Much more often and more general, than is the case with antimers, the metamers or sequential parts can attain the physiological value of an independent biont. Often, isolated metamers can maintain themselves in existence, for instance the proglottids of tapeworms. Further there appear metamers as virtual bionts in the individual development of all higher animals and plants. Finally, there are large sections of the animal kingdom, for instance the Molluscs, that as actual biont almost invariably only reach the form value of a (single) metamer.
The fact that in large sections of the animal kingdom the physiological individual does not reach, as in most higher animals, the morphological value of a person, but halts at the metamer, i.e. not reaching the fifth, but only the fourth order of morphological individuality, is -- writes Haeckel, 1866 -- until now totally overlooked, or at least, because of the confusion with respect to organic individuality, not correctly recognized.
What Haeckel wrote, still stands today (2002) because Tectology has not been further developed since it was introduced in Haeckel's Generelle Morphologie der Organismen, 1866. Although Haeckel recognized the actual bionts of Molluscs as appearing as metamers only, and in addition to them some lower worms, he did not recognize the Echinoderms in the same way -- i.e. that their actual bionts appear only as metamers. We discussed this in the Section on morphological fourth-order individuality -- metamers. Neither did he recognize the Hydroid Polyps, and the solitary Anthozoans (corals and the like) to be so (which we will do shortly). By the way, because now we can recognize the actual bionts of almost all lower animals to occur as to be just metamers, the tectological theory becomes more significant : to get stuck at the metamer, i.e. not reaching the form value of a person, appears to be a sign of being just lower organisms.
Indeed the mature species form very often does not, as actual biont, elevate itself to the same tectological stage as is reached by the actual bionts of vertebrates and arthropods, but remains one stage lower, namely at the stage of the metamer. This is the case in those animals which as actual bionts are not composed of metamers, as we see in (solitary) Coelenterates, Bryozoans, Echinoderms, lower worms, most Molluscs and most Tunicates. The metamerization of the body is in fact a precondition for higher (evolutionary) development (anagenesis), because it creates a sequence of more or less identical body parts, which are relatively independent of each other in virtue of which they can become differentiated with respect to each other. In this way a division of labor can emerge. After this is accomplished (evolutionarily) integration can set in, leading to a centralized multifunctional body.
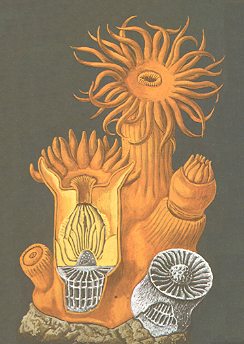
The next Figures depict some Coelenterates in which we can clearly see that the actual bionts are not articulated (segmented) along their main axis, and thus represent just metamers. The first Figure depicts, it is true a colonial Coelenterate, but the reason that we insert it here is that it clearly shows how an individual anthozoan polyp and its skeleton is constructed.
Figure 1. Starcoral, Astroides calycularis, Anthozoa, Order Madreporaria. Colony, one polyp sectioned, to the right : a calcareous skeleton. A hard basal plate (not visible in the drawing), above which later appear new transversal plates (tabulae) as the animal moves up, living on the highest plate. Not any metamerization of the polyp's body is discernable. The spaces between the successive tabulae cannot be interpreted as metamers. Along the main axis of the body often a column (columella) is developed.
( After HAEFELFINGER, H., in Grzimeks Tierleben,
Band I )

Figure 2. Solmissus albescens, a Hydromedusa (Hydrozoa). No (permanent) metamerization of the body is apparent.
( After HAEFELFINGER, H., in Grzimeks Tierleben,
Band I )
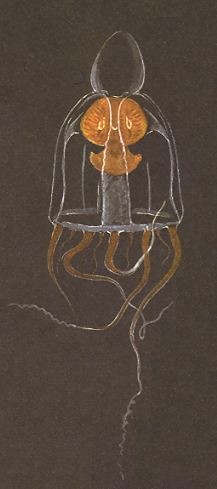
Figure 3. Leuckartiaria nobilis, a Hydromedusa (Hydrozoa). No (permanent) metamerization of the body is apparent.
