e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
In previous Part of Website ( Fifth Part of Website ) we have developed our noëtic theory of organic evolution. We'd done this by studying the evolution of the insect Orders Diptera and Hymenoptera from a most general point of view. And in order to further develop and test this theory, we will now, in the present Part of Website, consider the evolution of all insects, including their successive appearance -- as documented by fossils -- from carboniferous times all the way up to the present. For reasons already given (at the end of Fifth Part of Website) we will concentrate on the wings and flight-function of the insects in the context of their way of life, that is, in the context of their existential conditions in the Explicate Order. So the structure of the wings of insects is considered to determine the various flight-regimes, and these in turn, among other features that do so, reflect the way the adult insect moves around in its proper environmental medium. We may also say that the particular flight-regime co-determines how the insect experiences its direct environment (together with its eyes and antennae). In Fifth Part of Website we expounded the many functional wing-types in the Order Diptera. For Hymenoptera we did not do this because the wing-structure in that Order is much less diversified than it is in the Order Diptera. Only a few such types would be discernible in Hymenoptera. On the other hand there are insect Orders, such as the Order Hemiptera (bugs), the Order Neuroptera (Planipennia) (such as lace-wings and ant-lions), and the vast Order Lepidoptera (butterflies and moths), in which the range of diversity of wing-types is apparently much greater than in the Order Diptera. But this statement is in need of a little qualification. The number of wing-types present in the Order Diptera is probably greater than in any other insect Order. However, the morphological amplitudo -- the degree of difference between the types -- is, as it seems, much greater in the mentioned Orders (Hemiptera, Neuroptera, Lepidoptera) than it is in the Order Diptera.
The Class of [primarily winged] insects divides into two large groups or divisions, viz., Palaeoptera and Neoptera, precisely defined by the great Russian paleo-entomologist A. MARTYNOV, especially in his work entitled Otjerky geologitjeskoy istorii and filogenii otrjadov nasjekomich (Pterygota) [Studies on the geologic history and phylogeny of the orders of insects (Pterygota) ], Part One, Palaeoptera and Neoptera-Polyneoptera, 1938. The Palaeoptera are characterized by having their wings, also in their resting position, spread out. The Neoptera, on the other hand, have their wings, in resting position, folded back alongside or over-above the abdomen, either in roof-like fashion, or horizontally or sometimes vertically. In order to be able to do this they possess especially adapted structures at the base of their wings. In many originally neopterous insects the wings have been lost, but these insects nevertheless are considered to belong to the Neoptera.
Whereas the Palaeoptera -- paleozoic Palaeodictyoptera, and some other fossil groups, fossil and recent Mayflies (Ephemeroptera [= Plectoptera = Agnatha] ) and Odonata (damselflies and dragonflies) -- show only a few wing-types (such as long-wingedness [dolichopterygia] and paddle-wingedness [kopepterygia], it is in the Neoptera where we find the most diverse wing-types (such as, shield-wingedness [elytropterygia], straight-wingdness [orthopterygia], broad-wingedness [platypterygia], twofold-wingedness [dipterygia], etc.). And because the recognition of the Neoptera as a large division of insects is based on certain structures of their wings -- and in this way the qualitative content of this group predetermining the possible wing-types that can be formed in its representatives -- it is paramount to become more familiar with the Neoptera (to which, by the way, the great majority of insects belong!). And this, of course, can best be done by quoting the relevant pages in the mentioned book of Martynov (especially, because Martynov, like Rohdendorf, sees, in contrast to many western authors, evolution be driven by ecological factors, and thus not by random genetic mutations and natural selection). We will translate (from the Russian) [with additions and comments where necessary, in square brackets] the pages 66-70 :
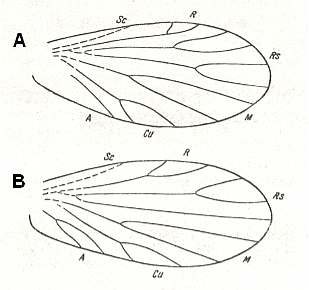
DIVISION NEOPTERAThis division includes insects from the very beginning of their history having acquired the ability to fold, in resting position, their wings roof-like or horizontally on their back. In connection with this, in the wings at their base separated off a special small jugal area, [see for this area (Jugum) and the jugal vein (Ju) next Figure],
which in further evolution of the group [the Neoptera] acquired enormous significance in increasing the area of the hindwing. It provides it with a usually special membranous area, called by me "neala" (1923). And after this special area I have called the whole division of these insects Neoptera.
This division, in upper carboniferous times being, as to its size, almost equivalent to the division Palaeoptera, in the lower permian forces the latter to second place, to hold first place from the upper permian onwards. In the mesozoic the Palaeoptera diminished to two Orders still living today. The Neoptera, on the other hand, continued to differentiate, and today includes practically the whole mass of Pterygota [subclass of winged insects] of the Earth's surface, adapted to live in the most diverse conditions.
In both up to the present preserved Orders of Palaeoptera, -- dragonflies and mayflies -- their nymphs [as the juvenile stages are called in insects with incomplete metamorphosis] inhabit fresh-water basins in which they also spend a large part of their life, while as adult, especially the mayflies, living very briefly.
Neoptera in their majority adopted a purely terrestrial life, but also among them there are some groups, of which the nymphs and larvae [as the juvenile stages of insects with complete metamorphosis are called] live a purely aquatic life, and it is not superfluous to note that those groups among their relatives distinguish themselves by the preservation of a number of primitive traits. Such are the Plecoptera [stoneflies], Megaloptera [alder flies], Trichoptera [caddis flies], many Diptera [two-winged insects], and Coleoptera [beetles]. The preservation of stoneflies (Plecoptera) is mostly due to the ecology of their nymphs, living in cool running water. In passed times, especially in paleozoic times, the relative number of aquatic groups was much higher.
The division of recent Neoptera neatly divides into three vast groups, which we called (1923, 1925) subdivisions, namely the subdivision Polyneoptera, the subdivision Paraneoptera, and the subdivision Oligoneoptera.
