e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Continuing the theory of polydiagrams (previous document)
In our noëtic theory of evolution we had developed the method of polydiagrams. In such a diagram we (symbolically) show the noëtic construction, in the Implicate Order, of strategy-contents, ready to be projected into the Explicate Order as soon as the appropriate existential conditions are present in that Order. The set of strategy-contents considered in the diagram corresponds to a group of related organismic species. And every ascending series of noëtic production-stages is in fact a bundle of noëtic pathways each one of which represents the noëtic construction of one or another (functional) character. And each such a noëtic pathway of successive noëtic states of a given character is approximately reflected in the Explicate Order by a series of formally related states of the corresponding material character. Such a series of formally related states in the Explicate Order, i.e. a series of character-states that can more or less be formally derived from each other, we had called a "derivational line" and placed it horizontally up into the polydiagram. And as explained earlier, for different characters these derivational lines may not coincide as to the derivational order in them. In the previous document we made such a diagram for two symbolically represented characters in insects, viz., wing-venation and mouthpart-structure. Of course, ideally, all (functional) characters present in the species of the group figuring in the polydiagram should be considered in it, but heuristically we may limit ourselves to one character only. And when dealing with winged insects, their wing-venation is most appropriate to figure as a character in polydiagrams to show at least one aspect of the noëtic construction of strategy-contents.
In the previous document we had made such a one-character polydiagram figuring six, allegedly, closely related organismic species A, B, C, D, E, and F as to the noëtic construction of a single character in them. They were supposed to be arranged such that the various existing states of that character (present in the species) could be, albeit approximately, formally derived one from the other. And so the species A, B, C, D, E, F together form a "derivational line", and as such these species were placed in the Explicate-Order-part of the diagram. Directly below them, now in the Implicate-Order-part of the diagram, A, B, C, D, E, and F were placed as representing the corresponding immaterial strategy-contents. Each such a strategy-content (in fact a noëtic description of the strategy) is the result of, in fact, a bundle of noëtic construction pathways, while we here in the diagram did consider only one of them, i.e. a pathway representing the noëtic construction of the end-state of that one character in each strategy-content A, B, C, D, E, and F, as (these pathways are) indicated by the blue vertical arrows. Letting A, B, C, D, E, and F represent closely related species of winged insects, and this single character be represented by the wing-venation, we had (in the previous document) the following polydiagram :
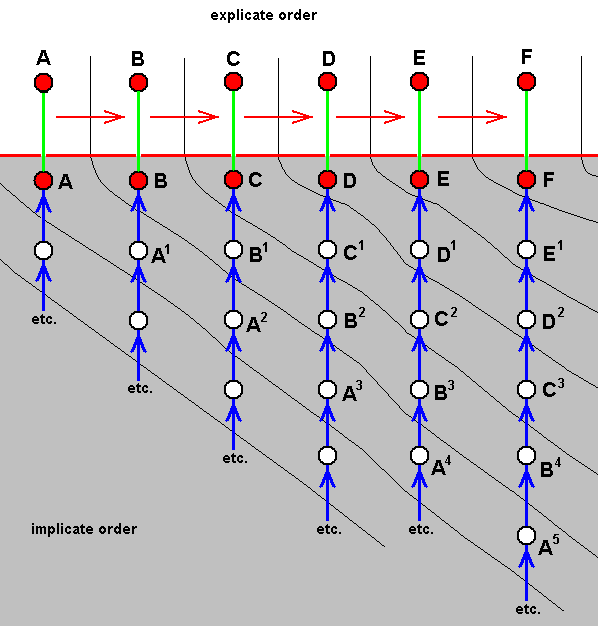
Figure 1 :
This type of diagram (i.e. polydiagrams) deals with specialization-crossing as it is so often encountered in organismic species. As a result of this phenomenon organismic species cannot properly be derived from each other which gives good reason that they did not materially evolve from each other at all. The diagram shows that things go via the Implicate Order, i.e. through the mediation of immaterial not-yet strategy structures that do not, and cannot, exist in the Explicate Order. And this here means, not even materially so (while other immaterial structures, the true strategy-contents, can so exist).
So in this diagram it is expressed, for example, that strategy-content E, insofar as wing-venation is concerned, cannot be considered to have noëtically been produced from strategy-content D, because the wing-venation of (the material) species E, although roughly derivable from that of species D, cannot (as is so often the case in real insects!) strictly be derived from that of species D. So the wing-venation as it is present in strategy-content E (in the Implicate Order) has been noëtically constructed, not from that of strategy-content D, but from that of not-yet strategy-content D1 which is similar to, but not identical with, that of strategy-content D.
The same can be said of strategy-content B not constructed from A but from A1 which merely looks like A, of strategy-content C not constructed from B but from B1, etc. And this means that in some way we may say that the wing-venation of species F (as it resides in the Explicate Order) at least partly "comes from" that of species E (also as such residing in the Explicate Order), but through (mediation of) the Implicate Order, i.e. through the venation as noëtically described in the not-yet strategy-content E1 (which itself does not exist in the Explicate Order). In the same way we may say that, as to the wing-venation, species E partly comes from species D through D1, species D from C through C1, species C from B through B1, and B from A through A1 (all as to their wing-venation only). So the species forming the derivational line in the diagram as to their wing-venation, are each for themselves not merely connected with their respective noëtic production-stages in the Implicate Order, but also, in a way with each other. But, as a rule, they are differently so connected insofar as these connections result from, say, mouthpart-structure only. This, and some of its implications, was already explained in the previous document, but here we need to look still more closely to it. Knowing that things hold for all cases just mentioned, let us take, as an example, the case of the venation of species D coming in some way from that of species C through that of the not-yet strategy C1. And, as before let us dipict things as a part of the original polydiagram :
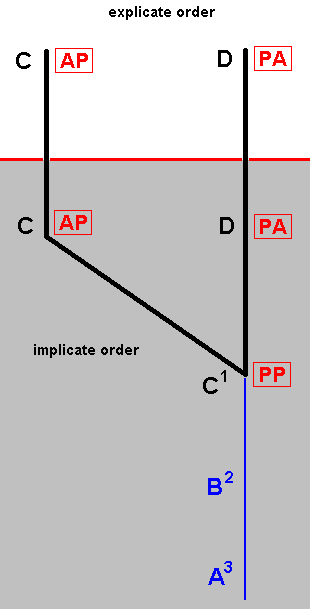
|
Figure 2 : Relationship of species D with species C, in the polydiagram in Figure 1, via the Implicate Order. Such a relationship between related species is a consequence of the general structure of a polydiagram : As can be read off from the diagram at the left (and generally from that of Figure 1), every ascending series of noëtic strategy-stages traverses a number of "influencial domains", or "areas of influence" in the Implicate Order (oblique strips in the diagram), where "influence" is always directed from upper-left to lower right only : Take for example the noëtic construction of the (noëtic description of the) wing-venation in strategy-content D, while considering only the segment A3==>==>D. While "going up", the next construction-stage upon A3 enters the "B-area" resulting in this next stage to become such that it looks like or "assimulates" (as to wing-venation) the noëtic stage B1 (of C), while the next construction-stage (of D) enters the "C-area" resulting this stage now to "assimilate" the strategy-content C (that is, now looking, as to its wing-venation, like C). Finally the next and last state is the wing-venation in strategy-content D, which (strategy-content) will, upon projection, appear as an insect-species in the Explicate Order. It must be noted that all this, i.e. the oblique 'areas' of influence in the Implicate Order, is not something that 'unexpectedly' happens to f o l l o w from our noëtic theory. The idea of the structure of polydiagrams is entirely dictated by the actually observed qualitative relationships, often involving specialization-crossings, in living and fossil organisms. Any proposed mechanism of evolution (no matter what this term is supposed to mean) must result in the organisms as we actually encounter them in Nature. Our polydiagrams describe these observed relationships in terms of the Implicate and Explicate Orders, while in other theories these same relationships are described in different terms. |
With all this in mind, let us now go back to the relationships between different organismic species, relationships via the Implicate Order, and especially to our fictitious example of such a relationship, as indicated in the above diagram, between the (symbolic) insect species C and D, in fact (only) between the wing-venation possessed by these species, and let us then show how we, in a polydiagram, deal with specialization-crossing.
