We now arrive, in ascending the ladder of organic morphological individuality, to that higher organic form unit, which ordinary speech as well as the general view in Zoology (but not in Botany!) usually assesses as the individuum proper. And although a closer inspection with respect to organic morphological individuality shows that also these "proper" or absulute individuals are indeed only relative, and that they cannot claim any other individuum status as can also be assigned to the metamer and to all other, earlier considered, individualities of lower order, and although these "proper" individuals appear only as subordinated constituents of a still higher unity, the colony or Cormus, in most higher plants and in many Coelenterates, the incorrect view, departing from the individuality as we see it in Man and in higher animals, that considers the fifth-order morphological undividuals as the individuals proper, has become so wide-spread, that it is the main cause of much confusion concerning organic individuality.
Definition of a Person or Prosopon and Off-shoot or Blastus.
The true p e r s o n or o f f - s h o o t consists in all cases of a multitude of subordinated first-, up to fourth-order individuals. Each single fifth-order form individual is accordingly composed of at least two metamers, at least two antimers and also of a multitude of organs and of a multitude of cells.
Any physiological individual that does not comply with this definition -- as we see in most Molluscs -- that is not composed of metamers, but being itself just one metamer, cannot be considered as being a person.
The person or the prosopon in our sense is the morphological substrate of the physiological individuality in all Vertebrates and Arthropoda, all articulated animals at all (so also the articulated worms -- Annelids and Tapeworms). As off-shoot it composes the colonies of many Coelenterates and higher plants.
The person or off-shoot is characterised by its s e g m e n t a t i o n or articulation, i.e. in virtue of the fact that a number of homodynamic parts (metamers) lie behind each other along the main axis of the body that is composed of antimers. Usually this articulation is accomplished when an initial single metamer generates a chain of terminal buds, that are distinguished by incomplete separation walls (nodes) and remain united. We thus can determine this very characteristic, typical form of the person, which appears as bion, i.e. as physiological individual, in most higher animals, and as a constiuent of a colony in many Coelenterates and plants, as a chain of metamers that lie behind each other, and have originated by terminal budding.
Now, in addition to such metamer complexes, which all correspond to aech other by their articulated chain form, there exist other metamer colonies, that are distinguished from the first by a complete absence of terminal budding, and as a result of this look so different (i.e. different from a chain of metamers) that one interprets them as true colonies, i.e. as true sixth-order individuals. To these belong all colony-like organic entities that do not consist of segmented persons, but of unsegmented metamers. This is the case in many animals, especially colonial Bryozoans and Tunicates. Like in true colonies (cormi) these pseudo-cormi are generated by l a t e r a l b u d d i n g. The buds are however no true off-shoots, like the segmented off-shoots of most plants and Coelenterates, but simple unsegmented metamers : About their morphological equivalence to the free-living "single animals" of Molluscs there can be no doubt. Like these they are composed of two antimers and of many organs and cells, without there being any series of homodynamic parts along their axis, and can therefore considered to be just single metamers, i.e. fourth-order form individuals.
Because the morphological characteristic of the person essentially is its composition of metamers, and every single person is a multitude of closely connected parts that repeat essentially the whole set of the features of the species, we cannot interpret the so-called colonies of Tunicates and Bryozoans as genuine colonies or cormi, but as single persons.
This despite the fact that the constituent metamers are not strictly s e q u e n t i a l l y o r d e r e d in those colonies. Here we have the problem of two different but in some way intermingling (or 'interacting') criteria for some organic form unit to be a metamer, or an antimer, etc. One criterium is the way that the unit is built in : repeated a r o u n d an axis, or a l o n g it. The other criterium is the position it has within (i.e. on the scale of) a relative scale order : cells aggregate to become organs, organs aggregate to become antimers, antimers aggregate to become metamers, metamers aggregate to become persons, and persons, finally, aggregate to become genuine colonies (cormi).
