When we look to animals like scorpions, insects, shrimps, and especially to earthworms, we see that their body is articulated, i.e. their body is composed of repeating form units. In many cases these units are almost identical to each other, like in earthworms, but in most other cases they become different, i.e. a strong division of labor among these units has been developed resulting in a different morphology, like we see in many insects (for instance in our common housefly). These repeating units are morphologically more or less clearly delimited which means that we can assess them as morphological individuals. We call them m e t a m e r s or s e q u e n t i a l p a r t s, because they are positioned b e h i n d each other (not next to each other).
The articulation as described above can, in many cases be repeated in other body parts (i.e. a repetition like in the case of metamers can happen again in other body parts), for example in the limbs of the animal. This we can clearly see in all Arthropods (which got their name as a result of this phenomenon). These repetitions are, to be sure, morphological units, but each one of them does by far not represent the animal's essential morpholgy. Only certain structures are repeated. Moreover the repetition generally is not along the body's main axis but along a cross axis. We have already discussed these repeating parts (parts, repeating after the fashion of metamers but which nevertheless are not metamers themselves) earlier, namely at the end of the section on organs and called them e p i m e r s.
Before we will extensively discuss (as always guided by the relevant sections in Haeckel's Generelle Morphologie) the metamers and epimers (and their relation to antimers and paramers) we will first define them. The ensuing discussion will then be more clear when we have these definitions in mind.
Definition of Metamers.
H o m o d y n a m i c p a r t s or M e t a m e r s ( s e q u e n t i a l p a r t s ) are form segments that originate by the articulation or division (as a result of multiplication (repetition) of form units) of the trunk of an animal, or of any off-shoot of a plant, i.e. an articulation of the body along its main axis. Such a metamer (potentially) more or less represents all the essential body parts and organs. The metamers, almost always lying along the body's main axis, together make up a person or fifth-order form individual, while a single metamer (free-living or not) is a fourth-order form individual.
Definition of Epimers.
H o m o n y m o u s p a r t s in our sense, or e p i m e r s, are form segments that lie behind each other, and which originate by an articulation or division (as a result of multiplication (repetition) of form units) of a part of the body that generally does not lie in the body's main axis but in one of its cross axes. In no case can the epimer represent the whole of essential body parts and organs. If there is a repetion of certain units along the body's main axis we can still distinguish between their being metamers or epimers. A metamer more or less represents all the body's essential parts and organs, while an epimer does not, it represents only a small part of the whole type set of body parts and organs. An epimer consequently is, as a form individual, subordinated either to a third-order form individual (antimer), or to a second-order form individual (organ), or (but now analogously) to a first-order form individual (cell).
Discussion of Antimers, Paramers, Metamers and Epimers (Generelle Morphologie, pp. 312).
We will now discuss counterparts (antimers and paramers) and sequential parts (metamers and epimers), in order to fully understand what they represent, and also to better understand of what Tectology is all about.
In this discussion expressions like homonymous parts play a role. Their initial meaning as they have it in the ensuing discussion will be clear, their FINAL meaning, however, is only determined in the above definitions.
More attention, as to the antimers, is devoted by researchers to the metamers, although metamers usually come together not in so constant a number and in so delimited a form as form-determining units, as it is the case in antimers. But because metamers more often than antimers play the role of physiological individuals, and moreover are conspicuously expressed in the axis of higher plants as stem members, in the trunk of Vertebrates (internally as vertebrate segments) and of articulate animals (externally as rings or zonites), they had received some closer attention in various considerations concerning some phyla of organisms.