As to the noëtic ascent leading to strategy-content D, the transition from B2 to C1 is more or less remarkable (as in fact all 'vertical' transitions in all species (strategy-contents) are). As we can see in Figure 1, above, it has to do with the mentioned oblique "areas of influence". Let us theorize about the nature of that influence. When stage B2 enters the "C-area" the selection of the production-rule (from the set of FSNSS-rules) that is supposed to transform this stage into the next stage, is influenced by (end)stage C (of the line of ascent lying next to it) resulting in the fact that the venation of this next stage becomes similar as to its overall structural pattern to that of C, except for the state of Subcosta in it, which is still not, as it was already in C, in a derived condition with respect to Subcosta in D, but in a still primitive condition, while the Cubitus is, like it is in C, still in its primitive condition, and only in the next transformation it will end up in a derived condition, while the Subcosta remains in its primitive state.
So the nature of connection between species D and C is not one of strict derivation or production, that is, the black line in the diagram going from species C to strategy-content C, and then to not-yet strategy-content C1 and further to strategy-content D, and finally to species D, does not represent a derivation or production along it, because C1 (PP) cannot be derived from C (AP), and, in the other direction, C1 (PP) cannot have been produced from D (PA), but it is a connection nevertheless, i.e. the black line indicates a connection between these two species -- as to their wing-venation --, a connection via the Implicate Order. It is a qualitative connection between the two species as a result of similarity of their respective venational patterns. Let us characterize the nature of this connection as it manifests itself between the 'stations' C, C, C1, D, and D of the black line, and show that not-yet strategy-content C1 is not produced from C, but merely connected in some way with it. It is produced from not-yet strategy-content B2, but this production is here influenced by strategy-content C which lies in the noëtic vicinity of C1.
Let us follow, then, the black connection line (the thread connecting species C with D) :
Qualitative hiatus between species, expressed by polydiagrams.
In the above example (in Figure 1) we had taken six supposedly closely related species that allow their wing-venation, -- albeit approximately and thus ignoring one or more venational specialization-crossings existing in the species but supposedly considered to be less significant ones, -- to be formally derivable from one another.
But there are, of course, many cases in which wing-venations cannot at all be derived from one another. As an example we may note the general venational schemes as they occur in the Order Mecoptera (scorpionflies) on the one hand, and those of the Order Hymenoptera (wasps, bees, ants) on the other. And although there is some evidence that the Order Hymenoptera has been produced from the Order Mecoptera, their respective wing-venational schemes do not match in some derivational way to one another in any way. And indeed, hymenopterous wing-venation does not show morphological affinity with that of any other insect Order at all, even when taking fossils into acount ( This is also the case with Odonatean [dragonfly] venation, with Ephemeropteran [mayfly] venation, and, probably, with Homopteran [cicadas, aphids, and the like] venation). Of course one may press things and simply say that hymenopterous venation is derivable from mecopterous venation, but that is only because the latter is so generalized ('primitive'), meaning that almost any other wing-venation is derivable from it. We cannot, in a detailed and, especially, in a unique way (meaning that it is the only possible way) derive the hymenopteran wing-venation from mecopteran venation. So if we insist that, nevertheless, there exists some connection between Hymenoptera and Mecoptera, we must admit that, as to their respective venational schemes, there exists a wide gap between them, not closed in any way by fossils.
But such a hiatus is nevertheless perfectly expressible in the method of polydiagrams.
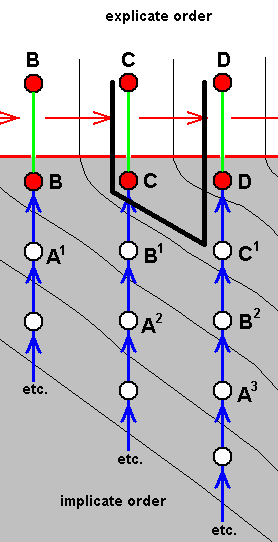
We may, in this, visualize A, B, C, D, E, and F again, but now representing a different case (and representing symbolically species quite different from those of the mentioned example ( Figure 1 ). And now we assume that no species D exists or has ever existed, and that, partly as a result of this, there is a wide venational gap (i.e. a hiatus as to the structure of the venation) between ABC on the one hand, and EF on the other (ABC could, for instance, represent relevant species (fossil and recent) of Mecoptera, while EF might represent relevant (fossil and recent) species of Hymenoptera). So now the venation of species E (and species F for that matter) cannot in any way reasonably (and uniquely) be derived from that of species C (and from species B for that matter). Between them there thus exists a wide venational gap not narrowed by venations of existing or having existed species. And consequently also the venation (as prescribed) in the immaterial strategy-content E cannot have been noëtically produced from that in strategy-content C in the Implicate Order. We may express this in the following polydiagram :
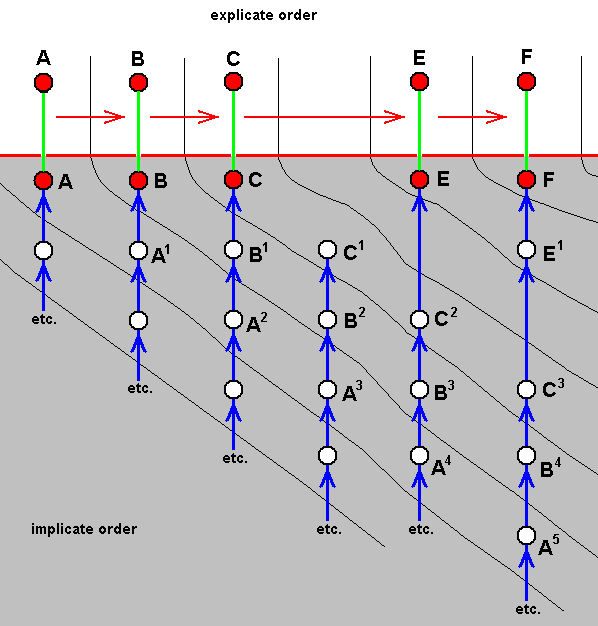
Figure 4 : Polydiagram expressing the hiatus between ABC on the one hand, and EF on the other, as to wing-venation.
Because, as has been explained, there are in the Implicate Order " 'areas' of influence", depicted as oblique strips in the polydiagrams, we may say that precisely the ascending series of strategy-stages, namely the series . . . ==> A3 ==> B2 ==> C1, which does not lead to a final conclusion (i.e. does not lead to a theorem expressing a true strategy-content), mediates in some way between the neighboring ascending series of species C on the one hand, and that of E on the other. We may say that species C connects (in the way described earlier) with species E via the Implicate Order. That is to say, the material species C is connected to the material species E through the immaterial strategy-content C, the immaterial non-strategycontent C1, the immaterial not-yet strategy-content C2, and the immaterial strategy-content E, of which C1 and C2 do not (materially) exist in the Explicate Order. In it the gap between the venational plans of species C and E is represented by the productional jump from C2 to E.