Tectology of Bryozoans
Let us give a description and show some features (in the ensuing Figures) that illustrate the tectological assessment of a "colony" of Bryozoans as a p e r s o n (and not as a genuine colony, i.e. not as a m o r p h o l o g i c a l sixth-order individual). All Bryozoa or Ectoprocta are colonial (with one exception : Monobryozoon ambulans, Grzimeks Tierleben Band III), most are marine, and in the majority of cases the units or zooids of the colony secrete tubes or boxes of lime partially encasing the soft parts. Such colonies often appear as encrustations on the surface of seaweeds or other objects. Each zooid is basically cylindrical, has a ring of tentacles and at first sight seems to resemble a small cnidarian polyp, i.e. a polyp belonging to the Coelenterata (corals, medusae and the like). But the zooids are coelomate, having a freely suspended gut with both mouth and anus, and are unquestionably of a higher grade of organization. A short description of some features of the genus Bowerbankia, Order Ctenostomata, Bryozoa, will exemplify and illustrate the tectological status of a bryozoan colony (i.e. pseudo-colony).
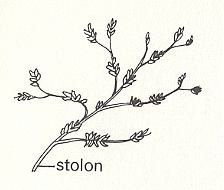
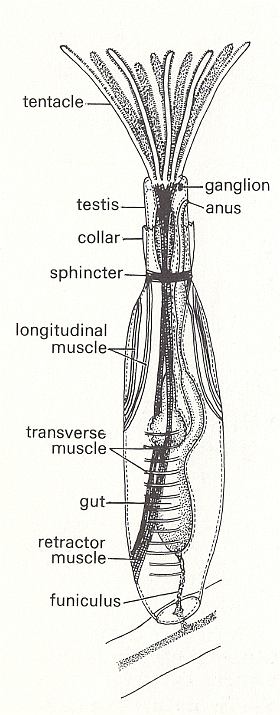
Bowerbankia (See Figures) has a creeping cylindrical s t o l o n from which numerous bottle-shaped zooids arise in clusters. Each living zooid is housed in a skeleton (membranous in this case) called the c y s t i d. The whole colony may be called a z o a r i u m. When feeding the zooid extends its l o p h o p h o r e, a ring of ten ciliated tentacles, into the surrounding water. These tentacles converge on a central mouth from which the gut descends into the body of the zooid. This gut is U-shaped, hanging down in the coelom, and has an oesophagus, stomach and intestine which opens by an anus close to the mouth but outside the ring of tentacles. The tentacles attach to the body by an eversible tentacle sheath. In Bowerbankia and its relatives the sheath is protected by a collar, but this is not represented in other Bryozoa. Between mouth and anus is a major ganglion from which nerves run to all parts of the body. From the base of the stomach a thread, the f u n i c u l u s, connects to a main f u n i c u l a r t u b e running along the stolon and connecting all the zooids. The tentacles can be either extruded for feeding or retracted entirely within the body for protection. When the zooid feeds, the tentacles are extended in an erect funnel by hydrostatic pressure of the coelom. There are no separate excretory, circulatory or respiratory organs. The colony grows from an initial zooid, the a n c e s t r u l a, by growth of the stolon (itself a series of modified zooids) and by asexual budding. New colonies, however, are produced sexually [CLARKSON, E., 1980, Invertebrate Palaeontology and Evolution].
Figure 1. Bowerbankia, recent member of the order Ctenostoma. Appearance of colony.
( After CLARKSON, E., 1980, Invertebrate Palaeontology and Evolution )
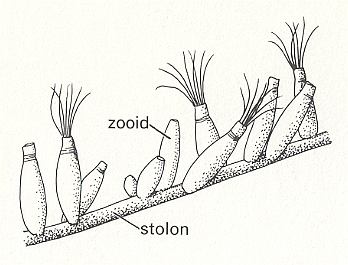
Figure 2. Bowerbankia, recent member of the order Ctenostoma. Section of zoarium with extended and retracted zooids.
( After CLARKSON, E., 1980, Invertebrate Palaeontology and Evolution )
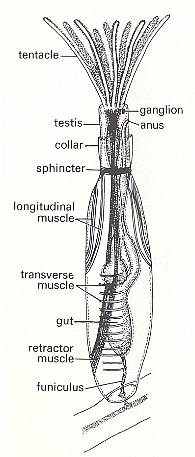
Figure 3. Bowerbankia, recent member of the order Ctenostoma. Zooid with tentacles extended.