These subdivisions are easily distinguished by different structural features, way of life and metamorphosis, but in connection with the fact that the classification of insects generally, in a considerable degree goes its way by the features of their wings, and also in connection with the fact that fossil forms are known to us largely by these organs, we also for a general differential characteristic of the subdivisions have taken the wings as a basis and have given to these subdivisions the corresponding names. The subdivision Polyneoptera corresponds to the whole of the orthopteroid insects [grasshoppers, cockroaches, etc.] in the old broad sense of the term plus the stoneflies (Plecoptera) and the Embioptera [gregarious insects living in silken tunnels]. The latter were sometimes placed near the booklice (Copeognatha [= Psocoptera] ) and even near dragonflies and mayflies (Pseudoneuroptera), but today not only the stoneflies but also the Embioptera, together with their fossil relatives to belong to the Polyneoptera is without doubt. As regards the orthopteroid insects in the broad sense, they were subdivided already by Handlirsch (1908, 1925) into two superorders -- Orthopteroidea and Blattaeformia. In the Blattaeformia he not only placed the cockroach-like insects, i.e. Blattodea [cockroaches], Mantodea [preying mantises], and Isoptera [termites], but also the psocids (Copeognatha or Psocoptera), Mallophaga [biting lice or bird lice], Siphunculata [= Anoplura, sucking lice], and Zoraptera. We cannot in any way agree with the inclusion into one single superorder with the cockroach-like insects also the four latter Orders, and already in 1925 (partly also in 1923) we have placed them closely to the superorder Hemipteroidea (Rhynchota) [sucking insects, such as bugs], having set up [for them, i.e. psocids, lice, etc. plus Hemipteroidea] a special subdivision Paraneoptera. Crampton already long ago pointed to certain similarities in the wing-venation of the Psocoptera and the Hemipteroidea [Hemiptera]. In recent times Tillyard and Carpenter considered the closer relationship of these groups [to be probable], and, finally, also Lameere inclined to it. In his most interesting survey of zoology, 1936, he divides his Neoptilota=Paurometabola into two parts : Orthoptera and Hemiptera, where in the first he counted two groups -- Polynephridia, as regards size corresponding (in recent groups) to our Polyneoptera, -- and Oligonephridia, corresponding with our Corrodentia [meant are, apparently, psocids and lice] with this difference that he includes in the latter group also the Thysanoptera [thrips], which we, with the present state of our knowledge of this group, have not with confidence decided to include in the Paraneoptera.
In the introductory statements to the survey of Hemiptera, however, he explicitly says (p. 383), that "they (i.e. Hemiptera) may be unified with the Oligonephridia. They possess only four malpighian tubes , three tarsal joints, and have lost the cerci [abdominal end-threads]. They suck, though not in the way Siphunculata and Thysanura do, but differently".
The similarity between the Psocoptera (psocids) and the Hemiptera [Hemipteroidea], in their wings, the structure of the head (the presence of a large head shield [clypeus] in Psocoptera and Hemiptera), and the jaws, and also in the three-segmented tarsi, not to speak of the low number of malphigian tubes, -- all this definitely bears witness to the close relationship of the psocids together with the neighboring to them parasitic Orders [lice], and probably also the Zoraptera, on the one hand, with the Hemiptera on the other, closer that is, than to the Polyneoptera. As to the structure and venation of the n e a l a, where it is developed, the Paraneoptera very significantly, even sharply, are distinct from Polyneoptera, and therefore we cannot unify the Paraneoptera with the Polyneoptera. When not putting together the Copeognatha with the Hemipteroidea, then yet for them [the Copeognatha] we have to create a special subdivision, for which there is of course much in favor, but taking into account the presence of a number of features connecting them to the Hemiptera we do not, for the time being, see in this a necessity. To our subdivisions we grant the meaning of being the largest units into which only may divided the Neoptera. All Polyneoptera, despite their extreme diversity, we take as a single subdivision, as also we take as a single subdivision all insects with complete metamorphosis (Oligoneoptera or Holometabola). In such a state of affairs the mutually related large superorders Hemipteroidea and Psocopteroidea -- (earlier we called them Corrodentia, but under this name one thinks usually of psocids only, while not including Mallophaga, Anoplura [= Siphunculata, sucking lice] and Zoraptera), -- we, as we did before, favor to unify into one single large subdivision Paraneoptera.
As we will see, the general classification, chiefly of recent insects, that we in 1925 proposed on the basis of wing-structure, and in a high degree based on the structure of the Jugal area of the hindwings, is more and more being confirmed in comparative analysis of other organs.
In Polyneoptera the posterior Jugal area either, if it is sufficiently well developed, includes a few, sometimes a whole series of longitudinal veins (venae jugoradialia), or the Jugal vein almost directly after its beginning forks (Plecoptera). In Oligoneoptera the Jugal area of the hindwings, no matter how well it is developed, contains not more than one true longitudial vein (vena arcuata), being always simple, without branches.
In Paraneoptera the n e a l a [the Jugal area], where it is developed, also contains one single vein, but this vein may sometimes fork in its distal part. As regards this, the Hemipteroidea, as we will see, differ less from Oligo- than from Polyneoptera. However, in other respects, in wing-structure as well as, and more so, in other traits of the organization, they strongly differ from Oligo- as well as from Polyneoptera. The Psocoptera are close to [both of] them, but because they are in various respects held back so to say in a very original stage they do not differ so much from the other subdivisions. But their status [i.e. their place in the classificatory system] is easily clarified by comparison of other structures and other organs.
One may ask whether we can truly not only characterize but also name our complexes by the terms Polyneoptera, Oligoneoptera and Paraneoptera, because under these terms do not go all groups that belong here. There are indeed exceptions, aren't there? We think it is possible, because similar "exceptions" are always present, and the task of classifying animals as evolving groups consists not in erecting artificial, albeit precise, rubrics as in a book catalogue, but consists in the separation, in the typification, and in the corresponding indication of [genealogically] related complexes having split off from others in earliest times. Their kinship, the presence of common and similar inherited [structural, morphological] beginnings in further evolution and in similar conditions inevitably, as a rule, leads to parallel formation of similar structures, and on the basis of precisely them we usually judge their degree of kinship. In the individual components of a given developing tribe these or those structures may, as a result of these or those causes, usually external, not develop or not finally develop, but from this does not follow that they should be placed apart [in the system], because as to their basic features they are built according to the very same plan. In these cases it is paramount to clarify the origin of these deviations.
With the terms Polyneoptera, Oligoneoptera, and Paraneoptera we wanted to express the very characteristic, and easily presentable in the evolution of the different groups, features appearing in the wings in a parallel way on a grand scale.
First, in the ancestors of all three subdivisions the hindwings were wholly similar to the forewings (o r i g i n a l h o m o n o m y) and had in a stronger or lesser degree the nature of t e g m i n a (stiffened wings). Distinguishing in them three future subdivisions based on the wing-venation would evidently be very difficult, -- just as it is dificult to distinguish future organs based on early beginnings of them. When, however, in our Neoptera clearly became visible the demand of increasing the surface area of the originally homonomous wings, in jumping from plants or rocks to the ground, then everywhere in a parallel way a broadening of the hindwings began to take place, beginning from the posterior part. This broadening of the hindwings began in all subdivisions of the Neoptera, in the most diverse groups, and everywhere membranization and broadening started off from the jugal and anal areas. This generality of the process indicates common causes which also consisted in the development of gliding and flight.
However, as is perfectly clear, different large groups, having already separated off in earlier times, reacted to the demand of broadening the wings in different ways, and these differences especially were expressed also in the jugal area. Of course, such differences appeared more clearly and sharply in later and larger representatives than in earlier ones. These differences of reactions, without doubt, come from the difference of inherited beginnings of large groups. They also determined the fact that in the different Orders of the Polyneoptera in parallel began to differentiate in the growing Jugal area some longitudinal veins, but in Oligoneoptera and Paraneoptera only one. But this single longitudinal jugal vein in many Hemiptera [members of the Paraneoptera] began to fork at its end. Although this process of formation of jugal veins went its way independently in the various groups, we may nevertheless, basing ourselves on the similarities in them, now, in a certain degree, decide also on the genealogic kinship of these and those groups [i.e. the features have evolved independently, but this was possible because the groups are genealogically related].
