
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Most expressed example of the type :

Description of the type
Morphological features
Retained is a markedly expressed homonomy of the wing pairs and of the meso- and metathorax together with the muscular apparatus. Wings elongate, at the apex more or less rounded. Base of both pairs distinctly narrowed in the form of a more or less individual handle, basiala. The wing-venation in the majority of representatives of the type shows little costalization and a more or less equal strengthening of the anterior and posterior margins of the wing. The absolute number of veins is small, and a reduction of their number takes place. Body elongate, with an extended more or less cylindrical, sometimes flexible, abdomen, and a large head with big eyes. Size of the insect medium or large, not less than 20 mm. Wings about equal to the body, sometimes shorter.
Functional features
Both pairs of wings participate in the flight-movements as organs supporting the body and creating traction. Representatives of the type are investigated in which there is a marked asynchronicity in wing-beat during flight (Odonata-Zygoptera, Lestidae [and allies]). The features of flight in the other members of this type (Mecoptera-Bittacidae) are not yet clarified. Speed of flight is also known only of Lestidae [and/or allies] and varies between widely separated values, reaching up to 14 m/sec., but normally much less, 1-2 m/sec. (Coenagrion sp.). Governability in flight is high, and was especially investigated. It is realized by a wide change of angles of the slope of the plane of wing-beat and of [the plane of] the wing-blade. The load on a unit of wing-surface is rather low and known only of the Zygopteron Coenagrion puella, measuring up to 0.0120 gr/cm2.
The biological significance of flight is really great, at least in Zygoptera. These insects are active predators to which flight is important for catching prey. As to the other representatives of the type, the Mecoptera-Bittacidae, there are observations that they ambush their prey passively.
Differentiations, connections, and representatives of the type.
This type, paddle-wingedness or kopepterygia, characteristic by the formation of distinctly individualized narrowed basal parts of the homonomous wings [implying that the wings are truly petiolate], is today not widely distributed, and is rather uniform. Two basic sources of its formation are evident : On the one hand long-wingedness ( dolichopterygia ), and on the other - certain forms of neuropterygia. The forms that have originated from dolichopterygia are the most diverse and are characterized by a relatively rich venation, having undergone far-reaching mechanical specializations.
The different forms of kopepterygia may be divided into three subtypes :
1. Prokopepterygia
The first most peculiar subtype, not represented in the recent fauna [except, perhaps, wings as we see them for example in Calopteryx (recent, Odonata-Zygoptera), is characterized by the moderately developed narrowing of the basal parts of the wings and a reduction of the venation having acquired a markedly expressed costalized nature [absent in Calopteryx]. The abdomen carries long threadlike cerci [as such absent in Calopteryx] having a definite significance in flight. This subtype, ancient paddle-wingedness, or, equivalently, prokopepterygia (pro-kope-pterygia), is in fact very peculiar, and may even perhaps be distinguished as an independent type of its own. To it [exclusively] belong extinct paleozoic palaeopterous insects [= insects that cannot fold their wings over the abdomen] -- Megasecoptera, Protohymenoptera [despite the name not ancestors of Hymenoptera. All the species of the 'Protohymenoptera' are now considered to belong to the Order Megasecoptera (suborder Protohymenoptera) ], and Sypharopteroidea. See next Figures.
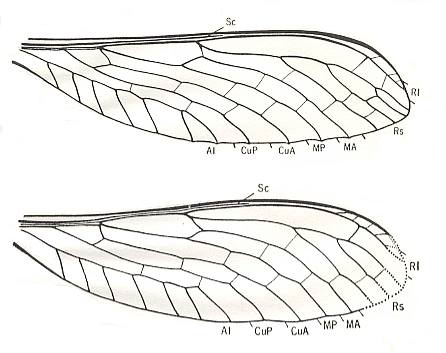
Figure 1 :
Top image - Alexahymen maruska Kuk. Family Bardohymenidae, Order Megasecoptera. Hindwing. Length 30 mm. Width 9 mm. Lower Permian of Czechoslovakia.
Bottom image - Alexahymen maruska Kuk. Hindwing. Length 31.8 mm. Width 9.9 mm. Lower Permian of Czechoslovakia.
In both these wings it is uncertain whether or not the wings were (moderately) petiolated.
(After KUKALOVÁ-PECK, 1972.)

Figure 2 :
Top image (1) - Fore- and hindwing of Protohymen permianus Till. Family Protohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). Forewing length 10-13 mm. Width 3 mm. Hindwing length 9-11 mm. Width 3.5 mm.
Middle image (2) - Forewing of Protohymen elongatus Carp. Family Protohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). Forewing length 20 mm. Width 3.5 mm.