( After HAEFELFINGER, H., in Grzimeks Tierleben,
Band I )
The above Figures cleary show that no metamerization (segmentation) along the main axis of body is apparent. When we study other Coelenterates, including Ctenophores (sea-gooseberries), we can conclude that all solitary living actual bionts of the Coelenterates have the form value of a s i n g l e m e t a m e r (when they occur as colonies, these must be interpreted as p l a n a r p e r s o n s, i.e. persons consisting, it is true, of metamers, but these metamers are not ordered along a line, but are placed in a (plicated) plane, i.e. they are not one-dimensionally, but two-dimensionally ordered. Although planar persons are form individuals of fifth order, their degree of individuality is lower than that of an axial person (in which the metamers are ordered along the body's main axis). So even when Coelenterates often occur as fifth-order form individuals, their individuality is much lower than the axial persons of, say, Arthropods (lobsters, insects and the like).
Not less significance than the actual, possesses the virtual individuality of metamers. We must, namely, according to the above considerations, assess all developmental stages of persons or off-shoots as virtual metamers, which are already composed of two or more antimers, but not yet of metamers.
Metamers as partial bionts are single metamers that can maintain themselves for a certain time, but are unable to develop into the actual biont.
The person or the prosopon is of all morphological individualities the most important insofar as it appears in most higher organisms as physiological individual, namely in most higher animals as actual, in most higher plants as virtual and partial biont.
The greatest importance as actual bionts the persons possess in the animal kingdom, because the mature species-representing form (i.e. the actual biont) appears in most higher animals as fifth-order form individual. This is the case in man and in all other vertebrates, in all arthropods, segmented worms (for instance earthworms and most tapeworms), further in colonial Tunicates and colonial Bryozoans.
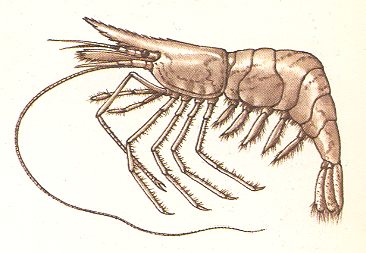
Figure 4. Pandalus montagui, a shrimp. Length up to 4 cm, sometimes more. Atlantic Ocean, North Sea, the Channel.
( After CAMPBELL, A., 1976, The Hamlyn Guide to the seashore and shallow seas of Britain and Europe. )
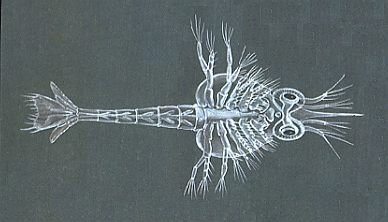
In the next Figure we depict a larva of a shrimp. Although such a larva is not yet a sexually mature biont it has already reached the tectological structure of the mature form and is as such a actual biont. It will tectologically not develop any further.
Among plants the axial person as actual biont is represented by most of the so-called simple plants belonging to the higher plants (phanerogams), and certain lower plants, i.e. off-shoots consisting of several stem members (segments), which do not have lateral off-shoots, and which have only one single (not composed) terminal flower, and which consequently do not form genuine colonies (cormi, sixth-order form individuals). Among the phanerogames only in a small number of species the actual bionts constantly appear as single persons, because most species either form branched cormi (composed of secondary asexual off-shoots), or carry a composed inflorescence (a cormus composed of sexual off-shoots). More often such single actual persons appear as exceptions in such species that usually form cormi.
While the fifth-order morphological individuals occur in most higher animals as actual bionts, they appear in most plants as virtual bionts. We then call them off-shoots or blasti. By lateral budding the person will, in the case of such plants, be multiplied resulting in a colony or cormus. Indeed in most plants the actual biont is such a colony, implying that here the person is just a virtual biont as long as it has not yet multiplied into a colony, but will do so later on. The virtual individuality of plant off-shoots is also evident by the fact that the asexual persons, i.e. the leaf off-shoots, almost always have the ability, when separated from the cormus, to develop again into a new cormus, because they produce new segmented off-shoots by lateral budding, off-shoots that remain united.