When (symbolically) considering not one but two characters (f.i. wing-venation and mouthpart-structure) in the same (as above) group of insects, we may get the following polydiagram :
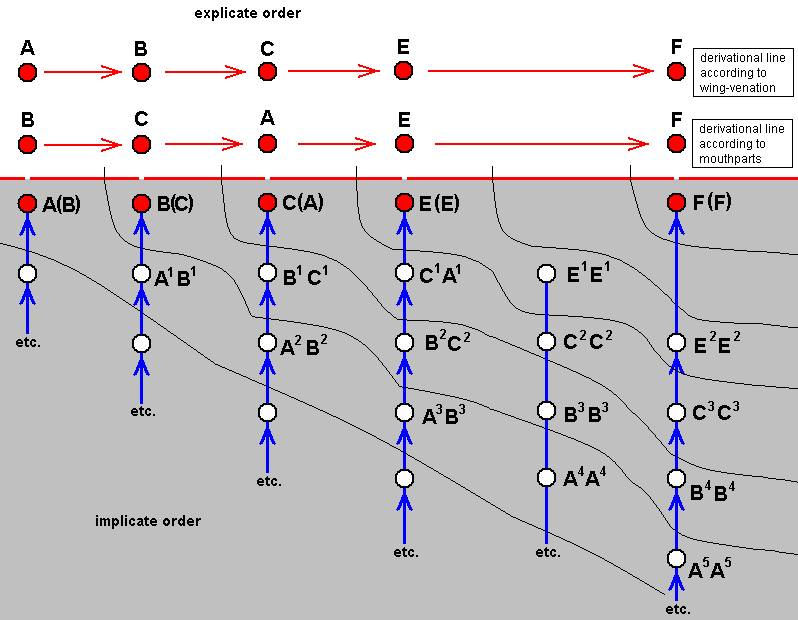
Figure 5 : Polydiagram expressing the hiatus between ABC on the one hand, and EF on the other, as to (symbolically) wing-venation and mouthpart-structure.
The following following "conclusion" of our "noëtic theory of evolution" is a bit complicated. Therefore we begin by giving a summary of it in a more or less schematic way.
First we must realize that in our theory the concept of "evolution" is NOT considered to express a mechanical process of given organismic species genealogically descending from other species and so constituting a "genealogic tree" of descent. Neither is it a construction of organic strategies from other such strategies. So 'evolution' is, in our theory, nothing else than the successive appearance, in the Explicate Order, of organismic species and ecological types, and the extinction of others, as a result of changing ecological conditions. This means that earlier species or types are not necessarily more "primitive" than later ones, and this, of course, makes true derivational directions of forms, species and structures much harder to assess. Earlier species or types are only ecologically different from later ones. Their time of appearance in the Explicate Order is dictated by prevailing ecological conditions only. And it is clear that in all this functional characters play a decisive role. The derivational direction within a sequence of character states can be read off from the degree of functional adaptation to newly prevailing ecological conditions.
Insofar as actual construction ('evolution') is concerned, it is a construction, in each case, of a strategy from non-strategy stages, a construction that takes place, not in the mechanical Explicate Order, but in the Implicate Order. Further, it should be realized that the Implicate Order is timeless and spaceless, in spite of our dynamical description of it. With such a description we only expound aspects of the structure of the Implicate Order. And only in the latter Order there exists a teleologic aspect necessary to create functional structures (here, noëtic descriptions of such structures). This teleologic element consists in the Implicate Order's 'aspiration' (= ontological inclination) to have its immaterial forms materialized and thus having its forms to become functional with respect to their existence and persistence in the Explicate Order. In this noëtic construction of strategies the Explicate Order does play some part, as described. And, methodically, we can get some insight into the noëtic process as a result of the supposed fact that derivational relations existing between structures of different but related organismic species approximately reflect the formal process in the Implicate Order of strategy-construction. For this to investigate we use polydiagrams, as was explained in the previous document.
Now, in the coming conclusion of our noëtic theory of evolution we have realized that in the construction of organic survival strategies the part played in it by the Explicate Order must be greater still than initially assumed. We, therefore, consider the assumed fact of subsequent specialization (and also facts of universalization) of a newly projected organismic species to be a result of its actual experience in its environment. And for this to be true we must conjecture that the initial strategy of a newly projected organismic species is still not yet fully defined or determined. This brings with it the problem of determinacy / indeterminacy of the Explicate Order, i.e. its modal structure. And while inorganic beings (such as atoms, molecules, and crystals) seem to be fully deterministic as to their behavior, organisms do not, and this [not being fully deterministic] is only possible because they are truly holistic entities, i.e. true substances in the metaphysical, i.e. aristotelian sense (see for this Back to Homepage, i.e. First Part of Website ). But while inorganic beings are such substances as well, they have already stabilized their habits (i.e. have already transformed their behavior into true habits) which now constitute these inorganic substances' intrinsic and constant nature, and this in contrast to organisms. Indeed, organisms must further determine their specific nature or strategy, and do this by learning and habit-formation during their existence in the Explicate Order. And only when new behavior happens to be continually repeated in more and more individuals of the species, it can be injected into the Implicate Order, and only then, as a result of noëtic reactions, morphological structures can be adjusted to further sustain, in the Explicate Order, the new behavior. This new behavior and accompanying morphological structures do not, however, result in a new organismic species, but only further delimit its strategy.
And in the section "Habitat and ecological niche" we will find out that the initial indeterminacy of the newly-projected strategy consists in the fact that, in many cases at least, the strategy, as it is constructed in the Implicate Order, is ordered not directly to the species' appropriate ecological niche, but to the species' appropriate habitat, which is the broader environment (-al type) in which it at least can live, but in which also a great many other species can live. The habitat satisfies only the very minimal requirements of such species. And as a result of experience and learning, a given species may specialize within its habitat by selecting certain not yet exploited resources in it with the help of adapting its behavior and thus by more narrowly defining or adjusting its strategy, and so finding at last its proper ecological niche, which is now its ecological station within its habitat. And it is the interaction in all this between the material organismic species in the Explicate Order and the noëtic strategy content in the Implicate Order that guarantees the eventual (but nonetheless temporary) stabilization and delimitation of the strategy.
Injection and projection, and formative causation
In the previous document we considered the 'construction', or, better, formation -- in the Implicate Order -- of organic strategies, and indicated the part therein played by the Explicate Order. Indeed, while stressing that the Explicate Order cannot create or produce true organic strategies all by itself, and thus us having devoted so much attention to the generative processes in the Implicate Order [where "generative processes in the Implicate Order" is just a dynamical way to describe the timeless structure of the Implicate Order. Only as 'seen' from the Explicate Order they are 'processes'], we realize that the part (that) the Explicate Order does play in it should be supposed to be a bit greater still. This is evident from the nature of all functional structures and instincts present in organisms. They explicitly reflect environmental structures and conditions. So we may perhaps theorize that strategy-contents, -- insofar as they have been constructed in the Implicate Order, where this construction was, it must be admitted, already ordered to explicate-order-existential-conditions, and thus [this construction being] already adapted to fill existing ecological niches, -- are still more or less crude and generalized at the time (seen from the Explicate Order) of them being projected for the first time. That is, although such a noëtically constructed strategy is suitable enough to being materialized and able to exist as an organismic species in its ecological niche, merely "able to exist" is still not good enough. It must be able to p e r s i s t as well. This means that the interaction of the new species (i.e. a strategy being projected for the first time) with its environment must be intensified in order to render the strategy complete and perfected, and especially rendering it such that competition with other, related, strategies is avoided. And competition can be avoided by specializing within the ecological niche, or, the other way around, by becoming more universal (i.e. becoming less of a fussy eater).