Bottom image (3) - Fore-and hindwing of Permohymen schucherti Till. Family Protohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). Forewing length 13 mm. Width 3.5 mm. Hindwing length 12 mm. Width 4 mm. The wings of this species are a little less typical of prokopepterygia, but nontheless should be included in this subtype. The same applies to the wing depicted in the next Figure.
(After CARPENTER, 1930.)

Figure 3 : Photograph of holotype of Asthenohymen kansasensis Till., 1926 (Doter minor Sell., 1907) Family Asthenohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). Lengh 7 mm. Width 2-3 mm.
Although this wing clearly belongs to the Megasecoptera, it is not a petiolate wing, not even moderately so. We may assume that it is in the evolutionary line eventually leading to petiolate wings. The Asthenohymenidae are known to have been able to fold their wings back over the abdomen. The widening of the anal area of the forewing might be connected with this ability.
(After CARPENTER, 1930.)
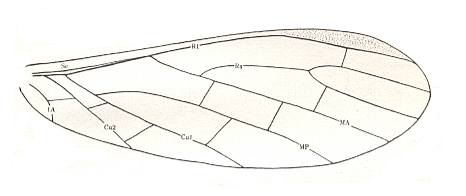
Figure 4 : Wing of Asthenohymen pusillus Till. Hindwing. Family Asthenohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). Length 5 mm. The small dimension is noteworthy.
(After CARPENTER, 1933.)
The order Megasecoptera, with their typically basally-narrowed, and largely homonomous wings, were already present in great diversity in upper carboniferous times. Carpenter, 1951, has revised the Megasecoptera found in the upper Carboniferous of Commentry, France. Before listing and figuring the wings of these species, let us insert here some general remarks of him concerning them :
All of these species belong to that series of Megasecoptera which were strictly palaeopterous, i.e., unable to fold their wings over the abdomen at rest. I have previously discussed the composition of this series in my account of some Megasecoptera from the Permian of Oklahoma (Carpenter, 1947). The palaeopterous nature of these insects is not inferred from the venational pattern but is indicated by the position in which the wings are preserved in the fossils. Neopterous insects [insects which do fold their wings over the abdomen at rest], as a general rule, die with the wings held in the resting position, over or along the abdomen, whereas the palaeopterous types die with the wings outspread. All of the specimens of Megasecoptera treated in this paper are preserved with their wings outspread, which is especially significant in cases where there are several fossils of one species.
Although no member of this particular series of Megasecoptera is entirely generalized [i.e. original or primitive] in its characteristics, the Aspidothoracidae seem to have more such features than any other family, e.g., the nearly even distribution of cross-veins over the wing, the independent origins of main veins and the complete absence of anastomosis of veins. On the other hand, the crowding of SC, R1, and the basal stems of M and Cu along the anterior margin is a specialization which, although appearing in some other members (such as the Permian Bardohymenidae and Protohymenidae ), is absent in other Commentry families. Within this series of Commentry species, also, there is a tendency for wing petiolation, which reaches its maximum development [in Commentry species] in the Sphecopteridae. The occurrence of wing maculations in some of the Commentry species is especially noteworthy, such markings being unknown in Permian Megasecoptera.
The body structure of the Commentry Megasecoptera is very incompletely known. Handlirsch's reconstructions of Mischoptera and Corydaloides, which were published in Schröder's Handbuch der Entomologie (1921) and have been frequently reproduced in geological and entomological texts, are highly imaginative and misleading. Since Handlirsch did not examine the actual specimens which he restored, the results could hardly have been otherwise.
Unfortunately, nothing is known of the immature stages of the Commentry Megasecoptera. Nymphal forms of Brodiidae, with wing pads in various stages of development, have been found together with adults, in Carboniferous strata of England. This record, along with the wing and body structure, seems to refute Forbes' contention (1943) that all Megasecoptera were Holometabola, a conclusion apparently based on the arrangement of veins constituting the Radial Sector.
After each Figure or Figures depicting the wings of a particular family of Commentry Megasecoptera we will add Carpenter's diagnosis of the family as far as the wings are concerned, and the supposed affinities with other families.
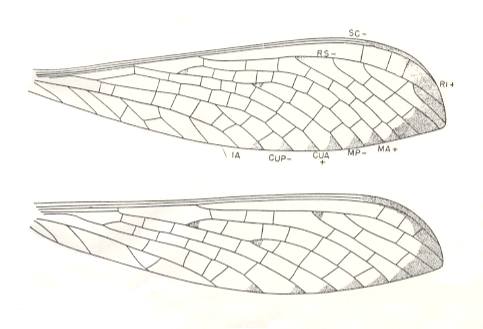
Figure 5 : Aspidothorax triangularis Brongniart. Family Aspidothoracidae. Order Megasecoptera. Upper Carboniferous of Commentry, France. Top : Forewing of specimen C1. Bottom : Hindwing of specimen 17-4.