Of much less significance than the actual and the virtual, is the partial individuality to persons. The cases in which persons spontaneously detach themselves from the colony, and independently maintain their existence, without developing into a colony again, are relatively seldom. Normally we rather have cases in which persons (off-shoots), that have artificially or naturally, separated from its colony, are virtual bionts, because they develop into a new cormus (colony) again. A very good example of partial individuation of a person is given by the peculiar aquatic plant Vallisneria spiralis, in which the short-stemmed male persons (flower off-shoots) at the start of the flowering separate themselves from the colony, in order to get to the surface of the water where they, while floating, fertilize the long-stemmed female persons (Haeckel, 1866). This interesting case has some (functional) analogues in the animal world : Many medusae, the proglottids of some tapeworms and the hectocotylus of the Cephalopods. In all three cases sexually mature parts separate themselves from the actual biont, to appear as independent partial bionts. In Vallisneria they are fifth-order form individuals, in medusae and in proglottids they are fourth-order form individuals, in the hectocotylus they are of second order (Haeckel, 1866).

Because the colonies or cormi are the morphological individuals of the last and highest order, it could at first sight seem that in all cases where cormi, consisting of a multitude of persons, occur at all, these must be at the same time physiological individuals. This is in a certain sense correct. For because there is no higher morphological individuality beyond the cormus, it can never be a form individual subordinated to it.
In all species of organisms which get to the formation of colonies at all, the mature, completely developed species form, that can reproduce, is represented by the sixth-order morphological individuum. Here every completely developed biont is a genuine colony, in the sense which is established by us. This is the case in the large majority of higher plants (phanerogams) and also in many lower plants (cryptogams). In animals it is seldom the case. In a sense we find it among some tunicates (star-ascidium, see Figure 6).
Because all independent genuine colonies (cormi) are, at the time of complete maturity, eo ipso, also actual bionts, we can assess as potential or virtual bionts all those cormi, which have not yet attained complete maturity, which they will later attain, i.e. all plant cormi which have not yet bloomed. Generally all simple cormi, which are able to develop into composed cormi, can be assessed as potential (virtual) bionts. Moreover we will assess as virtual bionts all simple asexual cormi, which appear in the framework of composed cormi, for instance all branched (forked) flowerless branches, insofar as they, detached from the whole, are able to independently develop into a composed cormus again. Excluded are the sexually differentiated cormi (inflorescences) of the higher plants, which can never develop (i.e. complete themselves) into a composed cormus as actual biont.
To interpret simple cormi, that can develop into composed cormi, as virtual bionts, as was suggested above, could be problematic from a tectological viewpoint : It perhaps is more reasonable to assess a biont that has tectologically reached that order of individuality that can be observed in the fully mature form of the given species as actual biont, despite its still being immature, which here means, despite its being not yet able to reproduce.
As apparent or partial bionts we can only recognize those cormi which, when separated from actual composed cormi, cannot by growth complete themselves such as to result in a complete composed cormus again, but can only maintain themselves for a longer or shorter period after their separation. Such partial bionts are represented by the sexually differentiated cormi, i.e. the inflorescences, of higher plants, which, when artificially or accidentally separated from the main cormus, continue to bloom for a while (as also single flowers (persons) can), but which cannot complete themselves to actual bionts. They as such contrast with the asexual persons of the higher plants (leaf off-shoots), which, the other way round, possess the virtual individuality in a high degree and very commonly.
Strictly spoken, there can be no cormi as partial bionts, because such a cormus is already a sixth-order form individual. The exact status of an actual biont and a virtual biont is however still not yet once and for all determined. If indeed only the (sexually) mature stadium deserves the status of actual biont, then the above considerations are correct.
With the treatment of the p h y s i o l o g i c a l i n d i v i d u a l i t y we have completed our exposition of general organic Tectology -- heavily based on Haeckel's Generelle Morphologie, 1866 -- which is a doctrine of the general macroscopic structure of organisms (i.e. their internal form). However, before we proceed with the general Promorphology of organisms, i.e. the study of their ideal stereometric basic forms, we will summarize our Tectology in the form of some theses.
These "theses" are only provisionally. Their purpose is to stimulate further research, especially a general research as was done with regard to the macroscopical structure of organisms (Tectology), and what has done below with regard to the ideal stereometric basic forms of organisms (Promorphology).
