
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Most expressed example of the type :

Figure 1 : Gomphus vulgatissimus. Family Gomphidae, Suborder Anisoptera, Order Odonata (Dragonflies). 45-50 mm. Wing expanse 60-70 mm. Recent. (After SEVERA, in Thieme's insektengids voor West- en Midden-Europa, 1977.)
Description of the type
Morphological features
A fairly neatly expressed homonomy of the wing-pairs is preserved. Homonomy of middle and hind thoracic segements also well expressed. The same is the case in the muscular apparatus, indicating a rigorously equated homonomous structure. The wings are distinctly elongate, with a straight anterior margin and well-expressed costalization. They are devoid of folds. The venation is very rich, with a large number of, not only longitudinal, but also cross-veins. Sometimes cross-veins are absent, and in their place an archedictyon is found [dense venational mesh work between the longitudinal veins]. Body of insect elongate, with a broadened head and thorax and a narrowed long abdomen. Size of the insects medium or large. Wings usually longer than abdomen, and about equal to the body. Body, although as a rule significantly elongate (such as in Aeschna), in a number of cases a little more stocky (as in Libellula and also in the fossil Palaeodictyoptera).
Functional features
Both pairs of wings take part in the flying movements in their role of supporting the body and creating traction. In this type we find synchronic movement in wing-beat as well as complete autonomy in both pairs. Features of flight are known only of some most specialized representatives of the type -- dragonflies (Order Odonata). Other, more primitive forms of the type are almost unstudied. Speed of flight may reach fairly high figures -- up to 10 m/sec in the dragonfly Libellula, -- normally much lower, about 4 m/sec. Wing-beat frequency is determined to be equal to 22 beats per second in the dragonfly Aeschna, and 28 in Libellula. Governability in flight is high, but not studied in more detail. Load exerted on a unit of wing-surface area varies between widely separated values -- from 0.0090 gr/cm2 in the male damselfly Agria virgo L. up to 0.0450 gr/cm2 in the dragonfly Libellula depressa.
The biological significance of the flight-function in the representatives of this type is very high indeed. These insects spend the greater part of their active life in flight. Most characteristic this is of dragonflies, being active predators, catching prey and mate in flight, performing long-lasting and long-distance flights.
Differentiations, connections, and representatives of the type.
Long-wingedness, or dolichopterygia, is one of the most ancient types of specialization of the wings and originated as early as in upper Carboniferous times. The most ancient carboniferous insects known to us already possessed fully expressed features of dolichopterygia. Therefore we may assess things concerning the ways of formation of this type only on the basis of comparative morphology. In addition, in unravelling this problem, of great help is the observed genesis of other representatives of the type, the neuropterous Myrmeleonidae [ant-lions], having clearly originated from the neuropterygia of their nearest precursors. This legitimates us to assume that also the long-wingedness of the most ancient Palaeodictyoptera (See for example Figure 2 ) was formed out of the neuropterygia of their ancestors unknown to us. The singleness of the source of the formation of long-wingedness determined the relatively uniformity of this type [i.e. its containing only a few subtypes]. Long-wingedness turned out to be the simplest way to improve the aerodynamic qualities of the wings, being realized as a result of the increase of their surface area and their elongation without having to evolutionarily work out any auxiliary adaptations. The known instances of long-wingedness may be divided into three groups.
1. Archidolichopterygia
The most ancient instances of long-wingedness (dolichopterygia) which are found in carboniferous insects are characterized by the weak development of the system of cross-veins, not yet having been formed out of the original meshwork (archedictyon), with the presence of already fully formed longitudinal veins in the elongated costalized wings, and [these most ancient instances together] make up the subtype of archidolichopterygia or, equivalently, ancient long-wingedness. The body of the representatives of this subtype is elongate, but not showing a distinct narrowing. Instances of archidolichopterygia may be found in some primitive families of the Order Palaeodictyoptera (for instance Dictyoneuridae). See next Figures.