Establishing the relationships between metamers and antimers is attempted only by BRONN (before Haeckel did). In his morphological studies he expresses himself about both essential form units in the following way :
"One has called h o m o l o g y the complete correspondence of parts of different plants and animals with repect to their relative original position as to other parts, without any consideration of their form. So the front legs of all Vertebrates are homologous, be they fins, wings, digging devises, walking legs or arms with hands. Earlier (1850) we had called parts of one and the same animal h o m o n o m o u s ( = following the same laws), which are built according to the same way or to one law or plan, but we now must, serving clarity, discriminate more precisely. We call such organs h o m o t y p i c, which occupy, according to the basic form of the type or systemic group to which they belong, a wholly identical place within the individual, and which because of this are also definitely determined as to their number. They accordingly will, in their position with respect to the main axis of the organism, equal each other so much, that they correspond to the two poles of one or another transverse axis or its two equal radii. So in every higher animal there will be only two homotypic legs, fingers, ribs, teeth, and in radiate animals [there will be] only four, five, six homotypic radii, arms, antennae, radial ledges and the like, present. We choose on the other hand the expression h o m o n y m o u s ( = bearing the same name) to indicate such form constituents of one and the same animal or plant, which, also in daily life, are called by one and the same name, and which are built according to one plan, but nevertheless always deviate from each other with respect to position such that they lie along a main or radial axis b e h i n d each other, or, in a pole of a transverse axis, lie n e x t to each other. To the f i r s t belong all rings of an articulated animal, all successive feet, teeth and rib pairs of an animal, the successive pairs or alternating individuals of members, feather branches, pedicells and tendrils on the arm of a crinoid etc. To the l a t t e r belong all fingers and toes of a hand and of a foot, and also the successively different cycli of radiate ledges in a coral calyx etc. The homotypic basic number [as was discussed above by BRONN] in radiate animals is 6, 5, 4, 3 , in higher animals 2. For the homonymous organs on the other hand there is no other basic number as the pair or unity."The last sentence seems to me a bit cryptic. BRONN comprises under the concept of homonymous parts, what we have called metamers, epimers and paramers. The metamers and epimers do not come in such constant numbers as is the case in homotypical parts, but the paramers behave, according to me, in the same way as with respect to their numbers as do the homotypic parts (antimers).
Let us first of all determine the difference between the concepts of homology, analogy, and homonymy.
The homonymy compares two corresponding parts of one and the same organism, while the homology compares two corresponding parts of two different organisms.
Although the difference between h o m o l o g y and a n a l o g y is so clear and definite a difference, both expressions nevertheless are often confused (Haeckel, 1866) and in this way cause obscurity. So it is not superfluous to indicate that both concepts only cohere with each other by the common tertium ( = third ) of c o m p a r i s o n, i.e. that they are two different sorts of comparison. The a n a l o g y or the p h y s i o l o g i c a l c o m p a r i s o n, only can refer to the corresponding f u n c t i o n of two parts, while the h o m o l o g y or m o r p h o l o g i c a l c o m p a r i s o n can only refer to the corresponding f o r m of two parts. A n a l o g o u s are for example the gills of fishes and the lungs of mammals, while the swim bladder of the fishes and the lungs of mammals are h o m o l o g o u s.
BRONN's concept of homonymic should be further determined and divided into several different concepts. In the sense of BRONN, as given above, the concept of homonymic generally means the relationship between two similar parts which "are built according to one plan, but nevertheless always deviate from each other with respect to position such that they lie along a main or radial axis b e h i n d each other, or, in a pole of a transverse axis, lie n e x t to each other." In this definition already essential differences of parts commonly named by the term "homonymous", are indicated. For it is clear that we may not, just like that, compare such sequential or next-to parts, without indicating whether they lie behind each other or next to each other in a main axis or in a tranverse axis. We should not, for instance, consider the single segments of the trunk of an articulated animal (segments along its main axis) as being directly equivalent to those of its limbs (segments along cross axes).
In order to get to grips with these difficult but important relationships of form composition, it is necessary to anticipate some matters from Promorphology (which comes after Tectology). These matters concern the indication of the most essential axes.
In the so-called bilateral-symmetrical animals (Vertebrates, articulate animals and Molluscs), the basic form of which we will later determine as d i p l e u r a l, we distinguish three body axes perpendicular to each other, corresponding to the three spatial directions (i.e. the three dimensions of space). The first or longitudinal axis (axis longitudinalis), which usually goes from front to back, will be interpreted as m a i n a x i s. One of its poles is the mouth pole (Polus oralis, or Polus peristomii), the other pole is the counter mouth pole (Polus aboralis, or Polus antistomii). The second axis, which goes from back to belly we call the a x i s o f t h i c k n e s s (Axis dorsoventralis). One of its poles is the back pole (Polus dorsalis), its other pole is the belly pole (Polus ventralis). The third axis, which goes from right to left, is the a x i s o f w i d t h (Axis lateralis). One of its poles is the right pole (Polus dexter), its other is the left pole (Polus sinister).