1. All morphological properties of organisms, their anatomical as well as their developmental features, and of the anatomic properties the tectological as well as the promorphological features, are the necessary effects of mechanically operating causes. With "mechanical causes" we mean natural causes, i.e. causes wholly within Nature, where "nature" is considered to be m o n i s t i c, which means that it is ontologically homogeneous, i.e. without being mixed up with immaterial subsistent entities. The direct and intrinsic "mechanical cause" of an (individual) organism is considered to be the d y n a m i c a l l a w of the dynamical system that generated the given organism. Metaphysically this law is interpreted as the organism's E s s e n c e. The mentioned dynamical system is part of a larger context of processes and causes.
2. Every organism or living natural body is a material spatial magnitude (material unity) which as such is composed of (chemical) atoms. These atoms are considered to be the ultimate system elements of the dynamical system that generated the given organism. As such this dynamical system is conceptual in its nature : it describes the constitution of the given organism as a pattern of atomic constituents. Because Nature did not create every single organism from scratch, i.e. from a collection of atoms, the actual dynamical systems involved have as their elements already evolved complex entities [ See for more of this, the first part of our website, the documents about Organisms ( I, II, III, IV ), accessible via the back to homepage link ].
3. The atoms, composing every organism (on Earth), belong to a small number of atomic species (chemical elements). These are however combined into a host of complex chemical compounds, some of which figure as higher order elements in the building-up of the organism.
4. Of the chemical compounds that form the material substrate of every organism, many, especially the proteins, occur in a semi-liquid condition (imbibition condition). Therefore the organism grows by intussusception, not by apposition as is the case in crystals.
5. All properties of organisms are the direct or indirect effects of the chemical compounds of which they are composed, and ultimately of the atoms that compose those compounds.
6. All properties of organisms either are physiological, i.e. they relate to the functioning of the organism and of its parts, or morphological, i.e. they reflect relationships of positioning of a hierachy of parts.
7. The performances or functions of the organisms (physiological features, life activity) are, as processes only apparent, observable and describable in a series of time moments and as such object of Physiology (Biodynamics).
8. The forms (shapes) or morphs of the organisms (morphological features, life structures) are as states only apparent in a single moment of time and as such object of Morphology (Biostatics).
9. All processes, chemical and physical, that take place inside the organism, which we call life activity, proceed according to the same eternal and unchangeable laws of necessity which control all of nature, like all processes in the inorganic nature. In all these processes, insofar as they are above the quantum level, we encounter effects of efficient causes.
10. The conditions in organisms which we call life-structures, i.e. form features of organisms, are determined by the same general eternal and unchangeable laws of absolute necessity, like in all lawful forms of the inorganic nature (crystals, inorganic process structures). All these forms (shapes, symmetries), insofar as they are above the quantum level, we can and must interpret as effects of efficient causes.
11. In all organisms we find two main types of macromolecules, the nucleic acids, that can store information, and the proteins that are encoded in the latter, and function as catalysts or as construction material.
1. Every single organism as living material unit appears in the form of a uniformly closed and independent spatial magnitude, which, completely or partially, is constituted of semiliquid organic material, and which executes a uniform total of performances (life activities).
2. Every single organism, considered morphologically, and investigated as a unity only with respect to its formal individuality, appears as a morphological individual (individuum).
3. Every single organism, considered from a physiological point of view, and investigated as a life unit only with respect to its functional individuality, appears as physiological individual or biont (bion : ho bios, to on, i.e. a combination of life and being).
4. The biont or physiological individual as life unit is bound to a material substrate, which either is a single simple morphological individual, or a unified complex of two or more intimately connected simple morphological individuals.
5. Every unified complex of two or more intimately connected simple morphological individuals, which forms a natural whole, an independent form unit, must be considered as a morphological individual of second or higher order.
6. All morphological individuals, which appear in the animal kingdom, the protists and the plant kingdom as material substrate of the bionts, i.e. as carriers of the uniform life features, allow to be categorized into six subordinated stages or orders, which we, ascending from bottom to top, indicate by the following morphologically definite expressions :
7. Every single form unit of higher order is a multitude of several united form units of the previous lower orders.
8. Accordingly only the cell as morphological individual of first and lowest order is a genuine simple form individual. Cells are the ultimate tectological elements. All other morphological individuals (of second to sixth order) are always composed individuals or complexes.