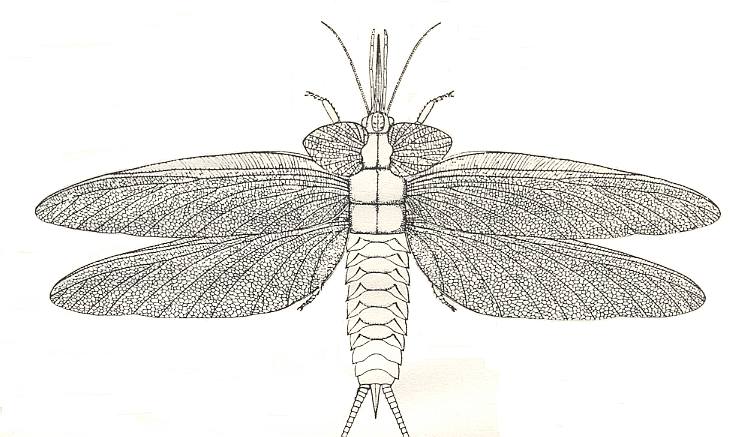
Figure 2 : Reconstruction of Stenodictya, Family Dictyoneuridae, Order Palaeodictyoptera, based on specimens in the Institut de Paléontologie, Paris, France. Upper Carboniferous of France. (After KUKALOVÁ, 1970.)
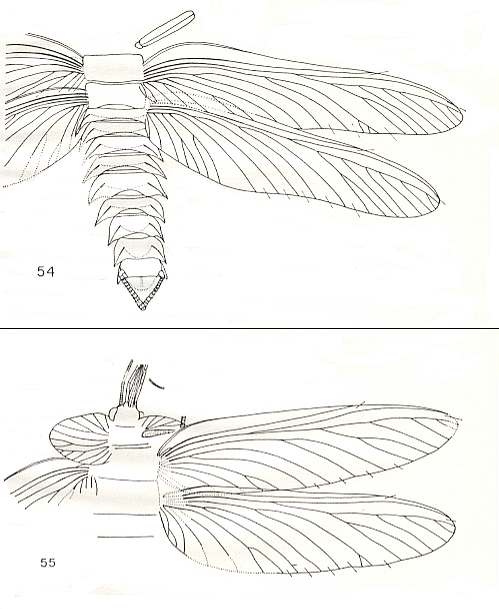
Figure 3 :
Top image : Stenodictya spinosa Brongniart, Family Dictyoneuridae, Order Palaeodictyoptera. Specimen 22-2. Upper Carboniferous of France. Archedictyon [see previous Figure] omitted in the drawing. Forewing length 61 mm, width 13 mm. Hindwing length 61 mm, width 15 mm.
Bottom image : Stenodictya laurentiauxi Kuk. Upper Carboniferous of France. Archedictyon omitted in the drawing. Forewing length 64 mm, width 17 mm. Hindwing length 64 mm, width 18 mm.
(After KUKALOVÁ, 1970.)
This subtype is the direct source of the next one -- true dolichopterygia or eudolichopterygia.
2. Eudolichopterygia
The great majority of the representatives of dolichopterygia is characterized by a fully developed cross-vein venation, having already lost the traits of the original meshwork (archedictyon), and by a long thin abdomen. This is true long-wingedness, or eudolichopterygia. To this subtype do belong numerous families of the same ancient Palaeodictyoptera, for instance Homoiopteridae, Spilapteridae, and others) :
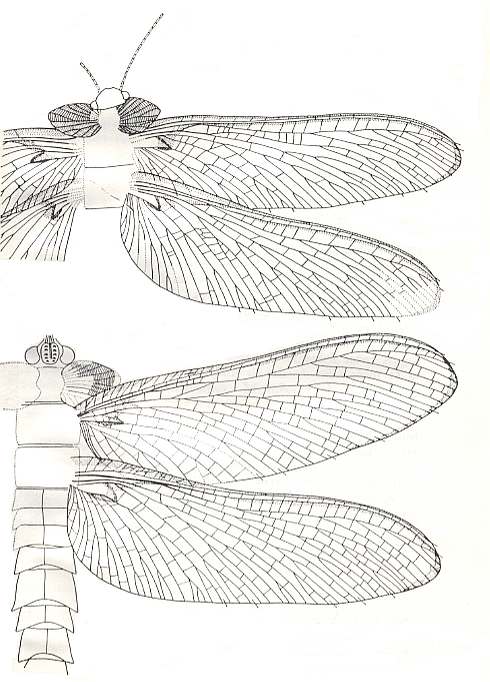
Figure 4 : Homaloneura bonnieri Brongniart. Family Spilapteridae, Order Palaeodictyoptera. Specimen 17-17 and 17-18. Upper Carboniferous of France. Length of forewings 43-40.5 mm. Width of forewings 12-11 mm. Length of hindwings 43-41 mm. Width of hindwings 15-14.5 mm. Kukalova further mentions (p. 174) "Wing membrane very thin", and thus indicating that the wings are not coriaceous. (After KUKALOVÁ, 1969.)