Forewing : length 31 mm, width 8 mm. Moderately slender, widest beyond middle.
Hindwing : length 20 mm in specimen 17-4, 32 mm in 17-5, 35 mm in 17-7. Width 8-8.5 mm. The wing is shaped very much like the forewing, but is slightly broader proximally.
As in Sylvohymen (lower Permian), in the wings of Aspidothorax the strengthening of the anterior wing-margin (costalization), together with the pectinate branches of the Radial Sector followed by the rest of the main branches parallel to the latter and to themselves, as such expressed especially in the forewing, is certainly a mechanical specialization, and may point to the wings not acting as mere paddles during flight, but as true organs of traction. And this already in upper carboniferous times!
(After CARPENTER, 1951)
Aspidothoracidae.
Morphology of the wings. (MA = anterior Media, MP = posterior Media)
The fore and hindwings are alike in venation. SC terminates well before the apex of the wing. SC and R1 very close together and submarginal. Stem of M very close to R basally but separate from it. MA free from RS and not diverging towards it. Stem of Cu very close to that of M, but not fused with it. CuA not diverging towards MP. One anal vein. Cross-veins numerous and nearly uniformly distributed over the wing area.
Affinities.
The family Aspidothoracidae appears to be most closely related to the Bardohymenidae from the lower Permian of Russia and that of Oklahoma. Both families have the crowding of SC, R1, and the stems of M and Cu, as well as the independence of RS and MA, and MP and CuA. Cross-veins are fewer in the Bardohymenidae, but those present are nearly uniformly distributed over the wing. More remotely related are the Permian Protohymenidae and Scytohymenidae, which are specialized by further crowding of the veins mentioned and by anastomosis of MA and MP with RS and CuA respectively.
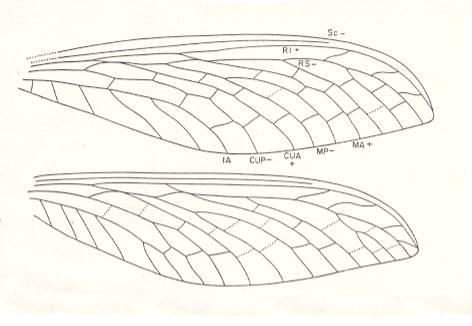
Figure 5a : Corydaloides scudderi Brongniart. Family Corydaloididae. Order Megasecoptera. Upper Carboniferous of Commentry, France. Drawing of fore- and hindwings, based on all known specimens. Veins not preserved are represented by broken lines. The forewing, complete, is 48 mm long and 14 mm wide in the type [specimen], and is approximately the same in other specimens. The hindwing has the same dimensions, but is broader proximally, and has a wider anal area.
These wings are only very moderately specialized mechanically. As in all prokopepterygian wings, they are not truly petiolate (as far as can be made out), but do have pointed apices indicating mechanical specialization.
(After CARPENTER, 1951)
Corydaloididae
Morphology of the wings.
Fore- and hindwings alike in venation, but the hindwing slightly broader basally. SC remote from the margin and terminating in the costal area well before the apex as in Mischopteridae. R1 remote from SC. Stem of M coalesced with R. MA anastomosed for a very short interval with RS, and CuA anastomosed with MP for a longer distance. One anal vein. A few weak cross-veins in costal and subcostal areas. Cross-veins between other main veins arranged in three rows.
Affinities.
The Corydaloididae combine a number of generalized and specialized characteristics found elsewhere in the order. The relatively broad costal and subcostal spaces are generalized traits, also occurring in the Sphecopteridae and Mischopteridae. The nearly even distribution of cross-veins is suggestive of the Aspidothoracidae. On the other hand, the anastomosis of CuA and M as well as of RS and MA is a specialization that is unknown in other Commentry Megasecoptera except for Ischnoptilus. The Corydaloididae appear to have been derivatives of the same group that gave rise to the mischopterids, before the acquisition of the serial cross-veins.
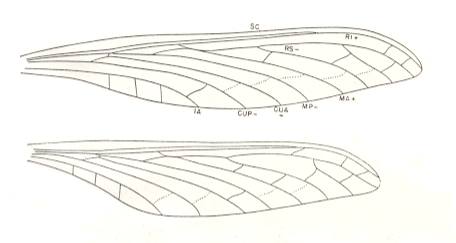
Figure 5b : Sphecoptera gracilis Brongniart. Family Sphecopteridae. Order Megasecoptera. Upper Carboniferous of Commentry, France. Fore- and hindwings of specimen 16-5. Veins not preserved are represented by broken lines (the cross-veins so designated are preserved in Sphecoptera brongniarti Meunier). The forewing is 57 mm long and 10 mm wide (maximum). The hindwing has the same width but it is slightly shorter with a length of 50 mm.