1. The cell, as the only simple organic individual, is the general form-element of all organisms. It is the common base of all animals, protists and plants, without exception.
The v i r u s e s cannot be considered as living beings, their life-functions are 'borrowed' from genuine living cells.
2. Every cell contains information bearing molecules. In the great majority of cell types these molecules are themselves contained in a cell nucleus. Only nucleated cells can form genuine tissues.
3. Because the cell is the lowest tectological unit, tectology does not need to investigate the constitution of cells, i.e. it does not need to distinguish parts that compose a cell. But it nevertheless does so, thereby realizing that the results are only analogous to the tectological distinctions made above the level of the cell.
1. All morphological and physiological properties and features of the composed organic individuals originate from the simple individuals, the cells, and ultimately from the constituent molecules and atoms. It must however be realized that the mentioned properties and features of the composed organic individuals are normally not as such present in the more or less ultimate (i.e. lower-level) constituents (cells and their constituents). They emerge from a complicated interaction of those lower-level constituents and only appear at higher scale levels (structural levels).
2. The composition of the complex individuals as an aggregation of simple individuals proceeds according to the same laws in all three organic domains (animals, protists and plants).
3. The Organ (in a purely morphological sense, as second-order morphological individual) is a complex of two or more united cells.
4. The Antimer or the homotypic part (counterpart) is a complex of two or more united organs.
5. The Metamer or homodynamic part (sequential part) is a complex of two or more united antimers.
6. The Person or Prosopon is a complex of two or more united metamers.
7. The Colony or Cormus is a complex of two or more united persons.
1. Every existing biological species of organisms is composed of physiological individuals, which under approximately equal conditions or at least under similar existential conditions, pass, in their individual development, through an equal or at least similar form sequence. This means that every member of the given species passes through a specific series of stadia. In the first part of our website we have called the total series of stages that are passed through, the h i s t o r i c i n d i v i d u a l, in contradistinction to the h e r e - a n d - n o w i n d i v i d u a l, which is the individual during an infinitely small time span. The latter corresponds to the morphological individual if the consideration is purely morphological. The physiological individual is neither of them, but is the individual seen at some point in time but considered to be changeable (thus in fact seen during a longer or shorter period, long enough for (functional) processes to be apparent).
2. For every biological species the stage or order of morphological individuality, that is attained by the mature state, is constant or typical, and will be indicated by the term a c t u a l b i o n t. It is the actual physiological individual.
3. Only the unicellulars can be considered as to be genuine simple organismic species, i.e. those species, in which the actual biont as wel as all its developmental stages have the form value of a single cell (either a non-nucleate, or a nucleate cell).
4. All organismic species, which are, as actual bionts, composed of two or more cells, and accordingly are second- to sixth-order form individuals, can be considered as composed organismic species. There is however a fundamental distiction to be made among those composed species : Genuine cell tissues can only be formed from nucleate cells. Organisms consisting of such true tissues we will call h i s t o n e s. The others are just cell fusions.
5. All organisms which as actual bionts appear as second- to sixth-order form individuals, i.e. all composed organismic species, pass, in their individual development, through all the previous lower orders of individuality, from the first onwards.
6. As long as the biont finds itself at a morphological individuality order which is lower than that order which it is supposed to reach as actual biont, it is either a virtual or an partial biont.
7. The physiological individual must be evaluated as a virtual or potential biont if it is able to develop into the higher order of morphological individuality, that is typical for the actual biont of the species.
8. On the other hand the physiological individual must be assessed as partial or apparent biont if it is, to be sure, able to exist for a longer or shorter period as independent life unit, but at the same time not able to develop itself into that morphological individuality order which is typical for the actual biont of its species.
9. The actual, as wel as the virtual and partial bionts can have each of the six morphological orders of individuality as material substrate.
10. All physiological individuals, all the same which of the six morphological individuality orders their material substrate assumes, are with respect to all their performances and form relationships reducible to the first-order morphological individuals, the cells, the "elementary organisms", because every biont either itself is a single cell or an aggregate of several cells. This reducibility is a reducibility only in principle, i.e. an ontological reducibility. The actual reduction of a multicellular organism's performances and form relationships, as is attempted in Natural Science, is hardly possible : As such we have to do with an epistemological irreducibility.