In the radiate animals, which are also often indicated as regular animals, and also in most sexual individuals (flower off-shoots) of the higher plants, the mentioned three axes can in some cases positively be distinguished (if they belong to the form group of the Centrepipeda), in other cases not. In the latter cases (in the wholly regular radiate animals and flowers) only the length dimension is determined by an axis and this main axis is the longitudinal axis, which goes from mouth pole to counter mouth pole, for example in a sea-urchin from its mouth to its anus, in a medusa from its mouth to the vaulting of its bell-shaped body, in the regular funnel-formed flower from its opening to its base. The other axes, which are determined by the number of radii, and which run along the midlines of the radii, and which meet in a point on the main axis, five in Echinoderms (Sea-urgins, starfishes and the like) and most dicotyledone flowers, four in most medusa and many dicotyledones, three in most monocotyledones, we call c r o s s a x e s (Stauri).
One of the most common features, present in the organism of higher animals, and related to its structural composition out of subordinated parts, is its a r t i c u l a t i o n or s e g m e n t a t i o n, i.e. the formation of parts that lie behind each other along an axis, parts that essentially repeat the same number of organs in the same or similar positions, composition, form, etc. This articulation which is most clearly expressed in Vertebrates and articulate animals (while it is totally absent in Molluscs (snails, bivalves, and the like)), can be present with respect to the trunk (i.e. a segmentation along the body's main axis) as well as with respect to lateral appendages, which are repetitions either along the the lateral axis (in arthropods) or along the cross axes (in radiate animals, for example Echinoderms) of the body. In both cases the segments are called homonymous parts. Precisely the same general morphological status as being possessed by the segments or zonites of the trunk of Vertebrates and articulate animals (Articulata), can be assigned to the single stem members in higher plants. Also these are repetitions of homonymous parts along the main axis. And also we do not oppose to equate the articulation, which is expressed in lateral parts (leaf organs) of higher plants, for instance in feathered leaves, with the articulation of lateral appendages (limbs, etc.) of Vertebrate and articulate animals.
In order to correctly determine the ranking of subordinated form groups out of which the whole body of those segmented animals and plants is composed, it is, however, absolutely necessary to discriminate between these two cases. We will (following Haeckel) accordingly reserve the term h o m o n y m i c (introduced by BRONN) to indicate the relationship of segments that lie behind each other, and which (segments) originate by articulation of a lateral part not lying in the main body axis, a lateral part that thus corresponds to a lateral axis or cross axis of the (whole) body. And if we nevertheless have to do with an articulation in the direction of the main body axis, then the resulting segments are (also) called homonymous if they are repetitions of only a small part of the body features, or in other words, if they are parts subordinated to one or another o r g a n (in its morphological sense) instead of being directly subordinated to a person, i.e. to a fifth-order individual. The mutual relationship of those segments on the other hand, that originate by the articulation of the trunk itself (and thus an articulation along the main body axis), will be indicated by the term h o m o d y n a m i c.
Further we will call (for short) the segments along the body's main axis or the homodynamic parts, m e t a m e r s, the segments along the cross axes or lateral axis or homonymous parts, and also all other parts lying behind each other but not repeating all the essential features of the body, (we will call) e p i m e r s.
H o m o n y m o u s p a r t s in our sense, or e p i m e r s are for instance the limb segments of the Vertebrates and Arthropods, further we see homonymous parts originated by the feathering of leaves of plants.
H o m o d y n a m i c p a r t s or m e t a m e r s on the other hand, are the single segments of the trunk in Verebrates (these segments are only internally detectable), and also those in articulate animals. With respect to the latter these segments (metamers) are already highly differentiated among each other (heteronomous) in Arthropods, while they are still essentially the same, or at least very similar (homonomous) in (articulated) worms.
In higher plants we see the metamers appear as the stem members.
The metamers are accordingly subordinated parts (segments), directly of a fifth-order form individual, the epimers on the other hand, are subordinated parts (segments), directly of a first-order form individual (but here in an analogous way) or, directly of a second- or third-order form individual.
The metamers or homodynamic body segments have as articulations of the main axis of course a much higher morphological status than the epimers have, which only appear as articulations of cross axes or of a lateral axis, or, if not so, only repeat a small part of the body features. Furthermore we will later see that the epimers never, or only seldom are capable of a physiological individuation in the animal kingdom, while the metamers are very capable of such an individuation. The metamers are in the lower forms of animals in which they appear, just multiplications of the specific form of the given species, repetitions which originally are so independent that they easily become separated from each other and that then each single metamer more or less completely represents that species form. The epimers on the other hand, will never in the same way represent the species form, because they are not repetitions of the whole organism, but only multiplications of some lateral parts of it, i.e. of organs of different orders. The epimers relate to the metamers analogously as do the paramers to the antimers.
The next two Figures diagrammatically illustrate a possible relation between metamers and epimers (and of antimers and paramers).