(After CARPENTER, 1951)
Sphecopteridae.
Morphology of the wings.
Wings slender, petiolate, and the venation of fore- and hindwings similar. SC remote from margin and terminating on R1 a short distance beyond mid-wing. R closer to SC than in Mischopteridae. Stem of M coalesced with R. MA usually anastomosed with RS for a very short distance, rarely (hindwing only) diverging towards RS and not quite touching it. Stem of Cu remote from R + M. CuA independent of MP. One anal vein. Cross-veins apparently absent from costal and subcostal areas, but forming only one complete row in the main part of the wing.
Affinities.
The sphecopterids appear to have been derived from a megasecopterous stock more generalized than any actually known. The relatively wide costal space eliminates the Aspidothoracidae as an ancestral line and suggests the Corydaloididae. However, the latter have an anastomosis of MP and CuA, not even indicated in the sphecopterids. The petiolation of the wings of Sphecoptera and Cyclocelis (both belonging to the present family), and the termination of SC on R1 are specializations of this family.
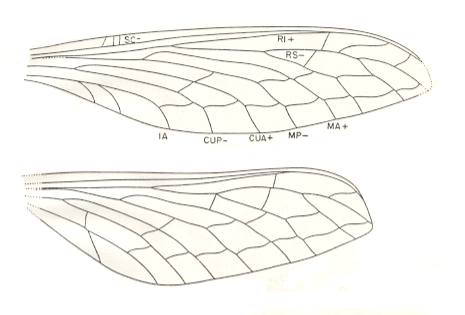
Figure 5c : Foriria maculata Meunier. Family Foririidae. Order Megasecoptera. Upper Carboniferous of Commentry, France. Fore- and hindwings of type [two specimens, both were designated to be type-specimen of the species]. The forewing is 35 mm long as preserved, and 9 mm wide. The entire length was probably around 38 mm. The hindwing is 30 mm long and 10 mm wide (maximum), and is broadest in the region of the end of 1A.
The most striking feature of the foririid wings, the differentiation [heteronomy] of the two pairs, is unique among the known Carboniferous Megasecoptera (Carpenter, 1951). Although narrowed in their basal portions, the wings are not truly petiolate.
(After CARPENTER, 1951)
Foririidae.
Morphology of the wings.
Hindwings broader and shorter than the forewings, but with the same basic venation. SC remote from the margin and terminating on R1 as in the Sphecopteridae. Costal and subcostal spaces about equal in width. Stem of M close to R but not fused with it. MA free from RS. Stem of Cu very remote from M. CuA not diverging towards MP. One anal vein. Cross-veins close together and straight in costal and subcostal areas. The others strongly sigmoidal and arranged so as to form two rows.
Affinities.
The Foririidae, though incompletely known, appear to be derivatives of the ancestral stock of the Mischopteridae. This is indicated by the serial arrangement of the sigmoidal cross-veins and the relatively broad costal and subcostal spaces. The absence of anastomosis of RS and MA, however, indicates a more generalized stock than the Mischopteridae, while the distinct termination of SC on R1 is suggestive of the sphecopterids. The most striking feature of the foririid wings, the differentiation of the two pairs, is unique among the known carboniferous Megasecoptera.
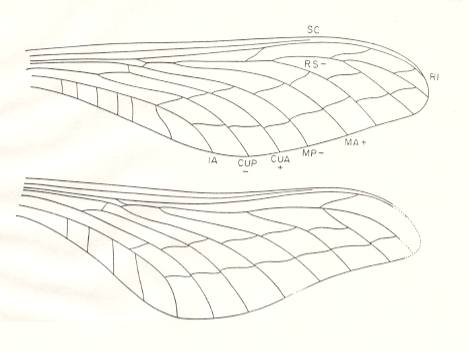
Figure 5d : Psilothorax longicauda Brongniart. Family Mischopteridae. Order Megasecoptera. Upper Carboniferous of Commentry, France. Fore- and hindwings, drawing based mainly on specimen 13-8. The forewing, which is complete in specimen 13-8, is 62 mm long and 18 mm wide (maximum). The hindwing, preserved in specimen 13-8, is completely known except for its very apex. It is about the same length as the forewing and has a maximum width of 20 mm.