For a higher quality picture click HERE
( After CLARKSON, E., 1980, Invertebrate Palaeontology and Evolution )
From these Figures (and the description) we can clearly deduce that a single bryozoan zooid is composed of (a complete set of) organs (themselves composed of cells), that together form (at least) two antimers. The zooid shows NO sign of segmentation (along its main axis), i.e. it has no metamers. So it itself must be a single metamer. And the colony, or parts of it, should be interpreted as a person.
We see, by the way, that we had to appeal to some p h y s i o l o g i c a l features, which in itself is not entirely appropriate within a strictly morphological consideration. This shows that FORM and FUNCTION cannot be wholly separated in any biological account.
The fact that in Bryozoans we don't have to do with true off-shoots is demonstrated by other Bryozoans, for instance Membranipora : The fronds of flat seaweeds such as Laminaria are often covered with the l a c y calcareous skeletons of this Bryozoan, each colony being composed of hundreds of individuals (CLARKSON, op. cit.).
Further, strong flat pieces up to 18 cm height, are formed by the wide-spread Bryozoan Flustra foliacea (See Figures 4 and 4a), the colonies of which, that are attached by means of a disk to stones, often form meadows on the sea floor at 5 up to 100 meters depth. The cystids ( = the part of the zooid that moreover consists of a skeletal box in which the zooid can retract), and thus the "individuals", are ordered two-dimensionally like adjacent cells in a lattice (KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6, p. 479). No "off-shoots" are apparent in such "colonies".
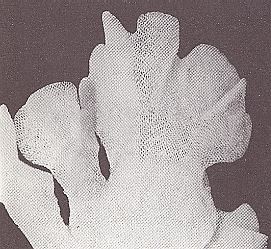
Figure 4. Part of a Flustra foliacea colony from the North Sea at Helgoland.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6 )
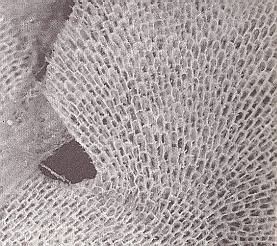
Figure 4a. Blow-up of the Bryozoan of Figure 4. The cellular array of cystids is shown.
( After KILIAS, R., 1971, in Neue grosse Tier-enzyklopädie, Band 6 )
In virtue of the considerations just given, we cannot interpret each single "off-shoot" of a Bowerbankia colony as a person, and thus the whole colony is not a sixth-order form individual, but a person, despite the fact that the repetition of its metamers is not sequential (in the single zooids there no repetition at all, except with respect to its having two body halves which are undoubtedly antimers).
So in Bryozoans we have to do with a special type of persons, in which the metamers are not sequentially, i.e. one-dimensionally, ordered. Their repetition takes place in two dimensions (a bush-like 'colony', as we see in Bowerbankia, can be seen as a plicated two-dimensional surface giving place to the individual zooids : These latter are, as far as I know, never placed directly on top of each other, which would hamper feeding).
We can call the usual persons with sequential metamers a x i a l p e r s o n s (Personae axiales), while all other persons can be called p l a n a r p e r s o n s (Personae planatae).
Haeckel, Generelle Morphologie, p. 325, distinguished these two types of persons by the names "chain persons" (Prosopa catenata) and "bush persons" (Prosopa fruticosa), but the last name does not cover all non-sequential persons, for example not the ones we see in the above mentioned Flustra and the like. The 2-dimensional aspect really is the main characteristic of those persons and distinguishes them from the chain persons. To emphasize this distintion we will call the latter axial persons.
Some Bryozoans have colonies that also should be tectologically determined as persons (planar persons) because of the structure of the constituent zooids, but in addition to this possess an overall 'colonial' structure that distinguishes them even more from what we normally consider to be colonies. These are the Fenestellids, which are palaeozoic Bryozoans of the Order Cryptostomata (Ordovicium -- Permian). Fenestellid 'colonies' are an example of a highly integrated colony, much more than just an aggregate of individuals. The colony operates as a whole, and this reminds us of the persons of higher animals, like Vertebrates and Arthropods. We will give some images of these fenestellids, followed by a description of their assumed way of feeding.