We spoke about "an aggregate of several cells". This is, in the case of multicellular organisms far from just an aggregate, i.e. far from just a random aggregate. The cells are organized in a very subtle way, and because of this emergent properties and features will appear at the level of the whole (multicellular) organism, which are not as such visible at the level of the cells. This is comparable with a novel : The story and the plot are not visible at the level of the letters which compose the book. They are only visible at a higher level. Nevertheless the story and plot are wholly contained within those letters, i.e. nothing of them comes from outside these letters.
1. The structure (the internal form) of an organism is the total of spatial relationships between the single constituents of that organism and the relationships of those constituents to the whole.
2. In unicellular organisms which, as actual bionts, remain at the first order of morphological individuality, their structure is determined by the cell constituents, i.e. by the spatial relationships between those constituents and the relationships of the latter to the whole.
3. In multicellular organisms on the other hand, which as actual bionts reach the second- or higher-order morphological individuality, the structure is determined by the existing spatial relationships of the constituent morphological individuals of all lower orders and ultimately those of the first order among each other, and by their relationships to the whole.
4. The different degrees of morphological perfection, which are shown by the different organismic species, are partially determined by their tectological features, partially by their promorphological features, and thus neither determined by their structure alone, nor by their basic form alone (The basic form of organisms will be dealt with in the next series of documents, together embodying the science of Promorphology or Doctrine of the Ideal Stereometric Basic Forms of Organims).
5. The different degrees of the perfection of the organisms are, insofar as they immediately refer to structure, determined by several different tectological factors, which are essentially grounded in the mutual relationship of the aggregated morphological individuals of different order among each other, and in their relationship to the whole.
6. The perfection of the organism is proportional to the height of the morphological individuality order to which it elevates itself, i.e. the organism is higher the more subordinated orders of individuality it encloses.
The latter statement we can read in Haeckel's Generelle Morphologie (p. 371). As such it maybe too simplistic. The "perfection" or "height" of an organism is a very vague concept, which consequently is hard to define. At first we should distinguish between morphological perfection and physiological perfection (the latter relates to internal processes, but also to the organism's behavior). If we confine our assessment to morphological perfection (as in 4.), then Haeckel's statement is more convincing.
7. The organism is, when it is composed of similar cells, the more perfect, the greater the number of constituent cells.
Of course there are unicellular organisms that are highly complex, and as such very "perfect" as can be seen in Ciliates and in Radiolaria. See the next Figure.
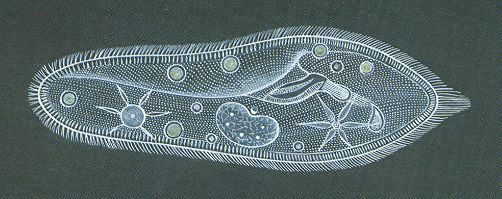
Such unicellular animals are not only more complex than a fusion of simple cells, like we see in colonial Cyanophytes, the cells of which do not have a nucleus, but probably also more complex than Sponges or colonial Diatomeans. Nevertheless we should realize that the formation of a multicellular body, even when the constituent cells are moderately simple, like a Diatomean cell, is the very condition of developing upwards (anagenesis), because from then on division of labor can set in : the constituent cells develop specialisms with respect to certain tasks, without affecting the rest of the multicellular body.
8. The organism is, when it is composed of dissimilar cells, the more perfect the more dissimilar the composing cells are, because here we have a sign of division of labor, which effects a higher performance of the whole organism.
9. Every morphological individuality of whatever order is the more perfect the more dissimilar the majority of the next lower-order individuals are which compose it, i.e. the greater the polymorphism of that next lower order is.
10. The organism is the more perfect the more the similar individualities composing it are dependent upon each other and upon the whole, and consequently the more the whole organism is centralized and controlling all subordinated individualities.
11. On the other hand every single form individual of whatever order is the more perfect the more independent it is of its coordinated fellow individuals (the other form individuals of the same order), and the more independent it at the same time is of the superordinated whole (Autonomy of the parts).