Further, the paleozoic may-flies, Triplosobidae and Protereismatidae :
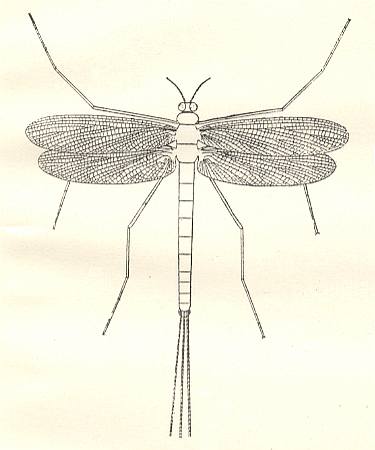
Figure 5 : The permian may-fly Protereisma permianum Sell. Family Protereismatidae, Order Ephemeroptera [= Plectoptera = Agnatha]. Reconstruction by CARPENTER. Lower Permian of Kansas (USA).
(After CARPENTER, in MARTYNOV, 1938.)
and, finally, the most ancient dragonflies Protodonata, Meganisoptera. Probably, representatives of the subtype are also some recent damselflies Agrionidae. And in this subtype we must also include representatives of a totally different group of winged insects, that is, of the Order Neuroptera, namely Myiodactylidae and Myrmeleontidae [and allies] :
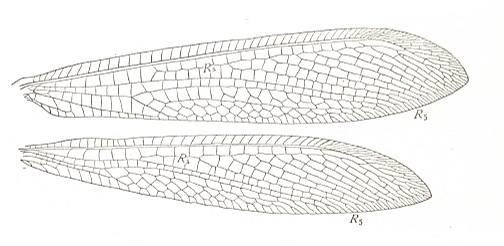
Figure 6 : Wings of Myrmeleon, Family Myrmeleonidae, Order Neuroptera-Planipennia. A species from Katihar, former British India. Recent. (After COMSTOCK, 1918.)
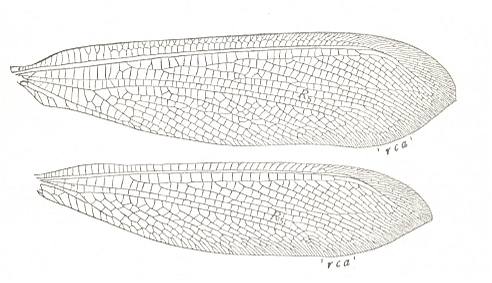
Figure 7 : Wings of Acanthaclisis, Family [prob.] Myrmeleonidae, Order Neuroptera-Planipennia. A species from the Seychelles Islands. Recent. (After COMSTOCK, 1918.)
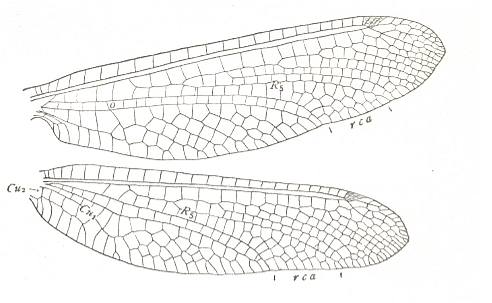
Figure 8 : Wings of Ululodes hyalina. Family Ascalaphidae, Order Neuroptera-Planipennia. Recent. The Ascalaphidae are one of the few planipennian families of which the representatives are able flyers. They are not nocturnal, but fly at daytime.
(After COMSTOCK, 1918.)
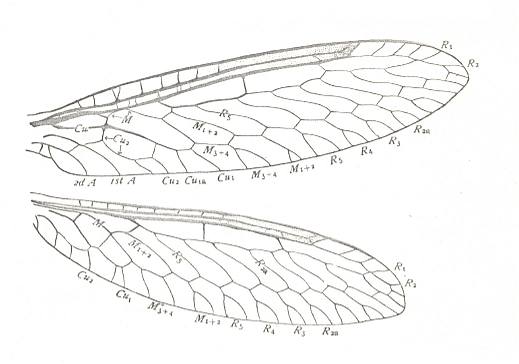
Figure 8a : Wings of Climaciella brunnea, Family Mantispidae, Order Neuroptera-Planipennia. Recent.
(After COMSTOCK, 1918.)
Wing pairs homonomous, hindwing only a little shorter than forewing. Venation and shape similar in both wings.
Representatives of some extinct orthopteroids -- Caloneuridea, possessing homonomous wings, undoubtedly also must be reckoned here to belong to the present subtype (eudolichopterygia). And maybe also the termites (Order Isoptera) belong to this subtype :

Figure 9 : A winged termite ( Archotermopsis), male. Order Isoptera. Recent.