The process of broadening of the hindwings and the formation of jugal veins took place on a massive scale, but in all three subdivisions we find individual small, but sometimes also large groups in which this broadening either did not take place at all, or was expressed very faintly. This does absolutely not mean that they are totally separated groups. The evolutionary process of broadening could not go its way uniformly in all groups of the given large complex, because it finds itself under the control of various factors, chiefly of mechanical nature, which could not be uniform in different cases. Can we make these general factors responsible for the delay in the appearance of the broadening of the "jugum" or for the weak development of it? This problem is hard, but, I think, not unsolvable.
Martynov is now going to show cases in Neoptera where the neala - and with it the ano-jugal area -- of the hindwings is only little or not at all developed (or secondarily reduced). For seeing the difference it is instructive to first show two of the many cases where it is well developed :
Wings of Schistocerca sp. Catantopidae, Order Orthoptera, Neoptera. Tropics and subtropics, Recent. Length 50 mm.
Ano-jugal area of hindwings well developed. Wingpairs heteronomous.
(After BRUNNER in HANDLIRSCH, in ROHDENDORF, 1949)
Wings of Blattella. Order Blattaria, Neoptera. Recent.
Ano-jugal area of hindwings well developed. Wingpairs heteronomous.
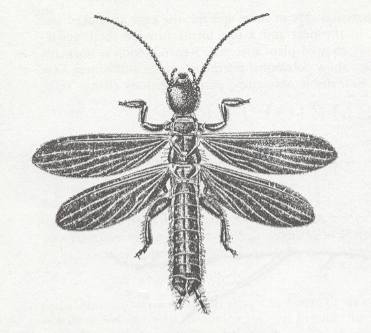
(After COMSTOCK, 1918, and REHN, 1951)For solving it, we must clarify in what feature precisely do all these groups [where the neala is weak or not developed at all] distinguish themselves from the others in the various subdivisions. In the subdivision Polyneoptera these groups will be the Embioptera, see next Figure,
Embia major, Embioptera, male, India.
[no developed neala in the hindwings]
(After IMMS, 1913, in Imms' General Textbook of Entomology, 1977)a large part of the termites,
A winged termite (Archotermopsis), Isoptera, male.
[no developed neala in the hindwings]
(After RICHARDS and DAVIES, in Imms' General Textbook of Entomology, 1977)and, perhaps, some stoneflies (Plecoptera), in [all of] which [groups] the broadening of the wing is very insignificant (partly here took place secondary reduction). All these groups distinguish themselves by [containing representatives of] especially small sizes. Of fossils one should place here [the Order] Miomoptera, which are also very small,
Fore (A) and hind (B) wing of Archaemioptera carbonaria Guth., Miomoptera, Archaemiopteridae, upper Carboniferous of Western Europe. [no developed neala in the hindwings] (After GUTHÖRL, in ROHDENDORF, 1949)
and certain small orthopteroids.
In the division or subdivision Oligoneoptera more usual are groups in which the hindwings very weakly or not at all were broadened in the ano-jugal region. To these belong, for example, the Orders Neuroptera
Right wings of Sisyra flavicornis, Neuroptera-Planipennia, Sisyridae. Recent.
(After COMSTOCK, 1918, in IMMS' General Textbook of Entomology, 1977)and scorpionflies
Panorpa communis, Panorpidae, Mecoptera. 20 mm Recent.
(After SEVERA, in Thieme's insektengids voor West- en Midden-Europa, 1977)and their derivatives, the Diptera. In other Orders of the Oligineoptera the ano-jugal part did not grow at all only in groups the representatives of which are small. From the Lepidoptera do belong here the Micropterygidae,
Wings of Sabatinea incongruella Walker, Micropterygidae, Lepidoptera. Recent.
(After TILLYARD, 1935, in HENNIG, 1969)Protoneuridae and related families, and then the Nepticueidae and some other families from the so-called Tineoda. In the remaining Frenata [= frenate lepidoptera], on the other hand, the jugal area experienced one or another broadening.
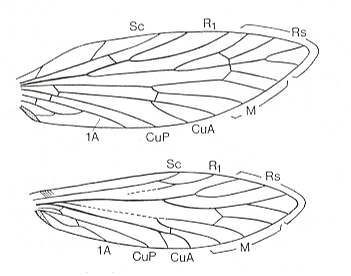
Among Trichoptera [caddis flies], next to families having a broadened ano-jugal area, we find neighboring families with more or less homonomous wings, and it is without doubt that such homonomy can be found chiefly in those families of which the representatives distinguish themselves by small sizes (Hydroptilidae, a large part of the Rhyacophilidae,
Venation of Rhyacophila fuscula, Trichoptera, Rhyacophilidae. Recent.
(After COMSTOCK, with legend slightly altered, in IMMS' General Textbook of Entomology, 1977)Philopotamidae, Beraeidae, Sericostomatidae, small Leptoceridae {genus Setodes} and others).Among Hymenoptera (belonging to the subdivision Oligoneoptera) the ano-jugal area was not at all increased in the hindwings (the jugal area was even completely reduced) in all parasitic forms and groups related to these, which originally were very minute insects, and also in some Aculeata, chiefly in ants [being also small insects].
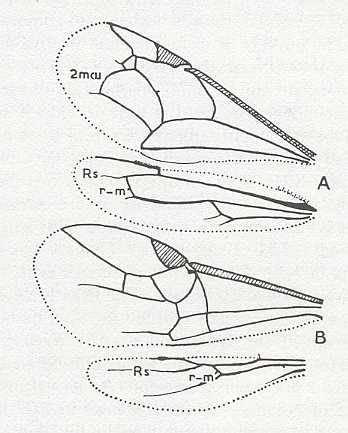
In order to clarify this we [JB] insert first some Hymenoptera in which the ano-jugal area of the hindwings is fairly well developed, followed by forms in which this is not so :
Wings of reconstructed ancestor of the Suborder Symphyta of the Order Hymenoptera. Ano-jugal area of hindwings developed. (After RASNITSYN, 1969)
Xyela julii Br., Family Xyelidae, Symphyta. Primitive Hymenopteron. Beginning of heteronomy of the wings (decrease of size of hindwings). (From BERLAND, 1951, in MALYSHEV, 1966]
Left wings of, A, an Ichneumonid, and, B, a Braconid (both belonging to the parasitic Hymenoptera).
Complete reduction of the ano-jugal region of the hindwings.
(After RICHARDS and DAVIES, in Imms' General Textbook of Entomology, 1977]In beetles (Order Coleoptera, also belonging to the Neoptera-Oligoneoptera), in connection with the fact that their forewings have the nature of elytra [shields], the hindwings, as a rule, are more or less broadened in their anal and jugal areas, but this broadening is extremely weak in many Staphylinoidea [short-shield beetles], which generally are distinguished by small sizes.
In the subdivision Paraneoptera the jugal area not only was not broadened, but even underwent a reverse development, first of all so in all Psocopteroidea, i.e. in Copeognatha (booklice, psocids), Zoraptera,
Peripsocus phaeopterus, Order Psocoptera (booklice).
Complete absence [or complete reduction] of the ano-jugal region of the hindwings.
(After SILVESTRI, in Imms' General Textbook of Entomology, 1977]
Right wings of Zorotypus snyderi, Order Zoraptera. pt - pterostigma.