So in theorizing about evolution, we must, still more so than we had done before, involve existing (or having existed) ecological conditions and the organisms' dealing with them, insofar as this takes part in the formation, and, especially, in the perfection, of organic strategies. And for this to do so, it turns out that certain ideas or elements making up the so-called "hypothesis of formative causation" proposed by SHELDRAKE in 1981, and further elaborated in 1988, may, when properly adapted to the tenets of our noëtic theory (as far as this is fair to do so), contribute to our insight in what way, precisely, the Explicate Order plays a formative role in the creation of strategies. As is perhaps well known, SHELDRAKE proposes the existence of "organizing fields" influencing organisms through "morphic resonance", or being themselves influenced by the organisms also through morphic resonance. This looks very much like our "projection" and "injection", while the "fields" themselves look very much like our "noëtic strategy-contents" (or strategy-strings), and while, finally, the overall behavior of our "materialized strategies", i.e. of the individuals of a given organismic species, has much in common with SHELDRAKE's "stabilized habits". However, although SHELDRAKE's theory may well account for the features of morphogenesis (individual development, especially when embryonic development is involved), it does not account for biological evolution, at least not sufficiently so. Nevertheless many ideas in it are useful to supplement our own noëtic theory of evolution. And one such idea is about learning. In the evolution of life, or we might say, already in the process of increasing the organism's match with its ecological niche, or in its specialization or universalization within that niche, learning, according to SHELDRAKE, plays an important role. And, also according to him, all organisms (all plants and animals) are able to learn, and, in contrast to what is held by conventional genetic theory, do pass on what has been learnt to the next generation. Here we must take 'learning' in the broadest sense and thus not only including behavioral responses but also morphological responses to prevailing environmental conditions.
What now follows is an argument explaining why it is necessary to introduce in the theory of evolution such controversial notions as "learning" and "inheritance of acquired characteristics". It is further considered what precisely the supposed process of making up and perfecting the strategy or habit of an organism means to our conception of the modality of the Explicate Order, i.e. how the modal categories such as (it is) possible, (it is) real, (it is) coincidential and (it is) necessary, and all their negations, do actually relate to each other (for instance, do they imply each other, or exclude each other?). And then the question of the indeterminacy or determinacy of the "real world" (the Explicate Order) naturally comes up.
SHELDRAKE argues that all "laws of nature" merely express more or less stabilized habits of things, i.e. they are the result of development of habit. This implies that, in the Explicate Order at least, there is no strict and fixed determinism. Such a determinism is only, in many cases, approximated to. However, this seems to contradict the modality of the real world (the latter as ontologically distinguished from the "ideal" world of immaterial entities). In the real world (the Explicate Order), if something, say A , actually exists, i.e. is real, it must at the same time be possible. But if A is truly possible, then the complete set of conditions (for it to be possible) must be present at that time. And if only even one member of this set of conditions is missing, then A would be impossible. So when A is indeed possible, the complete set of conditions for A to be possible must be present. But then, A not only exists as a result, but must exist, meaning that A is at the same time necessary (it necesarily exists). So everything that exists (in the Explicate Order), at a certain point in time, does so necessarily. It cannot not exist at that point in time. And this means that the real world (the Explicate Order) is fully deterministic. This analysis can be found in much more detail in the book " Möglichkeit und Wirklichkeit " written by Nicolai Hartmann already in 1938.
If qualitatively completely identical individual entities always, without exception, behave in precisely the same way in everytime identical ambient conditions (if "identical ambient conditions" are at all possible), then their behavior can be described as taking place according to a definite and constant natural l a w. As such, such a law is purely theoretical, because the individual cases are either separated from each other spatially or temporally, or both, implying that ambient conditions will differ. And there is good reason to believe that the overall collocation of matter and energy in the Universe as to their distribution, is constantly changing from moment to moment. So in actual practice the law (any natural law whatsoever) will not hold exactly, but this "not holding exactly" is merely a per accidens phenomenon. The law is, as a strict law, immanent in all the things for whose behavior it is a law. This is the assumption of strict determinism (at least as regards things above the quantum level). That it is the actual course of things to act in merely an approximated version, is extrinsic to it [i.e. to determinism itself] (indeed, because of the changing collocation of matter and energy, it does not get the chance to fully express itself).
But although this assumption of determinacy cannot be tested experimentally (because strictly everytime identical ambient conditions cannot be set up by any experimentor), is it really just a mere assumption? The above argument -- from "existence" to "possible existence" to "necessary existence" -- seems formally to prove such a strict determinism to be the case. Moreover, we may, following Aristotelian metaphysics, hold that natural laws are in fact descriptions of regularities that take place on the basis of, i.e. as an effluence from, the specific intrinsic nature or essence of things, that is to say, a thing acts and reacts necessarily according to its intrinsic nature. So the assumption of strict determinacy boils down to assuming that the specific nature of any given thing is fixed and constant. If it changes, it is replaced by some other nature, and that new nature is not the nature of that same thing anymore but of another thing.
But this statement is in fact a tautology (i.e. a statement that is always true independently of the truth of its component propositions). It says, in effect, that because the nature of a thing is always the same it doesn't change. In the present context 'a given thing' is always meant to be some given individual of a particular species of thing, and here a constant nature is presupposed ( See First Part of Website, Back to Homepage, in the document on the "Species-Individuum Structure" [of things] ). So the constancy of a nature, and thus the prevailing of strict determinism, is still not more than an assumption, an assumption, by the way, that makes natural science possible at all, even when this constancy would only be merely approximated (instead of kept to) by Mother Nature. That is to say, as long as a 'wavering', or state of undefinedness, of the whatness of things is indeed minimal, - to do natural science is possible (because only then it can base its theories upon repeatable experiments and observations). So in natural science the assertion of the (immanent or transcendent) existence of true "laws of nature" is based on the presupposition that all things have a species-individuum structure, where the 'species' aspect is the fixed intrinsic 'whatness' or quiddity of such a thing, and where "the thing" is taken to be a substance in the Aristotelian-Thomistic sense (i.e. an entity that ontologically does not need a substrate to carry it, and thus an entity that ontologically exists on its own). This species-individuum structure is quite clear in the case of inorganic things such as atoms, molecules, and crystals. As individuals they are true substances, each possessing the essential characteristics of its species. And as seen already in the more complex among them, for instance in crystals, their intrinsic whatness is enveloped as it were by further determinations that are not per se, but accidentally added to it as a result of their individual existence : many crystals exist, as to their shape, in a 'deformed' state as a result of the external conditions : Snow crystals (being just forms of crystallized H2O), for instance, all crystallize in the holohedric Class, 6/m 2/m 2/m, of the Hexagonal Crystal System, as this Class, i.e. the (point) symmetry described by it, is part of their intrinsic and internal form or shape, and as such expressing aspects of these crystals' true, i.e. intrinsic, nature. But the external shape snow crystals actually take up during formation is, in most cases, not that of a hexagonal dipyramid or prism (as we meet them in Antarctic snow) at all, but (their external shape is) often that of irregular and asymmetric star-shaped bodies. As a result, we here see in crystals (in contrast to atoms and molecules) the differentiation (within a single crystal) between (1) the essential qualitative and quantitative structure, representing their true fixed intrinsic nature or whatness, and (2) the accidental determinations, enveloping as it were this intrinsic nature, and being replaceable by other such features without altering thereby the (snow) crystal's intrinsic whatness.