(After RICHARDS and DAVIES in Imms' General Textbook of Entomology, 1977.)
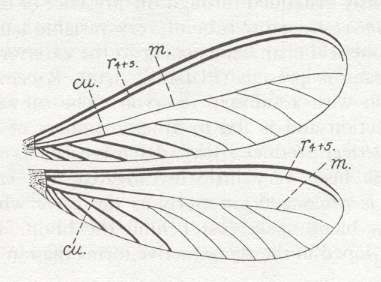
Figure 10 : Fore- and hindwings of Eutermes sp., Order Isoptera. Recent.
(After RICHARDS and DAVIES in Imms' General Textbook of Entomology, 1977.)
Not all termites have wings of the dolichopterygian type, for example Mastotermes darwiniensis which belongs to the subtype neur-orthopterygia of the type orthopterygia (straight-wingedness).
Yet another (albeit a remarkably small) wing (in addition to the ones figured higher above) of the Order Palaeodictyoptera is found (in permian strata) which gives a very clear example of eudolichopterygia :
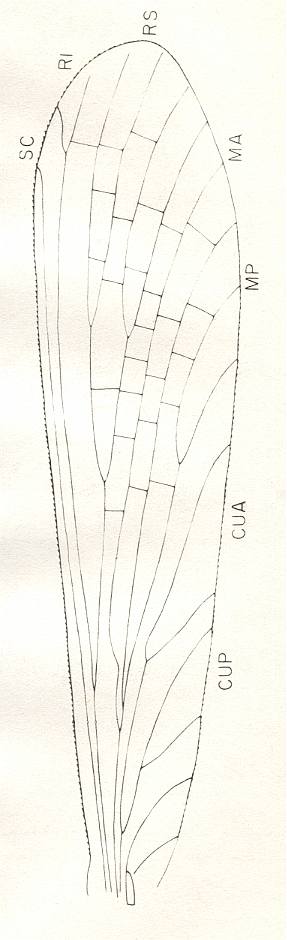
Figure 11 : Forewing of Elmoboria piperi Carp. Family Elmoboriidae, Order Palaeodictyoptera. Lower Permian of Kansas (USA). Length of wing 15 mm. Hindwing and body unknown. Because of the paleodictyopterous nature of this wing, we may safely assume that the hindwing was similar to the forewing (homonomy of wing-pairs) except for, perhaps, having a slightly broader anal area.
(After CARPENTER, 1976)
To the present subtype of dolichopterygia, eudolichopterygia, also may belong the wings of the family Panorpidae (and fossil relatives) of the Order Mecoptera (Scorpionflies) :
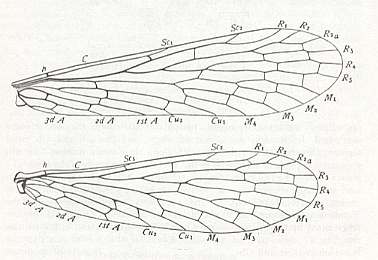
Figure 12 : Wings of Panorpa, Mecoptera, Panorpidae. Recent.
(After COMSTOCK, 1918, in IMMS' General Textbook of Entomology, 1977)
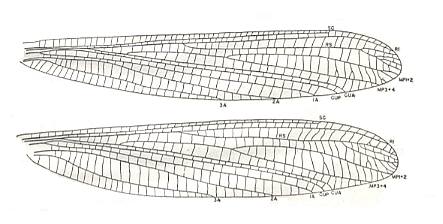
Figure 12a : Paleuthygramma acuta Carp. Family Paleuthygrammidae, Order Caloneurodea.
Top - Forewing. Drawing based chiefly on holotype and specimen no. 4563.
Bottom - Hindwing. Drawing based chiefly on holotype and specimen no. 4554.
Forewing length (holotype) 18 mm, width 3 mm. Hindwing length (holotype) 17 mm, width 4 mm.
(After CARPENTER, 1943)
The eudolichopterygia of some extinct orthopteroids was probably the source of the simplest forms of straight-wingedness ( orthopterygia ) by way of acquiring shielding qualities -- stiffening of the anterior wing-pair [and having the jugo-anal area of the hindwings expanded]. Another derivative of this subtype is paddle-wingedness (kopepterygia) having originated as a result of narrowing and stiffening of the wing-base.