Complete absence [or complete reduction] of the ano-jugal region of the hindwings.
(After RICHARDS and DAVIES, in Imms' General Textbook of Entomology, 1977]and -- presenting themselves to think about (as having had a reduced ano-jugal area) -- in the winged ancestors of the Mallophaga (biting lice) and Anaplura, in connection with the transition to parasitism having lost their wings. In Hemiptera [bugs], as a rule, in their hindwings has taken place a significant increase of the anal and jugal areas. Only the Homoptera-Sternorrhyncha did not undergo this process, i.e. precisely the groups (Coccidae [scale insects], Aleurodidae [White Flies], Psyllidae, and aphids) the representatives of which are distinguished by small, and often also minute sizes :
Psylla mali, Psyllidae, Order Homoptera.
Complete absence [or complete reduction] of the ano-jugal region of the hindwings.
(After CARPENTER, in Imms' General Textbook of Entomology, 1977]
Aphis fabae, Aphididae, Order Homoptera. Male.
Complete absence [or complete reduction] of the ano-jugal region of the hindwings.
(After DAVIDSON, in Imms' General Textbook of Entomology, 1977]The Superorder Thysanopteroidea [containing the (only) Order Thysanoptera, thrips] [see Figure below], by some (Lameere) having been placed close to the Psocopteroids, by others close to the Orthopteroids, possess extremely narrow wings, and to assume that even in their ancestors the hindwings, in the anal region, were broadened is out of the question. This Order entirely consists of minute forms, and many of them are even ultra-minute.
Taeniothrips inconsequens, Order Thysanoptera.
(In Imms' General Textbook of Entomology, 1977]From this concise survey the general picture becomes completely clear. In the process of working out first the ability of gliding from plants or rocks, and then true flight, undoubtedly there existed an inevitable demand of broadening the hindwings, but this expressed itself or was effective only in the larger and medium species, not in the very small forms. In these groups everywhere this process of broadening of the anal and jugal areas got started, going parallel with their membranization. The mechanical conditions of motion in the air, especially of flight [as distinguished from jumps], are highly different in very small insects [as compaired to larger ones]. Therefore it is not surprising that in them flight began to develop differently, and this was expressed in the structure of their wings. In groups consisting of small forms, the process of the indicated broadening of the hindwings was either weakly expressed or not at all. The corresponding groups, adapting to the special mechanical conditions during their flight, changed the structure of their wings differently. In many the necessity to increase the wing's surface area did not result in the broadening the blade of the hindwings, but in the elongation of a series of marginal hairs, i.e. in the formation of a so-called fringe. Along this evolutionary course went the Thysanoptera (see Figur above ), the Microtrichoptera, and the Microlepidoptera. In others almost the whole work in flight was assigned to the forewings, and the hindwings decreased or were completely reduced. In a third group the wings of the middle- and hindthorax began to work independently, etc.
Thus, the process of broadening the hindwings in the anal region took place as an elementary first event, i.e. in late devonic and carboniferous times in some [groups], and a little later in others (Oligoneoptera), on a massive scale. It did not occur, or took place differently, in groups with smaller representatives, by reason of mechanics. In various large groups this broadening took place, however, a little differently, and especially the venation in the increasing jugal regions formed differently, despite the commonness of mechanical factors.
These differences must be seen as an unveiling, in their latest stages, of the differences in their inherited [first] beginnings, acquired in ancient times. These differences are the more expressed (usually) the longer has lasted the evolution of the given organ.
Finally, different groups differ in the structure of the most different organs of different ancientry, but we hold that giving names and most concise diagnostic features is done perfectly appropriate on the basis of the structure of the neala of the developed forms, because the neala is a "new" part of the wing, in fact a certain adventive formation that appeared at the base of the anal region. After that it developed already independently within the ranges of the individual Orders and individual families, resulting, however, in similar forms within the ranges of separate large groupings. The name Polyneoptera signifies a complex of groups, in the hindwings of which, in cases of their more or less significant broadening, the jugal areas did form some radially divergent veins. To this complex also should belong those related groups of smaller insects, in which the mentioned process was slower or was not at all expressed, in connection with their minute sizes. ( End of quote from MARTYNOV, 1938).
In what follows we will now, following ROHDENDORF, 1949, establish types of wings as found in the Class of Insects. These wing-types, despite the fact that they are called "types", do not -- when classifying wing-types -- have the same rank as do have the wing-types as they have been established in Fifth Part of Website (there following ROHDENDORF, 1951) for the Order Diptera. In fact they are, relative to the latter, "super-types". We, however, keep calling them just "types" because of the awkwardness of terms such as "super-types". The ranking of types (and subtypes) will be clear from the context. So the wings of Diptera, representing the many types described in previous Part of Website, together are representatives of the more encompassing wing-type called "dipterygia" or two-wingedness. This type not only contains insects with only one pair of wings, such as all representatives of the Order Diptera, but also those (found in other insect Orders) in which the hind-wings are f u n c t i o n a l l y just a mere posterior lobe of the forewings. They function there as does the basal posterior portion of the wing (mainly the anal lobe) in the wings of Diptera. Such flight-organs each one of which morphologically consists of four wings but functionally of only two wings, we find, for example in many species of the Order Hymenoptera. There the hindwings are present but are definitely smaller than the forewings and are, during flight, attached with their anterior margin to the posterior margin of the forewings by a row of hooklets. So both pairs of wings are then one single functional unit. This dipterygial type (functionally two-wingedness) is also found in members of other insect Orders. And in addition to this type there exists a number of other types and subtypes of wings of representatives of several insect Orders, such as the types platypterygia (broad-wingedness) in many butterflies, ptilopterygia (feather-wingedness) in certain Diptera, Hymenoptera and Lepidoptera, and orthopterygia (straight-wingedness) in most Orthoptera (grasshoppers, locusts and crickets), etc. We will describe these types in due course.
In biological respect the whole order Hymenoptera is relatively well studied. The archaic Symphyta, possessing little-mechanized wings and a weak flight, are phytophagous forms, their larvae live independently, and moreover usually in open habitats. The larvae of the representatives of the vast, ancient suborder Entomophaga (Terebrantia) are parasites of other insects. The youngest group, geologically only relatively recently having reached its bloom, are the stinging Hymenoptera (Aculeata), which group is very characteristic as to the development of instincts of care for the young : The larvae of these insects develop in special nests prepared by the female. In some groups of aculeates we see the formation of special "social" forms of life. Such are the ants, social wasps and social bees. The winged mature insects of the aculeate Hymenoptera are therefore the most important developmental phase for the well-fare of the species. The adult insects of this group do not only have the function of reproduction [such as mating and laying eggs] and distribution, but also to supply food for them and for their larvae. Therefore the improvement of the quality of the flight-apparatus of the Aculeata is biologically a very important evolutionary process to the species.