Zooecia are the skeletons ('appartments') of the individual zooids. Zoarium means the whole living 'colony'.
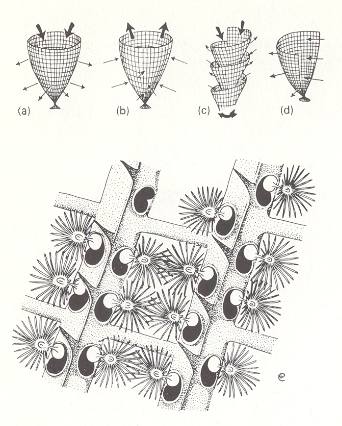
Figure 4b. Morphology and direction of currents inferred in fenestellids (Bryozoa).
(a) Carboniferous fenestellid with inward-facing zooecia.
(b) Silurian fenestellid with outward-facing zooecia.
(c) Archimedes (Carboniferous -- Permian), a spiral colony.
(d) Fan-shaped fenestellid, using unidirectional current.
(e) Fenestella (Carboniferous -- Permian) : part of zoarium showing zooids reconstructed as extending into fenestrules, i.e. as extending such that their tentacle fans occupy the rectangular openings of the skeleton. (x 8 approx.)
( After CLARKSON, E., 1980, Invertebrate Palaeontology and Evolution )
The arrangement of the zooecia and the form of the zoarium are clearly important in understanding how the colony operated as a whole. It has been postulated that the 'operational subunits' of the fenestellid colony are the fenestrules (i.e. the rectangular window-like openings of the skeleton). In the main branches, the lophophores (i.e. the tentacle bearing structures) of equally spaced zooids on opposite sides of a median ridge are considered to have extended into the fenestrule forming a filtering net, the cross-bars giving support and dividing up the fenestrules. A combined feeding current set up by the zooids would draw water in through the fenestrules, enabling them to strain off all the food material with their lophophores, and would thus give high filtering effciency.
Let us describe a second case of "colonies" that are not genuine sixth-order form individuals but (planar) persons : Social Tunicates.
Tectology of Tunicates
All Tunicates or Urochordata are marine animals, some of them free-living (swimming), others in their mature state sedentary Chordates. The fact that they are Chordates indicates that they are related to Vertebrates because in most of them the tail of the larva contains the c h o r d a : A characteristic feature of the building plan of the chordate animals (Chordata : Tunicata (Urochordata), Cephalochordata (Acrania) and Vertebrata (Craniota)), is, among others, the c h o r d a d o r s a l i s as a permanent or only embryonal or larval central structure of the axial skeleton.
The ectodermal ephithelium of the Tunicates secretes a cuticular m a n t l e containing cellulose and covering the whole body. The anterior part of the gut is transformed into a g i l l b a s k e t which is strongly developed. It serves for respiration and feeding. The gill openings (cracks) end up in a peribranchial chamber which is dorsally widened forming the c l o a c a, in which also the intestine and the outlets of the genital organs end up. The Tunicates are sack- or barrel-shaped. Only in one group, Appendicularia, the tail of the larva is retained.
The Tunicates divide into three groups,
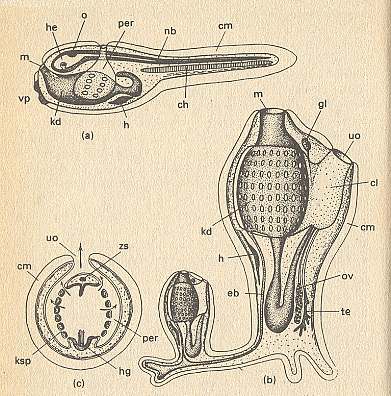
Figure 5. Schemes of the organization of Ascidia (Tunicata).
(a) Larva of ascidium.
(b) An ascidium which has generated a daughter ascidium at a basal sucker (stolon) by means of budding.