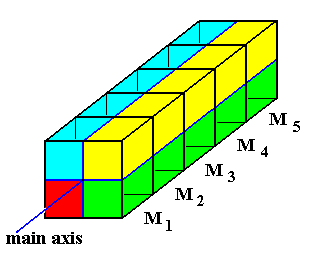
Figure 1. Diagram explaining the relationship between antimers (red, green, blue, yellow) and metamers ( M1, M2, M3, M4, M5 ). The m e t a m e r s are repetitions along the body's main axis. The a n t i m e r s are repetitions about the body's main axis.
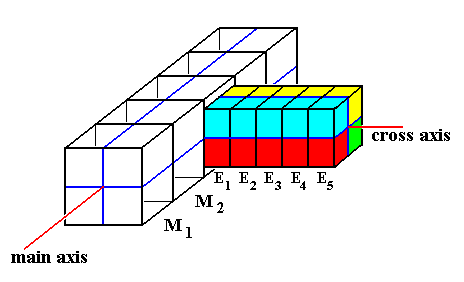
Figure 2. Diagram explaining the relationship between body trunk, paramers (red, green, blue, yellow) and epimers ( E1, E2, E3, E4, E5 ). The e p i m e r s are repetitions along a cross axis. The p a r a m e r s are repetitions about a cross axis.
The next Figure illustrates several subordinated types of epimers in a leaf of an umbelliferous plant.

Figure 3. A feathered leaf of an umbelliferous plant, showing two sets of epimers. The stem onto which the leaf is attached is also visible. The Figure below depicts this diagrammatically.
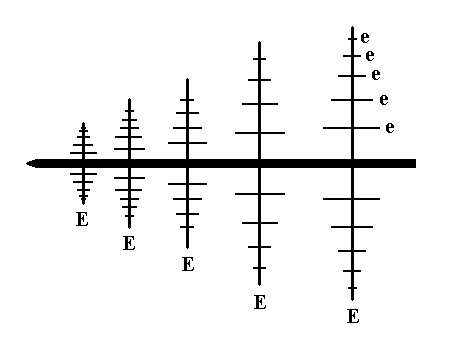
In the Figures 3 and 4 we see two subordinated groups (series) of epimers (sequential parts), which are leaflets composing a leaf belonging to an umbelliferous plant. In figure 4 these are indicated by the letters E and e. The leaf's morphology displays the beginning of the generation of a f r a c t a l : The same form appears at smaller and smaller scale levels. In our umbelliferous leaf we see the first two levels (a genuine fractal has an infinity of such scaling levels). The same segmentation (articulation) has occurred (only) twice here.
The Tectology of Echinoderms
The (adult) E c h i n o d e r m s (starfishes, brittle-stars, sea-urchins, sea-cucumbers and sea-lilies) are very instructive for a general understanding of Tectology (and also for Promorphology for that matter). And they are especially relevant in the present section that is dealing with fourth-order form individuals or m e t a m e r s, because, as we will show, their physiological individuality never reaches that of a person (fifth-order form individual). Their physiological individuals (bionts) are always represented by just a single metamer (fourth-order form individual), like in Mollusks. Antimers, paramers and epimers are clearly expressed in them. So it is probably a good idea to dwell especially here a little further on these remarkable animals, that defy being neatly placed as descendants into an ancestral tree. Let us begin with their promorphology, i.e. their axial relationships.
Some promorphological features of Echinoderms.
The a x i a l s y s t e m of individual adult Echinoderms, i.e. their p r o m o r p h o l o g y, can be described as follows :
The body's m a i n a x i s is heteropolar (unequipolar) which means that their poles represent different non-equivalent parts of the body. The one pole of the main axis is called the mouth pole (Polus oralis) while the other pole is called the counter mouth pole (Polus aboralis).
Perpendicular to the main axis are five semiradial transverse axes, semiradial c r o s s a x e s, each one of which is heteropolar too. Each semiradial cross axis consists of a radial half and an interradial half. These cross axes can either be equal to each other, as is the case in regular Echinoderms, or unequal, as in irregular Echinoderms. All this implies that their s t e r e o m e t r i c b a s i c f o r m o r p r o m o r p h o n t can be expressed by either a regular five-fold pyramid, representing the basic form of regular Echinoderms, or half a ten-fold amphitect pyramid (An amphitect pyramid is a pyramid that is more ore less flattened, and thus having a base that is an elongated polygon -- About amphitect pyramids see our exposition of Promorphology, where they are discussed in many places, for example in the document concerning the Stauraxonia).