In summarizing concisely the features of the evolution of the wings of the Hymenoptera, we must note the relative uniformity of these processes in the whole Order. Hymenoptera are characterized by a well-expressed "two-wingedness", which in some groups reaches an extreme. Together with the attainment of a great mechanical perfection in the structure of the wings, also has altered, with respect to strengthening, the muscular apparatus moving the wings -- the thoracic section of the insect-body is enlarged. The interesting rareness of cases of complete reduction of the hind pair of wings in Hymenoptera, apparently may be explained by the v e r y e a r l y-o n formation of true two-winged flight, i.e. a flight with a high wing-beat frequency, where the hindwings became a necessary element of the wing-apparatus, its posterior propelling membrane.
Dipterous insects show clear genealogic ties with the Order Mecoptera (Scorpion-flies) and the extinct Paratrichoptera, in today's systematics forming together with Lepidoptera (butterflies and moths) and Trichoptera (caddis-flies) representatives of the group Mecopteroidea (= "panorpoid complex" of Tillyard). Diptera are today one of the most widely distributed and rich in species Orders of insects, only coming after the Coleoptera (beetles), and competing with Lepidoptera and Hymenoptera, and, in fact, apparently, more rich in species than the latter two Orders which are much better studied. Moreover, as a result of a series of features of their organization and biology, the Diptera are, perhaps, the most specialized, "highest" group of insects. Such a conclusion is not only probable with respect to anatomic features of these insects, but also with respect to their individual development and biology : It is sufficient to remember the metamorphosis of various Diptera, often reaching an extreme fastness [in time] and a sort of perfection.
Such are the representatives of the group Pupipara, of the family Glossinidae (tsetse flies) and some others. They give birth to p u p a e or completely m a t u r e larvae. Larval feeding of these insects, consequently, is realized in the body of the mother. A similar kind of most useful adaptation may, with qualification, among other animals, be shown only in mammals and some other vertebrates. Moreover, vivipary (of young larvae) is widely distributed among the Order Diptera.
Diptera are very remarkable as to their features of flight. Their wing-apparatus has reached the highest degree of specialization on the evolutionary path to the improvement and perfection of the flight-ability -- the wings never assuming a protecting function [in the sense of protecting the body in periods when the insect does not fly]. Apterygia (= not having wings altogether) has a very limited distribution in the Order as compared to other Orders. Of Diptera, as does indicate the very name, the presence of the anterior pair of wings only, is characteristic. The hind pair is completely reduced as an organ of flight creating traction or support [lifting power] during flight. They are transformed into special, apparently sensory structures.
Cases of dipterygia are fairly numerous among winged insects, but do not in any group reach such wide distribution and such a complete morphological expression as in the Order Diptera. Loss of the hindwings in Diptera have, apparently, taken place already very long ago. Probably, the original group from which might have been split off the ancestors of Diptera, was very close to the extinct Order Paratrichoptera.
(end of quote)
So we have seen that the insect wing-types yet to be discussed may -- often through their subtypes -- have representatives in several different insect Orders. For example the type "dipterygia" (two-wingedness) is, in its most perfect form, realized in the Order Diptera (and in some separate forms in other Orders) by complete reduction of one pair of wings, while it is realized i n a n o t h e r f o r m in many species of the Orders Hymenoptera and Lepidoptera (namely in hawk moths and others). Here "dipterygia" consists in the hindwing's size being more or less reduced and in the fore- and hindwing functionally forming one single wing. Also other wing-types such as platypterygia (broad-wingedness) and dolichopterygia (long-wingedness) cross over from one insect Order into another.
When considering examples of wing-types and their subtypes -- in fact, when considering any species-constant feature in organisms -- we may determine their evolutionary condition, a condition as it exists (and as it is, or looks) within the particular natural group [of organisms] to which the organism -- representing such a type or feature -- itself belongs. Any such natural group, a taxon (such as a family or order), seems to inhere an evolutionary potential of its own, having, or not having, certain evolutionary possibilities in its members to be realized. So, for example, we saw that any form of "elytropterygia" (shield-wingedness) is absent in the Order Diptera. From this we may conclude that the very qualitative content of that Order is such that it cannot [evolutionarily] create, or is not compatible with, any form of shield-wingedness (such as has taken place in the Orders Coleoptera (beetles), Blatttodea (cockroaches), Dermaptera (earwigs), in many Orthoptera (grasshoppers and allies), in a way in Hemiptera (bugs), and even, albeit in a very light form, in certain Lepidoptera (namely in certian moths). On the other hand, any form of "dipterygia", as widely distributed it is in many Orders, does not occur in the Order Odonata (damselflies and dragon flies). So every order, family, etc. has its own particular qualitative content and therefore its own evolutionary potentials.
The evolutionary condition and status of a type, a subtype, or any species-constant feature in organisms may be one of the following :
Before describing the functional/morphological wing-types in insects and their distribution in the several Orders, it is parhaps instructive to present and figure the most general morphological features of the wings of insects, and their terminology.
General structure of the insect-wing
[ Most of what will follow (structure of wings, functional types of wings, characterization of insect Orders as to the type of wings possessed by their representatives) is taken [with changes and additions] from ROHDENDORF, Evoljutsia i klassifikatsia ljetatjeljnovo apparata nasjekomich ( Evolution and classification of the flight-appparatus of insects), in Trudy Paleontologicheskovo Instituta, Part XVI, 1949.] [ In this book Rohdendorf reviews many features of insect-wings and insect-flight. From this we only take the morphology of the wings. We do not here consider Rohdendorf's expositions and discussions of the articulating sclerites lying between the thoracic muscles and the wing-base, neither do we consider his expositions of the various flight-muscles, because this Part of Website is not supposed to be a textbook of entomology. Also the histology [tissue structure] of the wings, and especially the mechanics of the insect-wing during flight we do not consider here, because for these more recent litarature should be consulted if available. Of course data on shape and venation of the wing will be considered here, because these become the main theme of this Part of Website and because they are less vulnarable to become obsolete. Moreover, in fossil insects -- being used by us to clarify the evolution and successive appearance of insects during geologic time -- almost the only features that are generally well preserved are the shape and venation of the wing (chiefly forewings only) .
We also follow the mentioned book of Rohdendorf according to some other plan : Whereas Rohdendorf, -- after having expounded the generalities of wing-structure, musculature, alar sclerites, and a review of work on the mechanics of insect wings, -- starts with a review of all Orders of winged insects from the viewpoint of the different wing-structures occurring in them, and only after it embarks on the task of the description of functional wing-types and subtypes, we, in this Part of Website, after a brief introduction, start with the description of Rohdendorf's types and subtypes, and let it then follow by a review of the various insect Orders, while showing what wing-types in which Orders actually occur. So the insect Order is central in our treatment of the flight-function, and we will show the different capabilities of the qualitative contents of the different insect Orders of creating these or those functional wing-types, together with their different capabilities to occupy different ecological niches.]