3. Metadolichopterygia
Finally, the last, most specialized subtype of long-wingedness originated as a result of the working out by the wings of various mechanical adaptations increasing their elasticity while retaining a high degree of firmness involving the formation of special interruptions and nodes [nodi], and of the marked narrowing of the distal part of the body -- the abdomen. This kind of perfection of the wings took place together with a transformation of the muscular apparatus as a result of the development of pleural musculature at the expense of the reduction of other groups of muscles. The [in this way] achieved amazing distinction of the dragonflies perhaps justifies this subtype to be a type of its own. Perfect long-wingedness or, equivalently, metadolichopterygia, is expressed in representatives of the Order Odonata, namely in the Suborders Permanisoptera (extinct), Protanisoptera (extinct), Anisozygoptera, and especially in Anisoptera, see Figure 1, and next Figures.
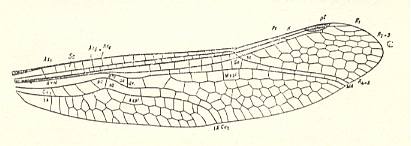
Figure 13 : Wing of Polytaxineura stanleyi Till., Order Odonata-Anisoptera. Upper Permian (Warner's Bay) Australia.
(After TILLYARD, in MARTYNOV, 1938)
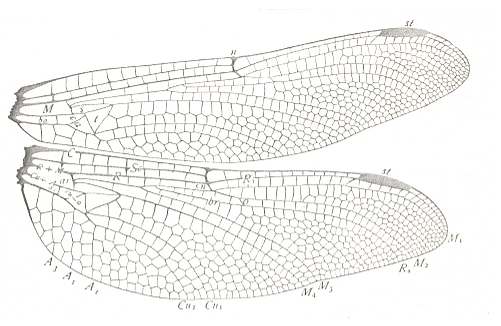
Figure 14 : Wings [i.e. fore- and hindwing] of Gomphus descriptus. Family Gomphidae, Order Odonata-Anisoptera. Recent.
Labelling and interpretation of veins and cells according to Comstock.
(After COMSTOCK, 1918)
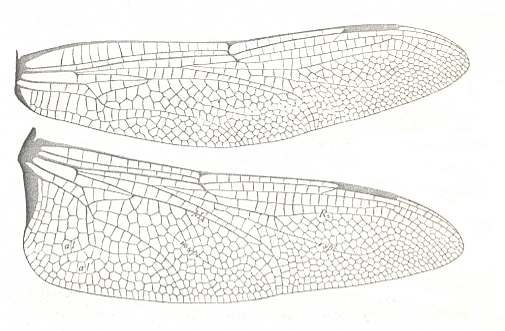
Figure 15 : Wings of Anax junius. Family Aeschnidae, Order Odonata-Anisoptera. Recent.
Labelling and interpretation of veins and cells according to Comstock.
(After COMSTOCK, 1918)
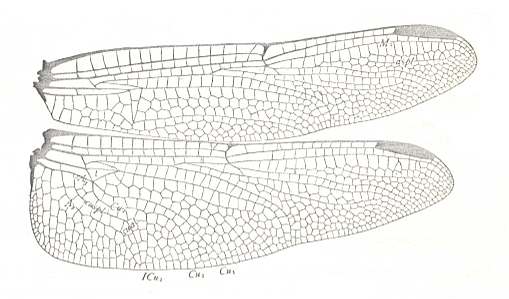
Figure 16 : Wings of Orthemis ferruginea. Order Odonata-Anisoptera. Recent.
Labelling and interpretation of veins and cells according to Comstock.
(After COMSTOCK, 1918)
Metadolichopterygia, undoubtedly, is the in the highest degree mechanically worked-out expression of long-wingedness, which is proven by its recent distribution, history, and its high flight-qualities (speed). This subtype, as well as eu-dolichopterygia, shows a connection with another type, namely paddle-wingedness, having been, apparently, one of the sources of its formation.
The relationships in the present type are depicted in the next diagram.
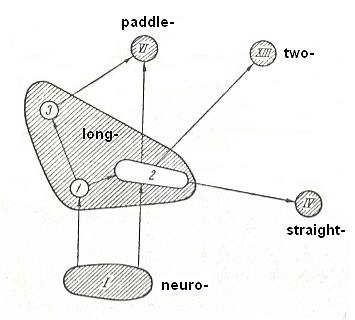
Figure 17 : Relationships with other types and structure of long-wingedness (dolichopterygia). (After ROHDENDORF, 1949)
With all this we have concluded the exposition of long-wingedness. In the next document we will expound the wing-type paddle-wingedness (kopepterygia).
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of the types of flight-devices in insects, Part VII, Kopepterygia.