(After CARPENTER, 1951)
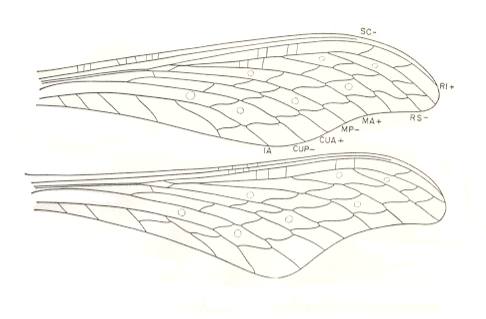
Figure 5e : Wings of Mischoptera nigra Brongniart. Family Mischopteridae. Order Megasecoptera. Upper Carboniferous of Commentry, France. Forewings [of the species M. nigra], length 73-75 mm, with a maximum width of 20 mm. Hindwings about 5 mm longer than the forewings and slightly broader. Wings like these anticipate wings of the next subtype, hyperkopepterygia (see below).
(After CARPENTER, 1951)
Mischopteridae.
Morphology of the wings.
Venation of fore- and hindwings similar. SC remote from margin and terminating before the apex of the wing. R and R1 remote from SC, the subcostal and costal spaces being of equal width. Stem of M very close to R basally but separate from it. MA anastomosed for a short distance with RS, near the latter's origin. Stem of Cu removed from that of M. CuA independent of MP and not diverging towards it. One anal vein. Cross-veins regularly arranged, close together in costal and subcostal areas, but forming two or three rows over most of the wing.
Affinities.
The family Mischopteridae, which includes the largest and most spectacular of the Megasecoptera, is more specialized than the Aspidothoracidae in nearly all respects, although there is less crowding of the anterior veins. The serial arrangement of the cross-veins and the anastomosis of RS and MA are suggestive of the Protohymenidae and other permian families, as well as the Sphecopteridae. The falcate wings and nygmata-like structures of Mischoptera are apparently not family traits, but specializations attained by a particular group of its species.
(continuing the description of the subtype prokopepterygia)

Figure 6 : Calopteryx splendens. Family Calopterygidae, Suborder Zygoptera, Order Odonata. male.
50 mm. Wing expanse 70 mm. Recent. Wings without a true petiole, but nevertheless having the anal area of fore- and hindwings not expanded : As in all prokopepterygian wings, the posterior wing-margin smoothly continues (its course) along the posterior marging of the anal area, which, as a result smoothly tapers toward the wing-hinge, without forming a petiole. The outline of the hindwing of this recent example of prokopepterygia is best comparable with that of the hindwing of the carboniferous genus Foriria.
(After SEVERA in Thieme's Insektengids voor West- en Midden-Europa, 1977.)
2. Eukopepterygia
The other most widely distributed subtype, also a derivative of long-wingedness, is characterized by well developed narrow basialae of the wings [i.e. the wings become truly petiolate]. The wing-venation does not show traits of costalization, but possessing, together with that absence, a far-reaching peculiar mechanical specialization. The body is very elongate indeed, but lacking long cerci [perhaps apart from later discovered permian truly petiolate Megasecopterous wings]. This subtype, true paddle-wingedness, or, equivalently, eukopepterygia includes almost the entire group of damselflies, Odonata-Zygoptera (except Agrionoidea) and their permian precursors Protozygoptera. See next Figures.
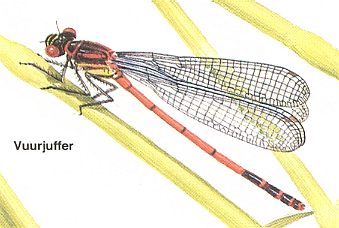
Figure 7 : Pyrrhosoma nymphula. Family Coenagrionidae, Suborder Zygoptera, Order Odonata. (After CHINERY in Nieuwe Insektengids, 1987.)

Figure 8 : Lestes viridis. Family Lestidae, Suborder Zygoptera, Order Odonata. 45 mm. Wing expanse 60 mm.
(After SEVERA in Thieme's Insektengids voor West- en Midden-Europa, 1977.)
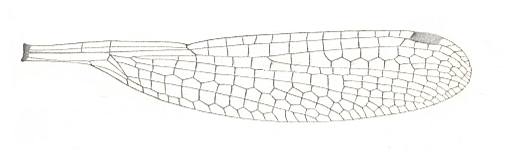
Figure 9 : Wing of Lestes rectangularis. Family Lestidae, Suborder Zygoptera, Order Odonata. Recent.
(After COMSTOCK, 1918.)
Such wings were already present in lower Permian times :
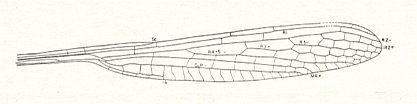
Figure 9a : Kennedya fraseri Carp. Family Kennedyidae, Suborder Protozygoptera, Order Odonata. Lower Permian of Oklahoma (USA). It is probaby a forewing. Length 40 mm, width 5.8 mm.
(After CARPENTER, 1947.)
