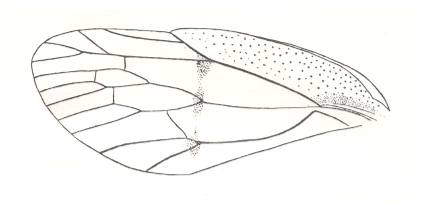
Figure 11 : Prosbole curvimarginata B.-M. Family Prosbolidae, Order Homoptera. Tegmen (forewing). Upper Permian of the Archangelsk Region, Sojana river, Iva-Gora, Russia. Length of tegmen 24.5 mm. (After BECKER-MIGDISOVA, 1940)
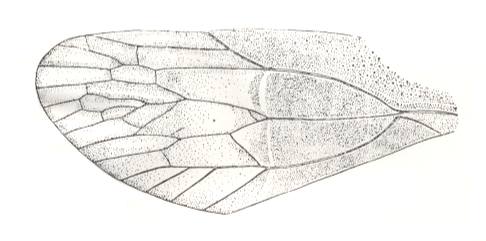
Figure 12 : Prosbole cellulifera B.-M. Family Prosbolidae, Order Homoptera. Tegmen (forewing). Upper Permian of the Archangelsk Region, Sojana river, Letopala, Russia. Length of tegmen 30.9 mm. (After BECKER-MIGDISOVA, 1940)
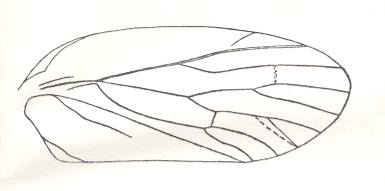
Figure 13 : Permocicada idelensis M. Zal. Family Prosbolidae, Order Homoptera. Tegmen (forewing). Upper Permian of the Archangelsk Region, Sojana river, Iva-Gora, Russia. Length of tegmen of species 9.45-12.0 mm. (After BECKER-MIGDISOVA, 1940)
After having figured and described many Prosbolidae, BEKKER-MIGDISOVA (1940) writes (p, 29/30) :
"As is evident from descriptions to be given later, the family Prosbolidae in general, and the genus Permocicada in particular, have much in common with recent singing-cicadas (of the family Cicadidae) as to the structure of the head, legs and body, as well as to the venation and structure of the wings. If we compare the venation of the tegmina in Prosbolidae [for instance that given in Figure 13] with that in the recent cicada Dundubia mannifera [next Figure], then, in the latter we find [as in Permocicada] R (which [in Dundubia, however] ends up at the wing apex), R2, RS [i.e. we firstly find the unbranched Radial Sector splitting off from R, and then, further distad, R itself again forking, giving R1 and R2 (as they are called in Homoptera) ] and a 4-branched Media. CuA differs a little in shape. The anal field [of Dundubia] has been decreased, and also the overall shape of the tegmen is different : The tegmen is narrower and stretched-out. The general plan of the venation, however, is the same. In addition it is possible that the oblique vein running between M1+2 and the base of RS is nothing else than the thickened dividing line. Near the wing's hind-margin this dividing line is preserved as a trace. This reminds us of the same such line in the genus Sojanoneura (for example S. edemskii Mart. and S. kazanensis Zal. [see Figure 15], running between RS and M1+2 [i.e. from RS to M1+2], and also [between] M3+4 and CuA [i.e. from M3+4 to CuA], having the nature of cross-veins."To this we ourselves would like to add that although the wings of cicadas like Dundubia do not belong to the present type (heteropterygia) but to a quite different type, namely to the type dipterygia (functional two-wingedness), the general plan of the wing-venation of these recent cicadas has remained relatively unaltered, as compared with that of the permian Prosbolidae, that is, although functional characters, connected with dipterygia, have altered the shape of the forwing (and the relative size of the hindwing) and have altered certain details of the venation, the general plan of the latter has survived these changes.