The next two Figures depict the promorphological features of regular Echinoderms.
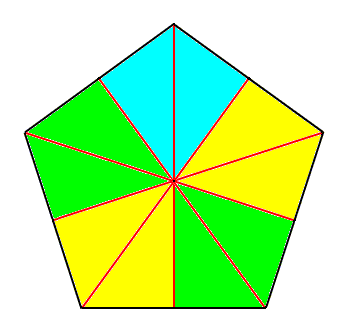
Figure 5. Base of a regular 5-fold pyramid (such a pyramid represents the basic form of the Pentactinota, which is a promorphological category). The 5 semiradial cross axes are indicated. The five antimers are indicated by coloration. This basic form is materialized in regular Echinoderms. The five radii of such an Echinoderm correspond to the five radial halves of the (five) semiradial cross axes. The radial halves of the cross axes end up in a corner of the basal polygon, while the interradial halves end up at the mid-point of a side of that polygon.
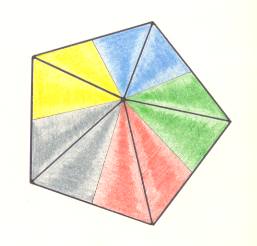
Figure 6. Slightly oblique top view of a five-fold regular pyramid as the basic form of the Pentactinota. The five antimers are indicated by colors. The main axis descends from the tip of the pyramid and ends up perpendicular to the base of the pyramid. The tip of the pyramid represents the aboral pole of the main axis, while the base represents its oral pole. The five antimers of the pyramid could for instance correspond to the five arms of a starfish.
In irregular Echinoderms those promorphological features are more complicated (involving half amphitect pyramids) but need not be discussed here.
General characterization of Echinoderms
The familiar starfish and sea-urchins which are so common in shallow waters are representative of an entirely marine phylum, the Echinodermata, that occurs since Cambrium times (c.a. 600 million years ago), which stands apart zoologically from nearly all other invertebrate groups. Echinoderms of all kinds have internal m e s o d e r m a l s k e l e t o n s of porous calcite plates [Calcite is a mineral], which are normally spiny and covered outside and in by a thin protoplasmic skin. Normally the skeletons have a five-rayed or p e n t a m e r a l s y m m e t r y, though in some fossil groups this is not so and in some modern and fossil sea-urchins a bilateral symmetry is superimposed upon the radial plan. Another important feature of Echinoderms is the w a t e r - v a s c u l a r s y s t e m : a complex internal apparatus of tubes and bladders containing fluid. This has extensions which emerge through the skeleton to the outside as the t u b e - f e e t or p o d i a (sometimes also called tentacles). Tube-feet have various functions, especially locomotion, respiration and feeding. They may be considered as all-purpose organs used by animals living inside a calcite box for manipulating the environment. Echinoderms are strictly marine animals (stenohaline). [CLARKSON, E., 1980, Invertebrate palaeontology and evolution]
All Echinoderms possess an (internal) ambulacral system, i.e. the above mentioned water-vascular system. It consists of a circum-oral ring from which five radial water vessels extend through the radii of the animal. From the radial water vessels there arise, at intervals, paired lateral tubes each leading to a tube-foot and associated apparatus. The circum-oral ring lies -- as the name already indicates -- close to the mouth of the animal.
In Echinoderms as living animals one can distinguish four h a b i t u s t y p e s (See Figur 7) :
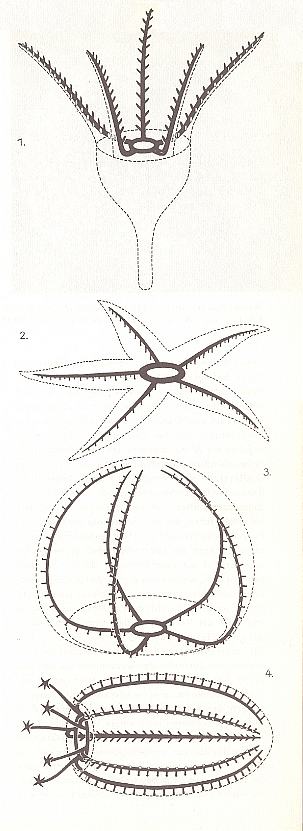
Figure 7. The four habitus types of Echinoderms, physiologically oriented.
1. Sea-lilies.
2. Starfishes.
3. Sea-urchins.
4. Sea-cucumbers.
Black : The water-vascular or ambulacral system.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
Tectology of Echinoderms
We are now ready to study the Tectology of Echinoderms.