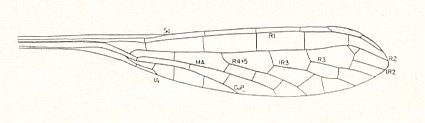
Figure 9b : Wing of Progoneura nobilis Carp. Family Kennedyidae, Suborder Protozygoptera, Order Odonata. Lower Permian of Oklahoma (USA). Length 13 mm, width 2.5 mm.
(After CARPENTER, 1947.)
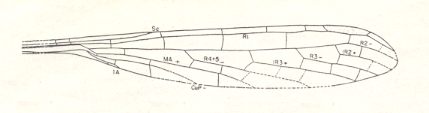
Figure 9c : Wing of Progoneura venula Carp. Family Kennedyidae, Suborder Protozygoptera, Order Odonata. Lower Permian of Oklahoma (USA). Wing length as preserved 16 mm, width 2.8 mm. Estimated whole length 17.5 mm.
(After CARPENTER, 1947.)
Many species of especially the Megasecoptera (suborders Eumegasecoptera and Protohymenoptera) (Carboniferous-Permian) show that there are gradual transitions from (the subtype) prokopepterygia (established above) to (the subtype) eukopepterygia. And indeed, there are even more or less smooth transitions from the type dolichopterygia (long-wingedness) to the type kopepterygia, and probably also from neuropterygia (through dolichopterygia) to kopepterygia.
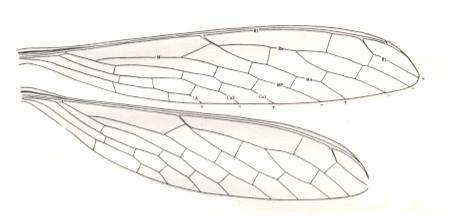
Figure 10 : Fore- and hindwings of Protohymen readi Carp. Family Protohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). Forewing length 19 mm. Width 3.5 mm. Hindwing length 17 mm. Width 4 mm. (After CARPENTER, 1933.)
The wings of this species (P. readi) and of many other Megasecoptera look much more specialized mechanically than those of the (recent) Odonata-Zygoptera. The venation of their wings shows some degree of costalization. However, they lack the specialized petiole of the Zygoptera and also the characteristic node present in zygopterous wings (the node expressing itself, among other things, by the indentation in the anterior wing-margin (see Figure 9 )).
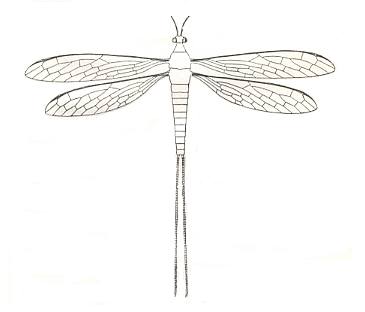
Figure 11 : Reconstruction of Protohymen readi Carp., with the addition of the cerci present in Protohymen elongatus Carp. The legs are unknown. Family Protohymenidae, Order Megasecoptera. Lower Permian of Kansas (USA). (After CARPENTER, 1933.)
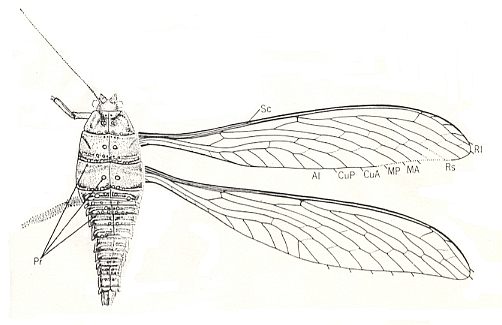
Figure 12 : Sylvohymen sibericus Kuk. Family Bardohymenidae, Order Megasecoptera. Lower Permian of Tshekarda, Ural, Russia. Forewing length 50 mm, width 9.1 mm. Pr - bases of hollow outgrowth of tergites. See for actual fossil, next Figure.
(After KUKALOVA-PECK, 1972)

Figure 12a : Sylvohymen sibericus Kuk. Reverse. The veins are secondarily colored by manganese [= a natural mineralogical event often seen in fossils]. Lower Permian of Tshekarda, Ural, Russia. Arrows point to hollow continuations of projections into the matrix.
(After KUKALOVA-PECK, 1972)
It is perhaps worthwhile to add the description (given by Kukalova-Peck, 1972) of the just figured Sylvohymen sibericus as far as wing-structure is concerned :
In the Order Megasecoptera (with the suborders Eumegasecoptera and Protohymenida) there do exist forms that are palaeopterous (not able to fold the wings over or along the abdomen at rest) and others that are neopterous (forms that are able to fold their wings over or along the abdomen at rest). In establishing t y p e s of wings we do not, for the time being, let the palaeopterous forms differ from neopterous forms although it might turn out an important difference with respect to wing-types.
Let us enumerate the families of Megasecoptera that do contain neopterous forms and those that do contain palaeopterous forms :
Asthenohymenidae - neopterous.