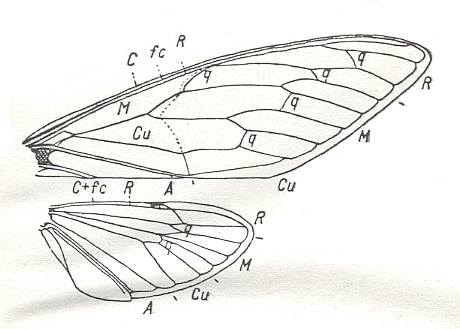
Figure 14 : Dundubia mannifera L. Family Cicadidae, Order Homoptera. Recent. Venation of fore- and hindwing. q - cross-vein (Querader). (After HANDLIRSCH, in BECKER-MIGDISOVA, 1940)
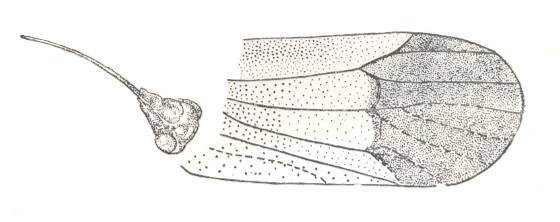
Figure 15 : Sojanoneura kazanensis Zal. Family Prosbolidae, Order Homoptera. Upper Permian of Archangelsk Region, Sojana river, Letopala. Russia. Tegmen and head. (After BECKER-MIGDISOVA, 1940)
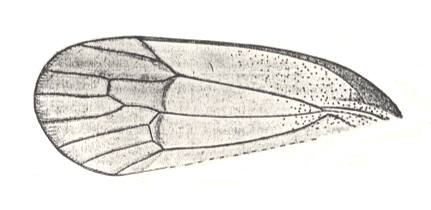
Figure 15a : Tegmen of Sojanoneura marginata Mart. Family Prosbolidae, Order Homoptera. Upper Permian of Tichije Gori, Ural, Russia. Length of tegmen 15 mm. Clavus not preserved. (After MARTYNOV, 1931)
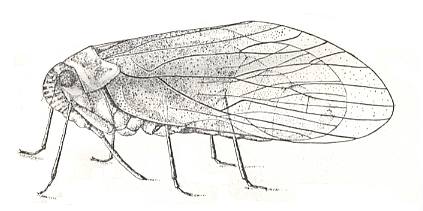
Figure 16 : Permocicada integra B.- M. Family Prosbolidae, Order Homoptera. Upper Permian of Archangelsk Region, Sojana river, Iva-Gora. Russia. Reconstruction. Length of tegmen 16 mm, width 6.3 mm. (After BECKER-MIGDISOVA, 1940)
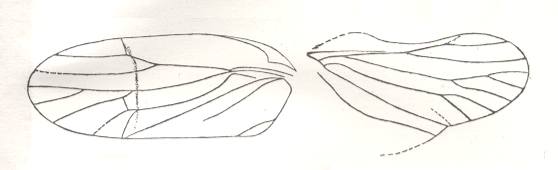
Figure 18 :
Left : Permocicada pusilla B.- M. Family Prosbolidae, Order Homoptera. Upper Permian of Archangelsk Region, Sojana river, Iva-Gora. Russia. Tegmen. Specimen 94/688. Length of tegmen 6.8 mm. Width 2.6 mm.
Right : Permocicada pusilla B.- M. Family Prosbolidae, Order Homoptera. Upper Permian of Archangelsk Region, Sojana river, Iva-Gora. Russia. Hindwing. Specimen 94/688. Length of hindwing 5.55 mm. Width 3.5 mm.