We'll begin with starfishes (Asteroida). Their morphology and anatomy will be evident from the next Figure.
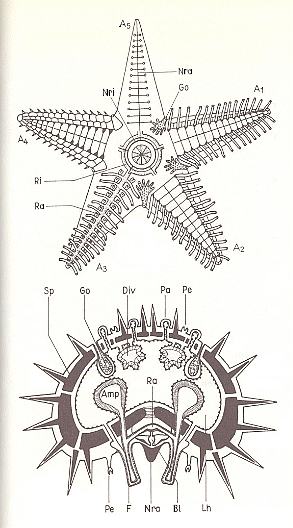
Figure 8. Organizational scheme (top), and tranverse section (bottom) through an arm, of a starfish.
A1 -- A5 Arms (A1 and A2 with extended tube-feet, A3 with water-vascular system, A4 with skeletal plates, A5 with nervous system).
Circum-oral ring (Ri) and radial canal (Ra) with tube-feet (F) and ampullae (Amp) of the water-vascular system.
Neural ring (Nri) and radial nerve (Nra), Go gonad, Bl blood lacuna, Div part of the gut, Pa papulae, Pe pedicellariae, Sp skelettal plates with spines, Lh body cavity.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
In (adult) Asteroids (starfishes) the radii are stretched out laterally as compared with Echinoids (sea-urchins) and Holothurians (sea-cucumbers). The segmentations of the arms are sequential repetitions along cross axes of the body and so are not metamers but epimers. In addition to that the repetition does not involve all or almost all essential parts of the body. There is no repetition along the body's main axis, so there are no genuine metamers. This implies that an asteroid is not a person : A physiological individual of an asteroid is not a fifth-order form individual, but a fourth-order form individual, i.e. just a single metamer. The five radii of an Asteroid do repeat all essential body parts and thus are genuine antimers (not paramers), as the next Figure diagrammatically shows.
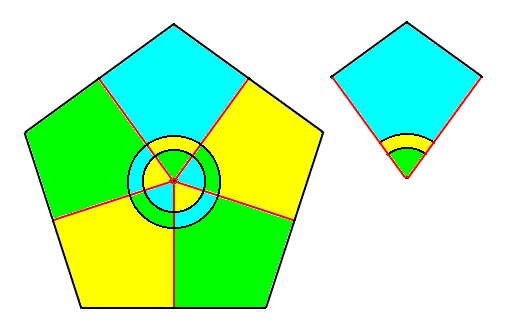
Figure 8a. Diagram of the five radii of a regular Echinoderm. The axial organs (in a geometric sense) are indicated schematically by concentric discs. Each radius contains one fifth of the whole animal, including one fifth of the axial organs, as can also be seen in the isolated antimer (right image). So such a radius contains all essential parts of the animal, and these parts are repeated five times around the body's main axis. These radii are therefore true antimers.
Even in regular Echinoderms we will encounter e x c e n t r i c elements (for example the location of the madreporite in regular sea-urchins (See Figure 12)). These can be seen as physiological necessities. Tectology, however, investigates the g e n e r a l c o n s t r u c t i o n p l a n of organisms, and abstracts from small individual isolated deviations from such a plan, i.e. it does not consider such minor deviations.
E p i m e r s normally are sequential repetitions along a cross axis of the body (at any place within a cross plane), but can also represent repetitions along the main axis of the body (or along a line parallel to the main axis). This we see clearly for example in (adult) Holothurians (sea-cucumbers). The reason that in this case they nevertheless are not metamers is given by the fact that not (even in principle) all of the essential body parts are repeated : only a small fraction of body parts is repeated. In Holothurians these are the tube-feet and their associated ampullae, nerve elements, etc. In the body as a whole nothing else is repeated along its main axis, i.e. the axial organs (in a geometric sense) are not segmented along the body's main axis. The sequential repetitions p a r a l l e l t o t h e m a i n a x i s of the holothurian (but also in the echinoid's) body is accomplished by the fact that the radial parts are (permanently) bent up and come to lie parallel to the body's main axis.
The Holothurians do not have a coherent skeleton, but possesses, if any, only isolated spicules.
Before we depict the organizational features of Holothurians, it is perhaps instructive to divert for a moment to an animal group in which the segmentation a l o n g t h e m a i n a x i s o f t h e b o d y is beyond any doubt. It concerns the Annelids (to which belongs the common earthworm). In them segmentation is expressed internally as well as externally. See the next Figure.