12. The organism is the more perfect the more intense the division of labor and the degree of interaction is among all subordinated individualities that compose it, the greater consequently is the differentiation and centralization of the whole organism.
1. The first-order form individuals, the cells, are, first of all more perfect when they contain a nucleus. In addition to this they generally are the more perfect the greater the number of organelles is, and the greater the division of labor is among these organelles, the more dependent those organelles are upon each other and upon the whole cell, and the more the whole cell is centralized and the more this cell is independent of the superordinated organ when present.
2. The second-order form individuals, the organs, are generally the more perfect, the greater the number of their constituent cells, the more diverse these cells are, the more dependent they are upon each other and upon the whole, and the more the whole organ is centralized and the more independent it is of the superordinated antimer if present.
3. The third-order form individuals or antimers are generally the more perfect the greater the number of organs constituting them is, the more diverse the histological composition of those organs is and consequently also their physiological function (and thus resulting in a greater versatility of the antimer's physiological function), the more dependent the organs thus are upon each other and upon the whole antimer, and the more the whole antimer is centralized and independent of the superordinated metamer (if present).
About the relation of antimers and metamers we can -- with Haeckel -- append the following (the remark still stands today) :
The many tectological difficulties which, in higher organisms, pop up because of the fact that the different morphological individualities are very diversely intermingled and entangled, and connected in diverse ways, are already partially discussed earlier. Very easily confusions can arise because of the mutual entanglement of antimers and metamers, as also in organs (the entanglement) of paramers and epimers, where similar complications reign. Partially this is the cause that the homotypic and homodynamic relationships until now have received little attention (as is the case for the whole of Tectology for that matter) -- only, as far as I know they received attention in BRONN (a short time before Haeckel's Generelle Morphologie) and Haeckel himself in the just mentioned work . Concerning the important tectological relationship between the antimers and the metamers, we want to emphasize explicitly that we in the tectology of persons, insofar as their composition out of antimers is concerned, always determine the number of antimers in the same way as we determine that number in metamers, and independent of the number the latter, because this is without influence. Strictly speaking we should attribute to a vertebrate, which consists of fourty metamers, eighty antimers because each metamer consists of two antimers. We however in this case attribute to the whole person only two antimers, because the same homotypic composition is repeated in all homodynamic segments. In the same way we do not attribute to a five-fold flower having six five-fold leaf rings (metamers, one consisting of sepals, the other of petals, etc) thirty antimers, but five antimers. Attention to this way of assessing is especially of great significance for a correct understanding of basic forms (in Promorphology).
4. The fourth-order form individuals, the metamers or sequentional parts (homodynamic parts) generally are the more perfect the more differentiated, the more dissimilar, their homotypic, organological and histological composition is (i.e. their composition out of antimers, organs and cells), and consequently the more versatile their (i.e. the metamer's) physiological function is, thus the more dependent the constituent cells, organs and antimers upon each other and upon the whole metamer are, and the more the whole metamer is centralized and independent of the superordinated person if present.
5. The fifth-order form individuals, the persons, prosopa or off-shoots, generally are the more perfect the more differentiated, the more dissimilar, their homodynamic, homotypic, organological and histological composition is, and consequently also the more versatile their (i.e. the person's) physiological function is, thus the more dependent the constituent cells, organs, antimers and metamers are upon each other and upon the whole person, and the more strongly the whole person is centralized and independent of the superordinated cormus if present.
6. The sixth-order form individuals, the colonies or cormi, generally are the more perfect the more differentiated, the more dissimilar, their prosopological, homodynamic, homotypic, organological and histological composition is, and consequently also the more versatile their (i.e. the colony's) physiological function is, thus the more dependent the constituent cells, organs, antimers, metamers and persons (prosopa, off-shoots) are upon each other and upon the whole colony, and thus the more the whole colony is centralized.
To begin the study of Promorphology (Stereometry of Organisms) click HERE
back to homepage
back to retrospect and continuation page
back to Internal Structure of 3-D Crystals
back to The Shapes of 3-D Crystals
back to The Thermodynamics of Crystals
back to Introduction to Tectology
back to Tectology, cells, organs, antimers
back to Tectology, persons and colonies
back to Tectology, Physiological Individuality, Cells, Organs, Antimers