And this state of affairs is supposed to be present in organismic individuals too. Indeed, that was Aristotle's very point of departure of his metaphysics. So every individual organism is to be understood as a true substance, having a specific essence or nature (commonly possessed by all individuals of the biologically defined species to which it belongs) and (having) a set of additional determinations that only per accidens have been taken up by the individual substance. All determinations that are ontologically called "accidents" because their way of being is to inhere (in contrast to independently exist) in substance, divide into two groups : (1) the so-called propria, which are determinations that not merely happen to inhere in the substance in question, but necessarily (with respect to the substance's intrinsic nature) do so, and (2) determinations that just happen to inhere in it, i.e. their inherence (or ontologically being carried) is not necessarily implied by the intrinsic nature of the substance having these accidental determinations. And it is the latter that I take to be true "accidents", while the propria are, as to their content (but not as to their ontology), a true part of the intrinsic nature or form of the substance in question.
And this true intrinsic nature is in the case of organisms -- in contrast to crystals -- the substance's strategy-to-exist, describing not only the organism's behavior, but also its morphology and physiology insofar as these characteristics are commonly possessed by all individuals of the species.
In inorganic substances, such as crystals, this intrinsic nature seems to be, as a result of their prolonged existence and thus of their continued repetition, well established and well defined and thus well fixed and constant in them, that is, their behavior has grown into a well-stabilized 'habit' already for an immeasurably long time. And so, as to their behavior, they follow "natural laws". In organisms, on the other hand, their intrinsic nature is not, as in crystals, a mere static qualitative structure or content, but a dynamic strategy, a strategy to materially exist and persist in the Explicate Order. In contrast to crystals, o r g a n i s m s actively maintain and secure their material existence. And by having, in the Implicate Order, evolved into intricate strategies, their material expressions are necessarily highly complex and therefore thermodynamically unstable. They cannot afford to adopt a state of thermodynamic equilibrium (such as in crystals) in order to be stable, because that would mean their structural disintegration and demise (where then the remains are stable). They must keep themselves far away from this equilibrium state and yet remain stable as well as they can. And they can only accomplish that actively, i.e. by being themselves a (materialized) strategy.
And now it is to be expected that a strategy, as developed in the Implicate Order and as such ready to be projected into the Explicate Order, is -- although of course already being ordered to certain existential conditions in that Order -- still not yet fully worked out. As to this, we might say, actual experience 'learnt' by the newly appeared organismic species in its proper environment, i.e. in its 'assigned' ecological niche, is needed for the Implicate Order to be able to supplement and perfect the strategy-content. A newly (i.e. for the first time) projected strategy-content results in a new species. And the individuals of such a species have a not-yet-fully-defined intrinsic nature. That would mean that while crystals are what they are, namely either H2O crystals, NaCl (salt) crystals, or whatnot according to their specific intrinsic nature, - organisms are, apparently, not what they are, namely either Homo sapiens organisms, chimpansee organisms, Musca domestica organisms, or whatnot according to their supposed "specific intrinsic nature", i.e. according to their specific strategy-content.
But isn't it so that any given organism simply is what it is according to even its still not-yet-fully-defined strategy? That is to say, doesn't any organism behave purely deterministically according to its albeit incomplete intrinsic nature, while at the same time also every organism that does possess a complete, i.e. perfected, strategy-content behaves deterministically according to its well-defined intrinsic nature? This, obviously, does away with the necessity to assume a degree of indeterminism in the world of material substances.
This may be so, - provided we assume an organism to be a machine-like entity, i.e. it metaphysically to be a mere aggregate of constituent true substances in the metaphysical sense. These substances, together making up the organism, are then its constituent chemical atoms, or even -- going still further down into the reductionistic hierarchy -- the sub-atomic particles (or wave-packets for that matter) making up these atoms. This is a reductionistic view as regards organisms, and such a view is not simply wrong because it is reductionistic (or mechanistic for that matter). But perhaps it is wrong, as many observed facts seem to point to. Indeed, even already crystals do have some holistic properties (as I showed in Fourth Part of Website). In star-shaped snow crystals, for instance, there must be some non-local connection between the six arms. And indeed organisms are held by many authors to be holistic entities par excellence, as for instance, their individual development (embryonic development) seems to suggest clearly.
If so, we cannot reduce an organismic individual, as to what it is, to its constituent parts, i.e. it is not its parts : The whole is more than its parts. The organism is an holistic entity or being. So an organism is a fundamental entity all by itself, it is not a mere aggregate of fundamental entities. And as such we may then see it as a true substance with its own intrinsic nature. It is, it is true, composed of atoms, but has thereby become a true single 'fundamental' being or substance. And although its behavior depends in some way on its constituents, it is not simply equivalent to the combined behaviors of its constituent atoms. Its behavior is not composed, but emerges, as a new kind of behavior, from the structural collection of the organism's atoms. And these latter have thereby become virtual atoms instead of real independent atoms, i.e. they have become mere qualities of the organism instead of its constituent particles. These particles are not true particles anymore and have lost their substantiality (which they still had when they were free, and regain when they are freed) and are ontologically integrated into the whole, i.e. into the organism, which now is the new substance. And while the composing (now virtual) atoms have their nature long since established as being carbon atoms, oxygen atoms, etc., the new organic substance, -- having emerged by these atoms composing an organism (and themselves turned from real to virtual), -- has in most cases its nature or essence (its strategy) not yet fully defined when it has for the first time been projected into the Explicate Order. It must further perfect its intrinsic nature as a result of the organismic species' continued experience in the Explicate Order, in order to remain able to exist in that Order. And because this process of adjustment and perfection will never come to a final conclusion (because existential conditions in the Explicate Order continue to change), organisms will never possess such a fully defined and constant intrinsic nature or habit as, for instance, crystals do possess it. Conventional evolutionary theory expresses this by saying that organisms either become extinct at some time or develop into other species. But because the process of speciation is by no means well-understood (because it doesn't exist?), and certainly cannot result, in and by the Explicate Order, in a long-term evolution of new species and types (by the proposed mechanism of random genetic mutation and natural selection), we hold that the part played by the Explicate Order is no more than getting a noëtically produced strategy-content to be revised through a specialization of the corresponding same species within its proper habitat (see below), resulting in it finding, in that habitat, its proper ecological niche. In this we assume that the nature or intrinsic whatness of a newly projected species is still not yet fully determined. That is to say, the individuals of such a species are substances with only a partly determined essence or nature. For this to occur at all, such substances must be holistic entities. And we take all true substances (in the Aristotelian sense) to be holistic entities. Mere aggregates of constituent substances (in the Aristotelian sense), i.e. mere aggregates of atoms, and thus such aggregates being fully reductionistic entities, do have a definite, albeit composed, 'nature' and deterministically behave according to that nature. And so, because all true substances (in the Aristotelian sense) are not such aggregates but are holistic entities, there can exist certain such substances with not yet fully defined essences or natures, and these are the organisms.