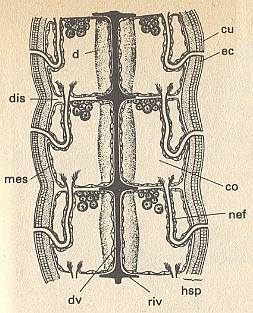
Figure 8b. Scheme of the internal structure of annelids (Annelida). Combined dorsal view and horizontal section through three segments.
co coelom, cu cuticula, d gut, dv dorsal blood vessel, dis dissepiment, ec ectoderm, hsp skin-muscle sack, mes mesoderm, nef nephridia, riv ring vessels.
( After HADORN, E., and WEHNER, R., 1977, Algemene Zoölogie )
The next Figures illustrate the organizational plan in sea-cucumbers.
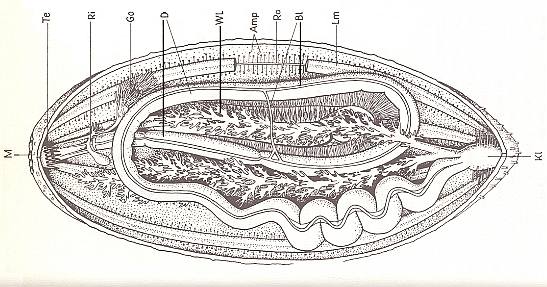
Figure 9. Organs of a sea-cucumber. Cut open at the underside.
Amp ampullae of the tube-feet, Bl blood lacunae, D gut, Go gonads, Kl cloaca, Lm longitudinal muscles, M mouth, Ra radial-, Ri ring canal of water-vascular system, Te withdrawn tentacles (i.e. the ones around the mouth, see 4 of Figure 7), Wl waterlungs.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
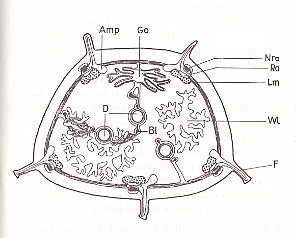
Figure 10. Organs of a sea-cucumber. Transverse section.
Amp ampullae of the tube-feet, D gut, F tube-feet, Go gonads, Lm longitudinal muscles, Nra radial nerves, Ra radial canal of water-vascular system, Wl waterlungs.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
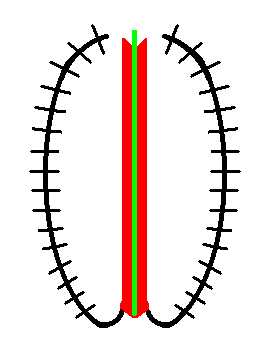
Figure 10a. Repetitional scheme in Holothurians (sea-cucumbers). The body's main axis is indicated by a green line. Axial organs are symbolized by the red elongate area (and also by the main axis itself). The (five) radial body parts (black, two of them drawn) are bent up. The short cross lines perpendicular to the bent-up parts symbolize the repetition of elements along those parts. This repetition takes place more or less parallel to the main body axis. There is, however, no segmentation of the axial organs. Oral pole of the main axis at the bottom of the image, aboral pole at the top.
The same tectological situation we find in Echinoids (sea-urchins).
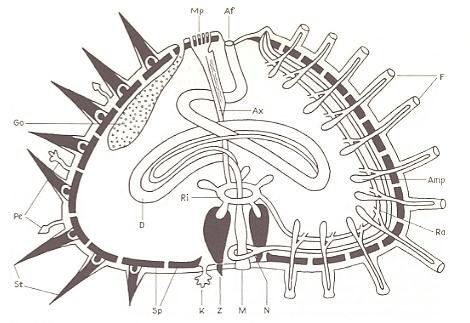
Figure 11. Schematic longitudinal section through a sea-urchin, left : interradius, right : radius.
Af anus, Amp ampulla of tube-foot, Ax axial organ, D gut, F tube-feet, Go gonad, K gill, M oral aperture, Mp madreporite, N nerv ring with radial nerve, Pe pedicellariae, Ra radial vessel of the mesocoel, Ri ring vessel of the mesocoel with collecting bladders, Sp skeletal plates, St spines, Z tooth of the masticatory apparatus.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
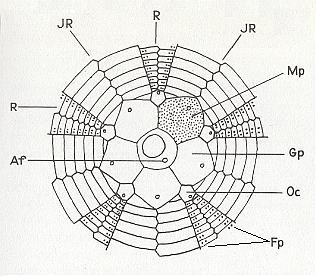
Figure 12. Scheme to indicate the skeletal elements of a regular sea-urchin.