Diaphanopteridae - neopterous.
Prochoropteridae - neopterous.
Elmoidae - neopterous.
Martynoviidae - probably neopterous.
Aspidothoracidae - palaeopterous.
Corydaloididae - palaeopterous.
Mischopteridae - palaeopterous.
Protohymenidae - palaeopterous.
(continuing the description of eukopepterygia)
The two described subtypes of kopepterygia -- prokopepterygia and eukopepterygia -- are defined by Rohdendorf, 1949, together with a third and last subtype, neurokopepterygia, based on the mecopterous family Bittacidae (recent). The latter will be described further below.
Many years later --in 1975 -- a number of new Megasecoptera were described from the lower Permian of Moravia, Czechoslowakia, which also have markedly petiolate wings, and as such should belong to the wing-type eukopepterygia. These wings, however, are in such a high degree elongate and have such a peculiar shape that they deserve to be placed into a new subtype of kopepterygia which we will call hyperkopepterygia (hyper-kope-pterygia) :
3. Hyperkopepterygia
Wings extremely elongate, and petiolate. Posterior margin undulate. A very large number of cross-veins, and in places an archedictyon. Wing pairs homonomous. Wings very long, up to 96 mm. Lower Permian.
This subtype is so peculiar that it may, perhaps, be considered to represent an independent wing-type of its own -- caulopterygia (petiole-wingendness). The wing outline represents an extreme case of specialization of wing-shape within the Insecta. However, to emphasize the close phylogenetic relationship of these megasecopterons with the other megasecopterons definitely belonging to the type kopepterygia, we will, for the time being, consider also the former (i.e. the extremely petiolated megasecopterous wings) as still belonging to the type kopepterygia, albeit to a special subtype of it, hyperkopepterygia.
The most important family in this subtype is the Hanidae (Order Megasecoptera). KUKALOVÁ-PECK, 1975, has reconstructed the various outlines of the wings of this family :

Figure 13 : Hanidae, Order Megasecoptera. Lower Permian of Moravia, Czechoslovakia. Reconstruction of the specialized wing outline (See also next Figures).
A - Hana filia Kuk. Fore- and hindwing in natural position. Forewing, length about 84 mm, width 10.5 mm,
B - Hana lineata Kuk. Wing, length about 96 mm, width 8.3 mm.
(After KUKALOVA-PECK, 1975)
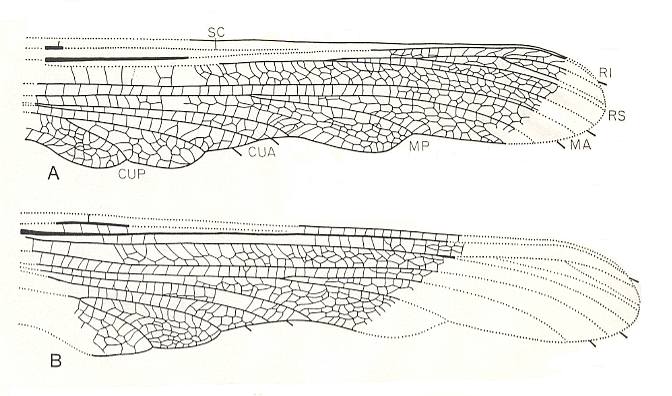
Figure 14 : Hana filia Kuk. Family Hanidae, Order Megasecoptera. Lower Permian of Moravia, Czechoslovakia.
A - Forewing fragment, length 45 mm, width 9 mm.
B - Hindwing fragment, length 38 mm, width 11 mm.
(After KUKALOVA-PECK, 1975)
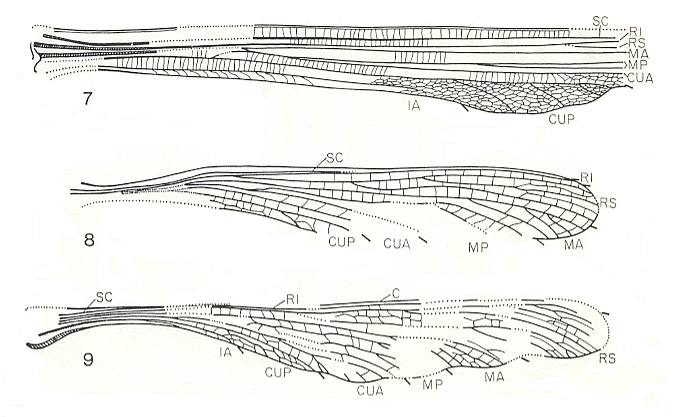
Figure 15 :
7 - Hana lineata Kuk. Family Hanidae, Megasecoptera. Wing fragment, length 58 mm., width 8.3 mm. Lower Permian of Moravia, Czechoslovakia.