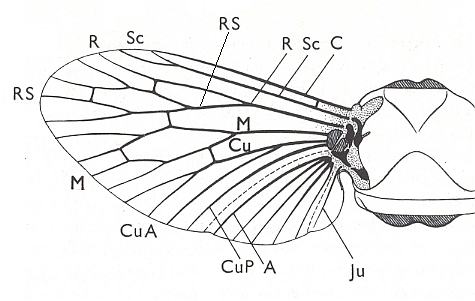
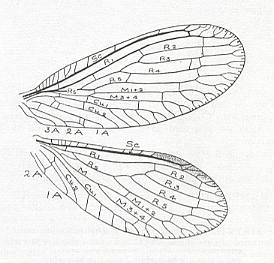
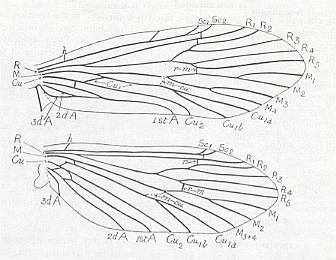
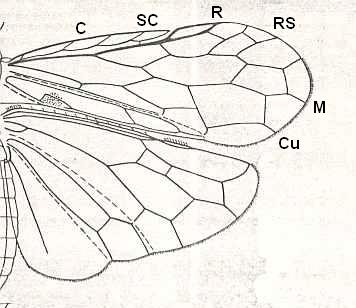
The structure of the wing of an insect has been a theme of investigations of numerous authors. As a result of their works, to us became known the general features of the morphology of wings in insects of the various Orders. The investigations of Comstock (1918) have provided the basis of assessing the homology of the skeletal elements of the wing, its venation. [And so did the works of Handlirsch (1906-8 and throughout the 20's and 30's), Tillyard (1919, and throughout the 20's and 30's) and especially of Martynov (in the 20's and 30's) ]. All these works formed the basis for further investigations of the morphology of the wings albeit not devoid of great insufficiencies. The basic, all-out general to the Class of insects, scheme of venation given by these authors -- costal, subcostal, radial, medial, cubital, and anal veins -- after a series of completions and changes remained the chief scheme, satisfactory expressing the true structural plan of the supporting apparatus of the wing, its venation.
Chief parts of the wing
The wing is an organ of the insect, the basic and chief function of which consists in the realization of flight -- the locomotion of the insect in the air by means of particular motions guaranteeing to overcome gravity and air resistance, and to create traction. In insects the wings also participate in controlling and steering of flight, and in keeping balance during it. For a successful performance of these functions there must exist sufficient surface area and a firm mechanism to transmit the strong and diverse muscular forces (in insects these muscles are contained in the thorax). Finally, in the majority of insects it is demanded that the wings be foldable, i.e. to take up the least possible place when the insect is not flying (its resting position) in order not to become a hindrance in another way of locomotion of the insect, for instance in swimming in the water, crawling in narrow spaces, and so on.
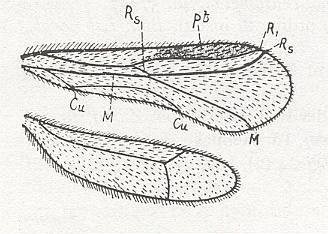
The wing usually is partitioned into two parts, functionally and morphologically very different -- the wing-blade and the wing-base. The wing-blade is, as to its size, usually uncomparably larger than the wing-base. In fact, the large majority of investigations of the insect-wing did concern precisely the description of the features of its blade. This large part of the wing is the chief working part of it as a flight-organ realizing wing-beat. At the down-stroke of the wing use is made of the force of air-resistance, which in this way creates a support for progressive movement -- flight. The structure of the wing-blade really is diverse in different groups of insects. Also diverse is the degree of its differentiation. See next Figure.
|
 (After ROHDENDORF, 1949) |
Figure 1 : Parts of the insect wing. Schematic drawings of the shape of right wings.
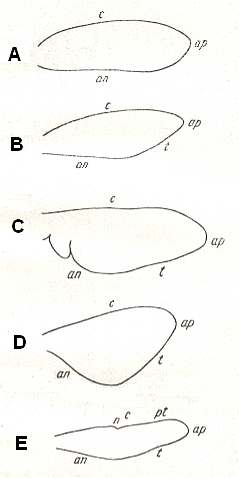
In insects that have a little-perfected flight, and in those [insects] of which the wings did not undergo any advanced specialization, for instance in cockroaches, some Neuroptera, and others, in the wing-blade no well-distinct parts did develop. In these insects one may distinguish merely an anterior or costal edge (costa), a wing-tip (apex), and a posterior or anal edge (analis), Figure 1 - A.
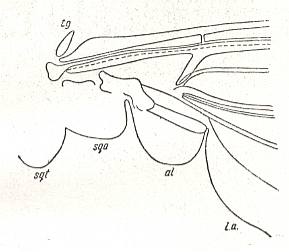
Sometimes the anal lobe may secondarily be reduced [meaning that its absence is not a primitive feature] (in the working out of a special kind of flight [regime]namely paddle-wingedness or kopepterygia). In some groups of insects a sharp distinction occurs between the apical part (wing-tip, apex) and the rest of the wing, which apex separates from the main costal position by the placing of a special thickening of veins or colored spot -- the pterostigma, or [the apex is distinct] by simply a sharp bend of the anterior margin. The proper costal part of the wing-blade usually is one whole most firm structure. Sometimes this part of the wing is subdivided by way of special disconnections or nodes (nodi) into two parts ( Figure 1 - E ).
A special significance the wing-base has in insects that are able to fold their wings back over or along the abdomen when in resting-position. In these cases the structure of the wing-base is significantly complexified by various folds and articulating sclerites. So, in the instance of diptera, we may find in the structure of their basiala special peculiar adaptations, which make possible the folding back in resting-position and expanding when functioning the posterior parts of the wing-blade -- anal lobe, alula, and scales (squamae). These adaptational structures consist in the formation of bristled hooks, chaetaria, and in complexification of the form of the basiala's surface, that is, in development of sharply concave and convex parts. In insects that have broadened wings with a well developed jugal area the latter directly connects with the wing-base.
Types of flight-devices in insects and the courses of their evolution
General remarks
In the previous chapter -- [ Rohdendorf refers here to a chapter (III) -- presenting a review of all Orders of winged insects (Pterygota) from the viewpoint of the flight-devices existing in the representatives of these Orders -- which we here, in the present Part of Website, will only present after the description of the different wing-types (expressing flight-devices) ] -- I gave a concise listing of the features of the evolution of wings and other flight-organs in the insect Orders. In spite of the superficial and cursory consideration, this review nevertheless made possible in general terms to describe the main features and the nature of this [evolutionary] process in the individual groups. With clarity was revealed the completely different degree of differentiation of the wings in the different Orders. Thus, in many Orders we find a relatively uniform development of only one type. Rare deviations, the appearance of any peculiar trait in the structure of the wings in these groups only take place as rare exceptions, so in Blattoidea [cockroaches and allies], Orthoptera-saltatoria [grasshoppers and allies], Heteroptera [bugs], Coleoptera [beetles], Diptera [mosquitoes, flies], and some other Orders. A number of other groups of insects, on the contrary, show diversity in the structure of flight-adaptations, such as Odonata [damselflies, dragonflies], Homoptera [cicadas, aphids, and allies], Hymenoptera [wasps, bees, ants], Lepidoptera [butterflies and moths]. In these Orders the development of several types of flight-devices [i.e. more than one type, where the extra ones are not mere rare exceptions] has taken place, each one of which is widely distributed and differentiated. Such features of the different Orders are undoubtedly the consequence of determined courses of their evolution, and, especially, the consequence of this or another significance of flight in the biology of these insects. Consideration of these questions may, in the framework of the present work, be done only partly and in a certain degree one-sided : The chosen measure of the whole investigation encompassing the whole enormous class of winged insects, forces me to limit myself to merely precisely establish or detect the existing forms of flight-devices, their types and subtypes [ This is also what we, in this website can merely do]. A detailed and careful analysis of the origin and history of each type or subtype, undoubtedly should be the subject of special investigations, devoted to the complete clarification of the courses of evolution of the different groups or Orders of the Class. A broad complex analysis of all the features of each particular group uncovers the causes of the realization of these or those pathways of their evolution and clarifies the significance in this process of the flight-organs much more precisely, than is done in the present investigation.