So if indeed organisms can be taken to be truly holistic beings (like independent atoms can be so taken), then we may have cases of a certain degree of indeterminacy to be present in the material world.
If, on the other hand, they could legitimately be fully reduced to their atomic constituents (i.e. if their atoms are still real instead of only virtual), they would at all times deterministically behave even according to an incomplete (i.e. not yet perfected) strategy.
But, as has been said, we, on this website, take organisms to be truly holistic entities. Certain versions of their behavior turn into habits as a result of repetition, while others do not (because not sufficiently often repeated). And through injection, the noëtic strategy-strings are further perfected insofar as bringing with it a revised noëtic description of morphological structures involved in this behavior, that is to say, only when the strategy-content is in the Implicate Order again, as a result of injection, the new (behavioral) habit (as it is noëtically codified) can noëtically react with other elements of the injected strategy-content, resulting in the morphological (and / or physiological) structures that sustain the new habit. And through projection of this revised strategy-content, now including new morphological details, into the individuals of subsequent generations (of the species in question) the perfected strategy-content is thus non-mechanically distributed among -- in an increasing degree so, as habituation is strengthening -- all the species' individuals however far apart they may live (i.e. however far apart spatially their morphogenesis may take place).
We have found out that projection of a noëtically constructed strategy-content results in that content to be transmitted or transferred from the Implicate Order into the appropriate ecological niche in the Explicate Order and thus resulting in the transfer of this content to, and into, the subsequent material individuals together constituting an organismic species. By this we mean, of course, that the strategy-content as a result of its projection appears in the Explicate Order in the form of organic individuals (of subsequent generations). And we may further theorize that such a projection is immediately followed by an injection again of this content back into the Implicate Order. And from then on a continued alternation will take place of projection and injection. And each time a projection of that content has taken place, the noëtic strategy-content, as it was present in the Implicate Order, has now turned into an intentional sign with its content -- its meaning -- now being permanently present in the Explicate Order (as long as the organismic species manages to exist there). This constitutes part of the Implicate Order's knowing the Explicate Order, while injection brings back to the Implicate Order new habits, i.e. a revised strategy. In this way, as generation after generation of the organismic species appears, there is a continued alternation of projection and injection between the "station" of the noëtic strategy-content/intentional sign and the "station" of the material individuals (generations) of the corresponding organismic species, revising and adjusting its strategy.
This means that the Implicate Order is in constant contact (through the acts of projection and injection) with the changing ecological conditions of the respective organismic species, i.e. it keeps a record of, so to say, or perhaps better, it continually knows of these conditions.
And on the basis of this knowledge it adjusts, perfects, and stabilizes the strategies of each one of these species by functional specialization or universalization within its overall ecological niche (see for this, next section). And this means that not only the species' strategy becomes more and more defined and stabilized, but also its ecological niche (i.e. also the determination and delimitation of its ecological niche). All this is directly evident in the case of the species' specialization (within its niche, - in fact within its habitat, see further down), but is, indirectly, also clear in the case of universalization (within the species' niche), because universalization here does not mean the strategy to be still poorly defined, but a strategy ordered, in a precise way, not to one, but to a definite series of "targets" (i.e. resources in the environment). This is, for instance, the case in a number of calyptrate flies (such as the housefly), (each single species) being able to individually develop (as larva) in a wide range of different breeding substrates.
The Implicate Order is spaceless and timeless in spite of us having it dynamically described ('construction' of strategies). And because its entities are immaterial, there are no 'individuals' or 'individual cases' in it. In the Explicate Order, on the other hand, spatially and temporally existing individuals and individual cases are typical of it. There, strategies are genuinely active through the behavior of the individuals. And because the Implicate Order is immaterial and non-individual, individual events in the Explicate Order cannot influence immaterial structures in the Implicate Order, i.e. cannot be injected into that Order. Only when an increasing repetition of a given behavioral change in a given organismic species is taking place, i.e. when a more or less new habit is evolving, it will influence the structure of the corresponding noëtically developed strategy in the Implicate Order together with revising or supplementing the noëtic description of the morphological structures functionally involved in the development of the behavioral habit of the species. In fact, injection into the Implicate Order brings with it this revision of the noëtic description. And through projection of the so perfected strategy-content, it will, in the Explicate Order be non-mechanically distributed (finally resulting in a state or condition of material distribution) among the species' subsequent individuals.
This is, then, the non-mechanical inheritance of an acquired habit to subsequent generations of the species. And the 'mechanism' of transmission of such a habit, as this habit was first developed in the Explicate Order, to the noëtically developed strategy-content, through injection, or, said differently, the 'mechanism' of, or the condition for, injection of the new habit into the Implicate Order, is continued repetition of the changed behavior when the latter becomes established in more and more individuals (i.e. when the new behavior is not individual anymore but becoming specific). Only through projection of the in this way further defined strategy-content all subsequent individuals of the species will acquire it, i.e. they will inherit it non-mechanically.
Habitat and ecological niche
We spoke about "specialization within the ecological niche". Maybe we should qualify this a bit more.
First of all, an ecological niche is a specific part, aspect, or station inside an organism's overall habitat. A habitat of a given organismic species is the overall environment that satisfies the general and minimal conditions for such a species, and for a number of other species as well, to live in. In it, the ecological niche is a precisely defined existential condition for a particular organismic species (which condition, like the habitat itself, may be instantiated in more than one different geographical regions). This species can deal with the biotic and abiotic elements of that niche in order to guarantee its continued existence. It can do so because it has (or is) a strategy to get hold of and exploit special resources present in the niche, resources that actually realize its prolonged existence. These resources consist in means of shelter and in food. Most important are the conditions that guarantee reproduction. These conditions first of all imply the necessity of the organism to reach maturity and the possibility and opportunity to mate, and are laid down, or expressed, in its morphology, physiology, and behavior, in short, in its strategy. Its whole strategy, as it is commonly possessed by all the species' individuals, must precisely match the elements of its ecological niche. So, in contrast to a mere habitat of a given organismic species, an ecological niche is defined such that every element in it has its counterpart in the strategy of the species, and all elements of that strategy have their respective counterparts in the elements of that ecological niche as it is present in the species' habitat. Said differently, the whole morphology, physiology, and behavior of the individuals of a given organismic species must reflect all its niche's elements. In an ecological niche, so defined, there are no irrelevant elements. Strategy and niche match precisely.
So we cannot in fact speak of "specialization within the ecological niche", because that would mean that this niche was not yet properly defined with respect to that organismic species. So we'd better speak of "specialization within the species' habitat" (which comprises potential or actual niches of other species as well ). And it is the habitat of the species that is the "not-yet-properly-defined ecological niche", and the species must find in it its own properly defined niche.