(After BECKER-MIGDISOVA, 1940)
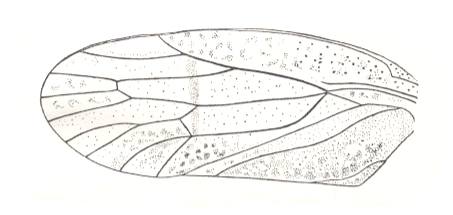
Figure 19 : Permocicadopsis letopalensis B.- M. Family Prosbolidae, Order Homoptera. Upper Permian of Archangelsk Region, Sojana river, Letopala. Russia. Tegmen. Length of tegmen of species 11.45-13.9 mm. Width 4.55-5.5 mm. In one of the specimens (the one depicted) the dividing line is clearly marked (dark transverse band). Sculpture and pigmantation (maculation) can be made out. The latter not so markedly. (After BECKER-MIGDISOVA, 1940)

Figure 20 : Permocicadopsis angustata Mart. var. typica. Family Prosbolidae, Order Homoptera. Upper Permian of Archangelsk Region, Sojana river, Iva-Gora. Russia. Tegmen. Found 15 specimen of this variety. Length of tegmen from 8.6 up to 10.15 mm. Width from 3.45 up to 4.0 mm. (After BECKER-MIGDISOVA, 1940)
2. Euheteropterygia
The great majority of bugs, Heteroptera, possessing almost triangular, broad tegmina (hardened forewings), [these tegmina] being divided into (1) firm leatherly parts representing the larger part of the wing and (2) a thinner almost membranous wing tip, represents the true partitive wingedness or euheteropterygia. See Figure 1 and also Figure3, and also this Figure :
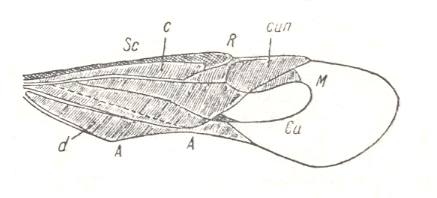
Figure 21 : Tegmen (forewing) of Calocoris sp. (Miridae, Europe).
d - clavus, cun - cuneus, c - corium.
(After HANDLIRSCH, in ROHDENDORF, 1949)
3. Diheteropterygia
One group of bugs, namely the representatives of the families Mesoveliidae, Hebridae, and some Reduviidae (from the subfamilies Tribelocephalinae, Holoptilinae, Stenopodinae) possess tegmina in which the compact leatherly parts significantly decrease as to their size and become narrow (as to their form) resulting in a very well expressed costalization (crowding of veins toward the anterior wing margin) of the wing as a whole. These forms of heteropterygia undoubtedly are of the most advanced nature from an aerodynamic point of view, and must represent their own subtype, costalized partitive wingedness or diheteropterygia. See next Figure.
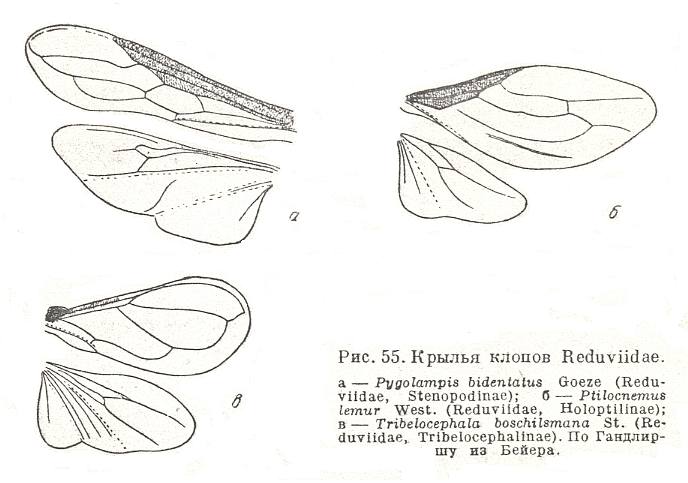
Figure 22 : Wings of bugs of the family Reduviidae.
Upper left - Pygolampis bidentatus Goeze (Reduviidae, Stenopodinae).
Upper right - Ptilocnemus lemur West. (Reduviidae, Holoptilinae).
Below - Tribelocephala boschilsmana St. (Reduviidae, Tribelocephalinae).
(After HANDLIRSCH, in ROHDENDORF, 1949)
* * *
What now follows is a two-document theoretic intermezzo on noëtic theory of evolution, and in Part IX, Strepsipterygia, we continue our exposition of wing types.
e-mail : ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for a theoretic intermezzo (two documents).