Aim of the present enquiry is the clarification of the fates of one single function in insects -- flight and the organs realizing it, the wings. Therefore, in a number of cases, where flight-abilities have played only a subordinated role in the evolution of the animals, views about the evolution of those groups have, evidently, a mere preliminary nature.
Schemes of classification of types of flight in insects, proposed by some investigators
The existence of different types of flight in insects was already clear long ago. This was expressed by attempts of classifying the flight of insects, proposed by some authors. Such are, for example, the schemes given by Voss, 1904, 1913. This author, carefully studying the structure of the thorax and wings of the house-cricket, further has used all, existent in his time, data in the literature on other insects and for the first time (1904) presented, in the form of a genealogic tree, a characterization of the wings of insects, dividing them into three large groups : a first, original, hypothetical group, and two actually existing groups of insects with (1) unfolded wings and with reduced longitudinal dorsal muscles (Odonata [damselflies and dragon flies] ), and (2) a group of insects with folded [in the sense of foldable] wings and well developed longitudial muscles (all remaining insects). Further (1913) Voss specifies his scheme, dividing the insects among three types of wings :
I. Straight-winged type (= "Orthopterentyp") -- the lowest type, of which the even development of directly working [i.e. direct transmission of force] and indirectly working muscles, and a more or less unification (functionally and anatomically) of the wings of both pairs, is characteristic. Here do belong, as model-groups the following Orders :
II. Dragonfly type (= "Odonatentyp"), of which a great increase of the number of veins, independently functioning fore and hind pair of wings, and the exclusive development of only direct muscles, is characteristic.
Hereto only the Order Odonata belongs (damselflies and dragonflies).
III. Hymenopterous type (= "Hymenopterentyp")-- the highest type, of which the complete kinematic unity of both pairs of wings, where the hind pair is often completely reduced, and the reduction of direct muscles, is characteristic. Here do belong as example-groups three Orders :
I [Rohdendorf] don't find it necessary to present a careful analysis and critique of Voss' schemes : With the present level of our knowledge such a general very little-detailed scheme cannot of course be satisfactory. The chief insufficiency of this classification consists in the non-evaluation of the existing multiformity of wings in the chief most differentiated insect Orders. In investigating changes of the flight-function and the flight-organs, the wings, one cannot consider [things in] individual Orders as a whole [i.e. assuming that each Order is homogeneous in this respect], as did Voss. On the other hand, mechanical specialization of the wings is not at all accounted for by Voss. As a result, his first "straight-winged" type is in fact a mixture of the most different, little similar to each other, forms of wings. The positive significance of the scheme of Voss consists in correctly distinguishing the "hymenopterous" type, indeed corresponding to the "highest" type of flight-adaptations, by us called dipterygia or two-wingedness.
Attempts do exist of a classification of insect-flight based on the nature of the movements of the insects' wings in flight. Such is the scheme of Pütter, 1912), in which the author distinguishes three groups.
In concluding it is appropriate here to refer to the attempt of classification of flight-features of insects done by Schwanwitch, 1948, in his investigations into the evolution of Pterygota [winged insects]. Fully correctly holding the development of flight to be one of the most principal leading factors in the evolution of insects, Schwanwitch, however, limits himself to only an outline of an extremely artificial classificatory scheme, based on the structure of only the muscular apparatus of the thorax. Not discussing the essence of the constructions of the author, we must note, that -- the separation of the Odonata (damselflies and dragonflies) into the special group of "direct-muscular" insects (= Orthomyaria) from the remaining, the "cross-muscular" insects (= Chiastomyaria), and the division of the latter into (1) "anteromotor" insects (= Anteromotoria"), in which are placed the mayflies, Hemiptera, Homoptera, psocids, thripses, sucking lice, Hymenoptera and all mecopteroid Orders [Mecoptera, (scorpionflies), Lepidoptera, Trichoptera (caddis flies), and Diptera], and (2) "posteromotor" insects (= Posteromotoria), in which are placed the beetles, the Strepsiptera, and the straight-winged insects (Orthoptera) in the broader sense (grasshoppers, locusts, crickets, etc), -- essentially repeat the known scheme of Voss, and only little improving on it. It is regrettable that not a single attempt was made to functionally and purely biologically clarify the significance of the phenomena "antero-" and "postero-motoria", as also that the scheme did not take into account the existing multiformity within the individual Orders, not to say anything about the complete absence of an analysis of the features of the principal organ of flight -- the very wing itself.
Basic premises of the establishment of a precise classification of the types of flight-devices and flight in insects
In composing a list of the different types of flight-organs, attempting to classify the existing changes of the structure of body and wings, we must in all clarity determine precisely those features which characterize this or that type. It is evident that the type must be characterized twofold : on the one hand, based on its functional features, and, on the other hand, based on the morphology of the entire flight-apparatus of the insect body. We take then, consequently, about the following scheme expressing the nature of the type [to be defined].
Functional features :
Biological significance of the changes of the wings and of the evolutionary course of the flight-devices
For a precise determination of the significance of the changes of an organ (in our case of wings) in the evolution of a group, it is necessary to understand their role in the biology of the animal. Only with complete knowlege of the functional significance of the change, say of the venation of the wings, we may clarify the evolutionary path of the given organ. [Rohdendorf here assumes that every morphological feature whatsoever of an organism has functional significance, i.e. it always is connected with the ability of the animal to execute a certain function or performance (locomotion, feeding, sensing, reproducing, etc.). For an organ this is clear enough. We must, however, seriously take into account the possibility that many morphological features, including parts of organs, may not at all be connected with one or another (partial) function of the animal body. They may simply be mere by-products of the evolutionary formation of other structures in the body that do have a connection with some function. Such "other structure" may also be the organ-as-a-whole of which some features or parts may be such by-products. Especially some details of the wing-venation may be functionless and be mere by-products of the development of the overall type of the wing or of its venation. Nevertheless, it remains a good point of departure to assume that every species-constant morphological feature in an organism has some functional significance, whether we have detected it or not.]. Moreover, a correct functional evaluation of observed features makes possible to solve the question of primary or secondary origin. Thus, for instance, a greater or lesser mechanical specialization of the wing-venation with simultaneous absence of such a [aerodynamic mechanical] specialization in the shape of the wing makes possible to hold the secondary nature of the loss of flight-ability (for instance in the group Gryllacridoidea from Orthoptera). Absence of specialization in the wing-venation in the shielding forewings of cockroaches, on the other hand, points to their primary origination [i.e. the wings became shield structures already before they would have been specialized flight-organs]. Such fruitful ways of analysis was indicated by A. V. Martynov, holding that the wings of cockroaches are primitive [instead of derived].