We might theorize, then, that the for-the-first-time-projected strategy-content ends up -- as a collection of material individuals of a particular organismic species, a materialized strategy -- as a rule not directly in its proper ecological niche, but in its proper habitat. And while being still a crude strategy it can live in it. But for guaranteeing its prolonged existence, it must, however, find its own 'place' within its habitat, i.e. it must find its ecological niche. And in doing so it (i.e. the species) must 'explore' its habitat and eventually set up its precise ecological station inside the habitat. And this is precisely what we should have meant by "specializing within the ecological niche". This specializing consists of increasingly emphasizing a narrower defined behavior ordered to exploit certain selected resources actually present in the habitat among all the other potential resources of that species. These chosen resources are precisely those that are not yet exploited (in the same way) by other organismic species, and so by selecting precisely them, competition is avoided. And as soon as more and more individuals do so, their behavior becomes a habit of the species and will, upon injection, supplement the noëtically constructed strategy-content in the Implicate Order including [supplementing] its noëtic description of the functional morphological structures (and of the physiological ones) directly involved in the new habit. And then, upon projection, the habit, including the mentioned morphologies (and physiologies), will be non-mechanically distributed among individuals of subsequent generations of the species. And now, at last, the species has found and occupied its proper ecological niche, and thus having secured its prolonged existence. Thereby we may assume that all this takes place in a step-by-step fashion, so that the species gradually secures its ecological niche within its habitat.
Of course, not in all the cases of appearance of a new species there is need for such a gradually homing-in into that species' properly defined ecological niche. On the contrary, in most cases where, as described earlier, a certain 'ecological hole' (i.e. a special ecological opportunity or possible existential condition) is to be occupied (ecologically exploited), we have to do, not with a mere (potential) habitat, but with a narrowly defined potential ecological niche, that can only be exploited by a very specialized organismic species indeed.
Appearance of new orgamismic t y p e s in the course of geologic time. A problem.
One might ask, why weren't there birds in the luxuriant carboniferous forests ?
Conventional theory will answer that the (morphological and physiological) organization in birds, being the first warm-blooded animals, took a long time to evolve, evolve that is, from certain (appropriate) reptilian ancestors. So birds could not appear before (upper) Jurassic times. The same sort of thing could be asked about, say, the insect Order Diptera (mosquitoes, midges, true flies) : Knowing that they today are utterly successful and to be found in almost every conceivable habitat, we may ask why they were totally absent (as far as we know from the fossil record) in Carboniferous times (of which, it is true, we know only the fauna of the tropical swampy lowlands) as well as (totally absent) in the Permian and in most, if not all, of the Triassic. Did their evolutionary development all the way up from carboniferous ancestors of the Order Mecoptera (Scorpionflies) take so much time that true Diptera could only emerge in the lower Jurassic, long after the origin of say, Orthoptera (locusts, grasshoppers), Homoptera (cicadas, leafhoppers, etc.), and many others? And the same goes for the Order Hymenoptera (wasps, bees, ants) : They are not known until the upper Triassic. But we can perfectly well imagine the coexistence of wasps (not bees), flies, and birds in a carboniferous forest. Also with respect to plants we might ask why the Angiosperms (plants with flowers) did not appear until the Cretaceous. And we may expect the same sort of answer : It took a long time for them to evolve all the way up from Gymnosperms (plants with no true flowers, such as pine-trees).
This type of answer to such questions is in itself perfectly reasonable, but is not actually demonstrated to be right.
Also, or even in particular, for our theory of evolution such questions are quite embarrassing. For we hold that all strategies are present 'already right from the beginning', at least in the form of noëtic descriptions or strings, residing in the Implicate Order and appear as organismic species in the Explicate Order as soon as their ecological niches are present in that Order. But were then in the carboniferous forests no such niches present for any species of bird, fly, or wasp? Even in our theory it is clear that changing climatic conditions, marine transgressions and regressions, changes of elevation of regions, etc. will cause the destruction of existing ecological niches and the creation of new ones, resulting in the extinction of species and the appearance of new ones. And this indeed, accounts for the succession of species as we see it in the fossil record. It does not, however, account for the initial total absence of entire organismic types such as birds or flies, and their more or less sudden appearance and continued persistence much later on. Organismic species will, in the course of time, replace one another as a result of changing external conditions. So why do we, apparently, not see (as far as the fossil record goes) certain bird species appearing intermittently amidst the appearance of species of other organismic types? We can perfectly well imagine that for particular species of birds no resources existed in carboniferous forests. But we cannot imagine that they were absent for all species of bird (and the same goes for flies and (solitary) wasps). Maybe the necessary ecological niches in carboniferous forests were already occupied by other species than birds. But this cannot be so, because a same niche harbors a same species. Ecological niches of different organismic species may, it is true, partly overlap (same resource being exploited, but all other elements of strategy being different), and this may lead to competition which might result in the expulsion of one species by another. And why couldn't the latter be a species of bird? We, within our noëtic theory of evolution, do not see why this would be forbidden. So our theory, as it stands until now, allows for, and even predicts, the intermittent appearance of certain species of birds, and also of certain species of Hymenoptera and Diptera for that matter, already in carboniferous forests. This prediction, however, has not come true (insofar as at least the fossil record tells us). One is pretty sure that no birds, wasps or flies were already existing in carboniferous times. Neither were they in Permian times. The reader may judge for him/her-self whether this uncovers a fundamental flaw or weakness of our theory, or whether it is not more problematic than the assumption of conventional theory that organismic species and types materially evolve from one another and that some of these evolutionary developments take a long time indeed. This assumption is not in itself implausible, i.e. that evolutionary development of a new organismic type may take a long time, but the assumed evolutionary development itself becomes implausible as soon as it is considered to be a material process. And, moreover, why took the development of birds (having appeared for the first time only in the upper Jurassic) from, ultimately, carboniferous or permian reptiles so much longer than, say, the development of true dynosaurs (having appeared for the first time already in the Triassic) from these same reptiles? Or why took the evolutionary construction of the wasp- or fly-strategy so much longer than that of the strategy of the members of the (now extinct) Order Palaeodictyoptera (large winged insects, abundant in carboniferous forests), or [took so much longer] than that of the highly specialized Megasecoptera (also large winged insects present in carboniferous forests, and especially later on, in the Permian) ? If we, following our theory, reject the assumption of species and types having materially descended from one another, the initial absence and later appearance of new organismic types must be explained in some other way. As we have shown, many such cases cannot be explained by changing ecological conditions as our theory would suggest. In some way the order of appearance of organismic types and their fates in the Explicate Order must make up some precondition for the appearance of new types. That is to say, earlier types pave the way for new types to appear. So for new organismic types to appear it is important what had appeared before. And this, as such, independently of changing small-scale ecological conditions. The latter determine the appearance of new species of the same type. Maybe, earlier types in some way set up the large-scale ecological conditions for next types. And the latter may then coexist with, or replace, the former. In the case of bees this is clear : Only after angiosperm plants had appeared, the bee-type of Hymenoptera could appear, because bees are -- as "vegetarian wasps", or " flower wasps" -- totally ordered to, and dependent on, flowers as a source of nectar. Unfortunately, in most other cases of the appearance of new organismic types matters are not so clear. In some cases it may be the demise of a type that sets the conditions for a next type to appear, while in other cases it may be the very blooming of a type that sets such conditions (for example the blooming development of grassland vegetation in the early Tertiary, setting the stage for grazing mammals). The demise of a type may, however, not be considered to 'freeing' particular, formerly occupied ecological niches, because these can only be re-occupied by the same species again. So the demise of a type must create new ecological niches, i.e. niches of a fundamentally new nature that can receive species of a new organismic type.
So while species appear and replace each other within a given overall organismic type, the types themselves appear and often replace each other within the Whole of Life.