8 - Arcioneura juveniles Kuk. Family Arcioneuridae, Megasecoptera. Nymphal wing, length 18.3 mm., width 2.6 mm. Lower Permian of Moravia, Czechoslovakia.
9 - Anconeura havlatai Kuk. Family Arcioneuridae, Megasecoptera. Wing, length 40 mm., width 5.7 mm. Lower Permian of Moravia, Czechoslovakia.
(After KUKALOVA-PECK, 1975)
One of the three wings just figured is actually a nymphal wing : Arcioneura juveniles, while the others are adult wings. Kukalova-Peck, in Megasecoptera from the lower Permian of Moravia, 1975, writes about nymphs of paleozoic palaeopterous insects the following :
Besides the adults, three nymphal forms are described : Cauloptera colorata, Arcioneura juveniles, Alectoneura europaea. The nymphal wings are easily distinguished from the adult wings, which are held at right angles to the body, by the characteristic bend in the proximal third of the wing length. This bend determined the oblique-lateral position of the wing pads in the living nymphs, as described by Carpenter and Richardson (1968) in the completely preserved early stage of the megasecopteran nymph of Mischoptera douglassi. However, in all isolated nymphs from Obora [in Moravia, Czechoslovakia] the angle of the bend is more obtuse than in Mischoptera douglassi. This fact together with the larger size and good sclerotization of the venation (the sclerotization being weak in douglassi) implies that the nymphal wings from Obora belonged to late, possibly the ultimate, instars.
There is no evidence as to whether or not a subimaginal stage [as we still see it in today's mayflies (Ephemeroptera)] was present in Megasecoptera. In the Palaeodictyoptera, parts of a shed cuticle have been found attached to a female of Lycocercus goldenbergi (Kukalova, 1969, p. 449, Fig. 32), including a smaller and less sclerotized ovipositor than that of the adult. It is therefore conceivable that the related Megasecoptera, which clearly descended from the Palaeodictyoptera, had adult molts.
In order to further understand the present subype -- hyperkopepterygia (hyper-paddle-wingedness) -- we add some more information about the family Hanidae (Order Megasecoptera) as new family described by Kukalova-Peck, 1975.
Wing : petiolate, broadened at about mid-wing, narrowed in CuA area. Posterior margin deeply undulated. Venation simplified. SC terminating shortly beyond mid-wing. Stems of R and M not coalesced in the basal third. RS diverging from R1 about mid-wing. MA and CuA simple, MP and CuP with few branches. MP anastomosed with CuA for a short distance. 1A [= A1] simple. Archedictyon and irregular cross-veins present. Intercalated sectors absent.
The large elongate, extremely narrow wings of Hanidae, with an intricate outline, are among the most unusual wings within the Paleoptera. The preservation of a primitive archedictyon, which in other families becomes reduced to cross-veins by Westphalian time [upper Carboniferous], is a remarkable example of mosaic evolution in paleozoic Paleoptera.
4. Neurokopepterygia
Finally, into a last subtype one must place derivatives of neuropterygia [or, perhaps, of dolichopterygia] -- scorpionflies (Mecoptera) of the family Bittacidae in which are developed wings markedly narrowed at the base, which is the most characteristic feature of kopepterygia. The venation of the wings does not possess traits of costalization, although [this venation being] fairly reduced. The legs are long, the body narrow. This subtype may be called primitive paddle-wingedness, or, equivalently, neurokopepterygia (neuro-kope-pterygia). See next Figures.
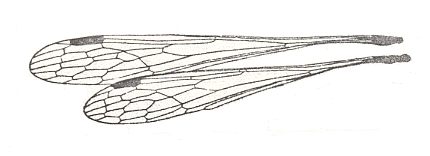
Figure 16 : Wings of Pazius gracilis Nav. Family Bittacidae, Order Mecoptera. Africa.
(After ESBEN-PETERSEN in ROHDENDORF, 1949)

Figure 17 : Bittacus italicus. Family Bittacidae, Order Mecoptera. Wing-span 35-40 mm. Europe.
(After SEVERA in Thieme's insektengids voor West- en Midden-Europa, 1977.)
Although the wings of the Order Diptera (midges, mosquitoes, flies) all belong to the type dipterygia, it contains an analogue of kopepterygia of which the wings are very similar to those of Bittacidae (Mecoptera) :

Figure 18 : Tipula oleracea. Family Tipulidae, Order Diptera. Length 15-23 mm.
The hindwings are (in all Diptera) (evolutionarily) transformed into halteres which are clearly visible in the Figure.
(After SEVERA in Thieme's insektengids voor West- en Midden-Europa, 1977.)
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for further study of the types of flight-devices in insects, Part VIII, Heteropterygia.