The basic factors, having determined the realization of this or that course in the evolution of wings, were on the one hand, the demand to improve on the quality of flight (its speed, governability, lifting power), and on the other hand, the necessity to guarantee shielding adaptations, making possible to preserve the wholness of the delicate wing organs during inhabiting or visiting by the insect of whatever concealed stations [habitats] -- soil, vegetable remains, tissues of living plants, etc.
The interaction of these opposed factors has determined, in the context of a certain biology of the insect, the realization of this or that course of evolution of the wings.
Up to now, to us unknown are the very first, original, forms of the structure of the wings, i.e. representing transitions from not-projecting sideways notal sclerites or from not-articulate paranotal outgrowths, devoid of venation, to moving blades -- wings.
About these original forms of wings [to have existed] we may judge only indirectly. Evidently, such appendages had to be short and broad, their venation did not have to carry mechanical specialization, being mechanically indifferent, dispersed across the blade of the wing symmetrically with respect to fore and hind margin, evenly strengthening its entire surface. Greatest approximation to such a structure we find in the example of the prothoracic wings of certian paleozoic insects [i.e. short wing-like structures attached on the first thoracic segment -- while normally the wings of insects are attached on the second and third thoracic segments]. See next Figure.
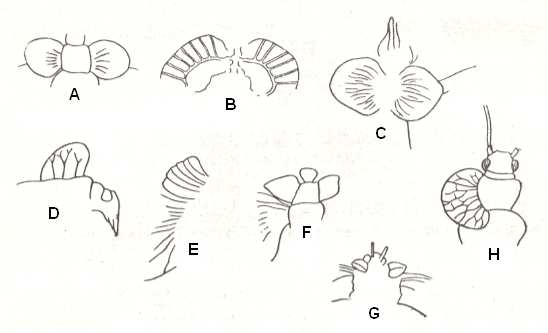
Figure 4 : Prothoracic wings of fossil insects.
A - Stenodictya lobata Brong., Palaeodictyoptera, Dictyoneuridae. B - Stilbocrocis heeri Gold., Palaeodictyoptera, Dictyoneuridae. C - Lithomantis carbonaria Wood., Palaeodictyoptera, Lithomantidae. D - Lycocercus goldenbergi Brong., Palaeodictyoptera, Lycocercidae. E - Homoioptera woodwardi Brong., Palaeodictyoptera, Homoiopteridae. F - Homoeophlebia gigantea Agnus, Palaeodictyoptera, Homoiopteridae. G - Homaloneurina bonnieri Brong., Palaeodictyoptera, Spilapteridae. H - Lemmatophora typa Sell., Protoperlaria, Lemmatophoridae.
Drawings A - G after Handlirsch, H after Tillyard.
(In ROHDENDORF, 1949)
Another example do represent the middle-thoracic wings of many cockroaches, in which, however, may be found already a number of determined traits of specialization, it is true, mainly of a shielding nature.
Figure 5 : Hypothetical ancestors of winged insects.
(After RASNITSYN, 1976, In ROHDENDORF and RASNITSYN, 1980)
The consequent fate of the wings of insects was determined by the working-out of shielding adaptations, which first of all appeared in the form of the acquisition by the wings of the ability to guide them backwards when at rest, to fold them along the body's back [later followed by hardening of the forewings]. This adaptation was realized in one group only, namely in the Neoptera. In the other part of the carboniferous insects the wings did not acquire the ability to fold, and remained spread-out laterally -- this is the group Palaeoptera [today by far the most insects belong to the Neoptera (such as grasshoppers, beetles, butterflies, wasps, flies, etc.]. The working-out in the group Neoptera of shielding adaptations enforced, undoubtedly, an inhibiting effect upon improving the flight-qualities of the wings. As a result of this, precisely among Neoptera we find examples of most primitive, mechanically little-specialized, wings. Such are the wing-types blattopterygia and neuropterygia. Both these types are, what will become clear from what comes later, original [i.e. evolutionary starting points] to the large majority of the other forms of wings.
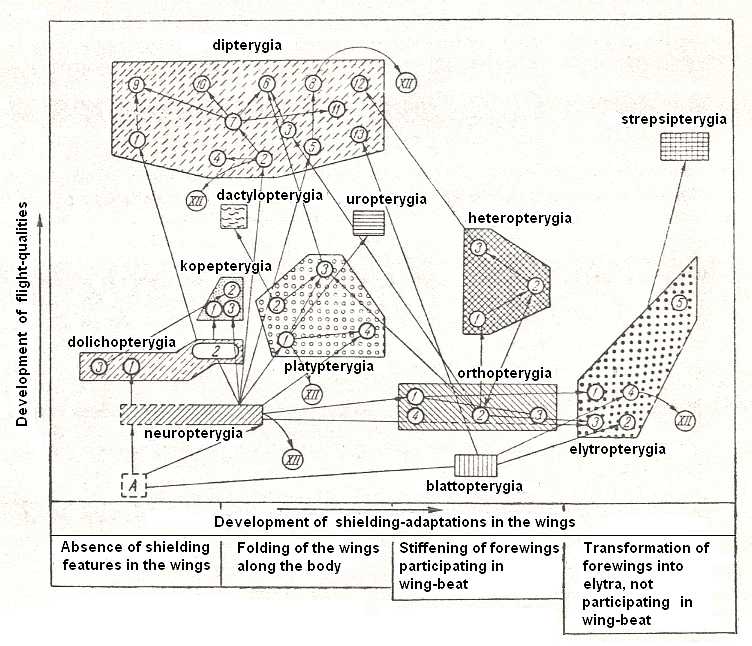
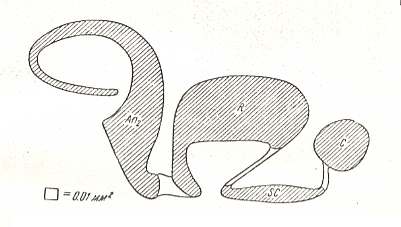
Figure 6 : Scheme illustrating the courses of evolution and interrelationships of the types of flight-features in insects. Types are drawn as polygons placed according to the two axes of the scheme (i.e. these axes serve as coordinate-axes]. The horizontal axis measures the improvement of shielding qualities. The vertical axis measures the improvement of flight-qualities. Arrowed lines indicate courses of evolution of types and their subtypes. Circles with numerals, placed into the background of the polygons, indicate individual subtypes. The square, drawn with interrupted lines in the lower-left corner, with the letter A, signifies the original structure of the first wings -- the paranotals.
The type ptilopterygia (feather-wingedness), as a result of its regressive nature, could not be placed in the scheme with the assumed range of coordinates (it would have been placed down-left outside the field of the scheme] and is everywhere marked by curved lines with an arrow, pointing to a circle with the numeral XII.
(After ROHDENDORF, 1949)
[For the subtypes, indicated by circles with numerals, see the next documents dealing with the types and subtypes of flight-devices in insects]
In the documents to follow we will give the descriptions of the various types and subtypes of flight-devices in insects, following ROHDENDORF, 1949 (with comments and additions). We will start with n e u r o p t e r y g i a, the most original type known, in the next document.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of the evolution of insects, Part II, Neuropterygia.