All this conjecturing does not pretend to offer an "explanation" in terms of our noëtic theory of evolution. It is just a proposed ecological description of the demise and appearance of organismic types in the course of geologic time. But if this description does contain some elements of truth, the appearance of new organismic types is, like that of organismic species, dependent on existing states of affairs in the Explicate Order. And the appearance in geologic history of new organismic types is only in so far a contribution (to evolution) attributed (by our theorizing) to the Explicate Order insofar as that appearance depends on ecological settings. On the other hand, stabilization and specialization of a strategy within the habitat into which it is projected do supplement its noëtic construction and are as such a creative contribution attributed (by our theorizing) to the Explicate Order.
Mechanical and non-mechanical medium of organic evolution.
With all this, we have now extended the part played by the Explicate Order in creating organic strategies. But it is still the Implicate Order which does the actual 'construction'. Indeed, although behavior of organismic individuals may change or become more strictly defined, the Explicate Order is not capable to construct or adapt the morphological structures involved in this behavior [i.e. structures that sustain, stabilize, or further enhance a new behavioral habit]. And indeed, a mechanical medium, as is the Explicate Order, cannot, all by itself, actually construct any functional morphological structure, because it does not contain a teleologic aspect. This problem is surely recognized and acknowledged by conventional evolutionary theory. To solve it mechanically nevertheless, conventional theory has proposed the idea of random genetic mutation and natural selection of useful mutants. In this, the phrase "natural selection of useful mutants" is in fact a mere tautology. It says no more than what is finally left of such mutants are the ones that could better reproduce, that is, those ones that become more in number are precisely the ones that are more in number. So in the creation of functional structures the focus must lie in random genetic mutation. And indeed, random mutation can create everything. But because "creating everything" is equivalent to "creating nothing at all," there is the natural selection of only the successful ones. This selection is just automatic because "biologically more successful" is equivalent to "creating larger numbers" (of individuals). But the creative power of spontaneous genetic mutation is limited. It cannot account for long-term evolution of the many intricate, and especially, clever adaptations or strategies as we encounter them in the majority of organisms, as we had argued earlier. Therefore, they must have been 'constructed' in a non-mechanical medium, which we, following David Bohm, have called "the Implicate, or enfolded Order", or "noëtic space".
(will be continued if necessary . . .)
Subsequent to the conclusion (part XIV) of the functional wing-types in insects, that is, beginning (still in the present series of documents) with part XV, Basic Features of the Inorganic World . . . , we will develop the general theory of the Explicate-Implicate Orders still further, by investigating the basic features of the inorganic world (which is, of course, the broader existential context of organisms). We will inquire whether, and if so in what way, notions like intensive qualities, holism, and being-a-substance do refer to noëtic space, i.e. to the Implicate Order (i.e. do actually involve that Order). Especially, it will be demonstrated that the import into physics and chemistry of (1) the existence of intensivity (existence of intensive qualities, not only in heat and light, but even -- as impetus -- in local motion, and in mass -- in the form of density), of (2) the holistic constitution of certain inorganic beings like atoms, molecules and crystals (and with it the virtual existence of their constituent particles), and, therefore, also (the import into physics and chemistry of ) the existence of true substances (in the metaphysical sense) even in the inorganic world, will not have any effect on the content and truth of (1) classical atomic theory (i.e. the theory of chemical elements and their compounds, in short, the theory of atoms and molecules), of (2) conventional crystallography, and, finally, (on the content and truth) of (3) classical mechanics (still applicable to macroscopic objects) and quantum mechanics. And that means that modern physics and chemistry can be viewed as specifications, not of any form of democritean mechanical and reductionistic metaphysics, but of the non-mechanical and holistic metaphysics of Aristotle and St Thomas. And, certainly, our noëtic theory of organic evolution does fit into all this very neatly indeed!
In addition to this "natural philosophy" we will, also in the series of documents following upon part XIV, give an exposition of true metaphysics, i.e. a discussion of "Being, only insofar as it is Being". And then, finally, we will work out the "theory of qualitative space-points", i.e. a theory about the changing distribution of different qualities among points making up discrete space and underlying all of Being.
In the next document, that is going to be about Strepsipterygia in insects, we explain our temporary interruption of the series on insects (here reproduced) :
Important Remark on the succession of treatises and subjects :
[Being now the summer of 2010, we still have to finish the exposition of all existing, and having existed, functional wing-types in insects. But we have temporarily interrupted the work on these expositions, in order to first work out further the broader physical context of living beings. This we are, at the moment, doing in a series of documents from part XV of the present Series onwards (thus from the part following upon the document that is going to be about the last wing-type (Dipterygia)). And these documents concerning the physical context of organisms and evolution are in fact treatises on Natural Philosophy. They will concern fundamental things like Time, Place, Space, Motion, Quantity, Continuity, Quality, Extensionality, Intensionality, Dimensions, and the Aristotelian (versus Mechanical) explanation and interpretation of these and other things. The findings will be integrated into our theory of the Implicate-Explicate Orders.
This Natural Philosophy, worked out along Aristotelian lines, will be followed by precisely that of which this natural philosophy is supposed to be a specification, namely the Theory of Being qua Being, chiefly how it was originally worked out by Aristotle in his Metaphysics. It is about beings and sorts of beings, but only insofar as they are beings. It will be seen that Being is found in the form of groups of so-called "pros hen equivocals", i.e. groups of instances of fundamental types of beings with one primary instance in each group to which all the members of the group refer.
And this Theory of Being qua Being will then be followed by a theory delving into the very foundation of real Being, and thus not dealing with Being qua Being, but with Being insofar as it is material things and patterns, i.e. insofar as it is qualitative content as such. It is the Theory of Qualitative Space-points, space-points that ultimately underly and make up the things and patterns in the Explicate Order. It will dissolve these things into 'qualitative points' making up the discrete space of the Explicate Order. This means that the reality of substances (atoms, molecules, crystals, and organisms) is on its most basic level a discrete space consisting of qualitative points, i.e. a space completely made up by these points. And because these points are kept at unit-distance from one another they will form a regular point-net. And precisely this opens the way to see the Explicate Order with its patterns and substances as the computational result of an overall Cellular Automaton (CA) : The current configuration or patterning of different qualitative content in discrete space, i.e. the configuration, in discrete space, of the points ('cells') as to their current qualitative content, of the present moment is the result of the transformation of the configuration of these qualitatively differing points ('cells') as that configuration was in the previous moment. And these successive transformations of one configuration of 'cells's into another is governed by "rules of projection" (from the Implicate Order into the Explicate Order), i.e. CA-rules. These overall cellular automaton rules are the result of the integration of countless other rules including those that have to do with organismic strategies and also with the constitution and constancy of other substances as they are being projected into the Explicate Order. The concept of discrete, but nevertheless 'gapless' (voidless) space will be expounded by working out an argument against even the logical possibility of the actual infinite, physically as well as mathematically. And for the successive transformations of qualitative configurations we will refer to the part [i.e. the document] having dealt with cellular automatons already in our First Part of Website.
And only when all this has been done, Natural Philosophy, Theory of Being qua Being, and the Theory of qualitative Space-points, we will return to insects, their wing-types, and their evolution, now within a firm philosophical setting and background.]
And so, having developed and concluded our noëtic theory of evolution, we have finally found some directives as to how actually proceed in interpreting fossil and recent insects, and all organisms for that matter, and laying bare, in more or less general terms, the noëtic construction of their strategies in the Implicate Order, and the part played in all this by the Explicate Order, and thus accounting for their morphology and behavior in the latter Order.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of the types of flight-devices in insects, Part IX, Strepsipterygia.
Back to first part of theoretic intermezzo