R radial plates, IR interradial plates, Oc ocellar plate, Mp madreporite, Af anus, Fp pores for the ambulacral tube-feet.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
In Crinoids (sea-lilies) the situation with respect to e p i m e r s is more difficult to interpret. The sessile stadia (and also some adult forms) possess a stalk (stem) which is composed of a large number of sequential elements. These elements are repetitions along the body's main axis. It is, however, hard to interpret these stem segments as being metamers, because almost nothing of the body elements is repeated. The calyx (the main body part which is attached to the stem) on the other hand, also in free-living forms, seems (also) to be articulated (segmented) along the body's main axis. But even these rings (originally, the ring of infra-basal plates, ring of basal plates and ring of radial plates) are hard to interpret as being metamers. Because of an apparent lack of complete (or almost complete) repetition, we, for the time being, do not interpret them as metamers, implying that we assess a physiological individual of a free-living, as well as of a sessile crinoid, as a fourth-order form individual, i.e. as a single metamer. The next Figure illustrates the organizational plan of a crinoid.

Figure 13. Skeleton of the calyx of a primitive sea-lily (Hyocrinus bethellianus) with high calyx walls composed of : basalia- (B) and radial plates (R), and with arms (A) only to emerge from the calyx's edge. The arms with long arm segments (Br), that branch only further up (Av). Af anus, M oral aperture, Co stem, composed of discs (columnaria). Diameter of calyx 0.5 cm.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6. )
Indeed, the Echinoderms proved to be instructive for tectological considerations and problems.
After having diverted for a while into the special tectological relationships in Echinoderms, we return to our general tectological discussion, concerning metamers or fourth-order morphological individuals.
The so-called s e g m e n t a t i o n or homodynamic composition of the whole organism (of which the physiological individuality appears in the form of a person or fifth-order form individual), as we see it in vertebrate and most articulate animals, and -- but now as articulations of off-shoots -- in most higher plants, represents a significant advancement of the organization, and we can generally assess those organisms as higher and more perfect, as compared to those in which the segmentation (resulting in a sequence of metamers) is absent, and in which consequently the physiological individual (biont) itself only reaches the value of a single metamer, as in lower worms, in Molluscs, and, as we saw, in Echinoderms.
An instructive example with respect to a correct insight into the homodynamical or metameral formation could serve the gradual transitional series of unsegmented to segmented forms, like we see in the lower worms (especially Cestodes, i.e. tapeworms) : Here we can see clearly in what way the same parts (metamers), which appear in the lower forms as physiological individuals, appear in the higher forms only as homodynamic parts.
The main reason why s e g m e n t a t i o n is evaluated here as an advance, is the following :
When an organic body becomes -- in the course of evolution -- segmented, the result is a repetition of -- in principle -- the whole body. So in a way, all segments, except one, are superfluous. Because of this, and because of the relative independence of each segment, it can specialize with respect to a new function by being reorganized (without causing harm to the organism). So in virtue of the segmentation a d i v i s i o n o f l a b o r can be developed, making the whole organism much more capable of coping with environment. When evolution proceeds further this 'colony' of specialized metamers can become centralized (i.e. getting a central 'government') which effects it to become more efficient, and this means that the metamers will (in the course of evolution) be more and more i n t e g r a t e d, which in turn means that the relative independence of the metamers is partly given up again. This has happened in the Vertebrates and in the Arthropods. There, especially in their higher forms, the segmentation is hardly visible anymore.
On the basis of this, it can be imagined that there are organisms in which this integration of metamers is pushed to the very limits, which means that no trace of any segmentation is left. And such an organism would then be tectologically assessed as being just a single metamer. In the context of Tectology, as a doctrine of macroscopic organic structure, this adjustment is perfectly right, i.e. it is right from a m o r p h o l o g i c a l viewpoint (while, in a way, wrong from a genealogical viewpoint). Indeed such an integration lowers the morphological 'value' with respect to the scale of subordinated individualities. We can even imagine that the whole process will, at some time, be repeated, i.e. the metamer becomes segmented, etc.
In the next document we will study the next two orders of morphological individuality, persons and colonies.
To continue click HERE to proceed further with systematic Tectology (Persons and Colonies)
back to homepage
back to retrospect and continuation page
back to Internal Structure of 3-D Crystals
back to The Shapes of 3-D Crystals
back to The Thermodynamics of Crystals
back to Introduction to Tectology
back to Tectology, cells, organs, antimers