(c) Transverse section through an ascidium at the level of the outlet.
ch chorda, cl cloaca, cm cuticular mantle, eb entodermal tube going out from the gill basket, penetrating the stolon, and furnishing the entoderm of the bud, gl ganglion coming from the larval brain, h heart, he brain, at its underside a static sense organ, hg hypobranchial groove, kd gill basket, ksp gill openings, m mouth, nb neural tube, o eye, ov ovary, per peribranchial chamber, te testis, uo outlet, vp papillae for attachment, zs nerve string that remains as left-over of the neural tube.
( After HADORN, E., and WEHNER, R., 1977, Algemene Zoölogie )
Among the A s c i d i a we have the star-ascidium Botryllus schlosseri. Its length is 2--2.5 mm. This is a colonial Ascidium. The animals are embedded in a jelly matrix and approximately radially positioned around a common cloaca. See next Figures.

Figure 6. Photograph of the star-ascidium, Botryllus schlosseri. It is a colonial Ascidium. Each colony consists of a few radially positioned individuals around a common cloaca.
( After KRAUS, O., in Grzimeks Tierleben, Band 3 )
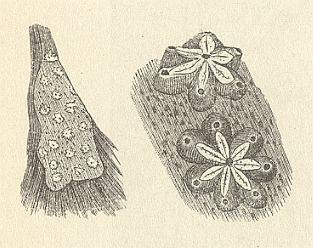
In the above photograph of Figure 6 we see many small colonies of Botryllus schlosseri. The number of participating individuals varies as can be seen. Some colonies consist of six, others of seven, others of eight, etc. The radiate ordering is not a very regular one.
The free-swimming S a l p i d s (Thaliacea) are animals of the open sea, living at the surface levels, and often appearing in very large quantities. Their body is barrel-shaped, in- and outlet are positioned at the body ends, and thus opposite to each other. A number of them displays a very complex alternation of generations. Temporarily or permanently colonial.
The salpids proper (Salpida) show an interesting colonial behavior. The single living salpid originates from a fertilized egg and should accordingly be called an o o z o i d.
One can denote the oozoids as "nurses". In them no genital glands are present. Reproduction in them is asexual and takes place at a special organ, the s t o l o p r o l i f e r. It is placed ventrally behind the body center of the salpid, namely behind the end of the hypobranchial groove. Here many buds arise, that finally develop into sexual animals (gonozoids). These sexual individuals are ordered in two symmetric rows. Often they remain, also after unleashed from the stolon, united by means of attaching papillae. In this way the well-known salpid chains originate, for instance in Thalia democratica. They can reach lengths of over 25 meter. See next Figures.
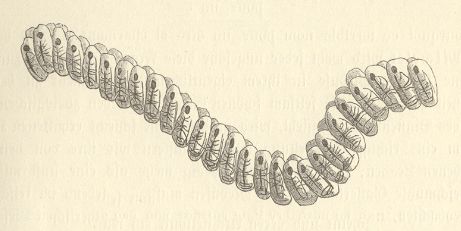
Figure 9. Thalia democratica.
End part of a young salpid chain (three individuals shown). Each individual contains an ovary and testis.
( After KRAUS, O., in Grzimeks Tierleben, Band 3 )
As has been said, the salpid chain, as we see it in Thalia democratica, is a double row of individuals. The rows are symmetrically positioned with respect to each other. A pair of gonozoids is repeated along an axis (the axis of the colony). Each pair consists of two individuals each having the value of a metamer or fourth-order form individual. So in this case we have to do with a p l a n a r p e r s o n, (Persona planata), because the constituent metamers are repeated over a s u r f a c e (If the salpid chain would consist of just a single row of individuals, then it would represent an axial person (Persona axialis)). In the case of salpid chains the individuals are more or less loosely connected, in sharp contrast with the metamers that compose a Vertebrate or Arthropod body. But the chain phenomenon is constant and so is intrinsic (i.e. not accidental), so it is part of the building principle of the species, i.e. of its tectological character.
The Bryozoans and Tunicates have shown us a different type of person : Their "colonies" are not genuine colonies, i.e. not true sixth-order form individuals, like we see in most higher plants. They are persons, but not the usual persons, axial persons, like we see in Vertebrates and Arthropods, but planar persons. In them the constituent metamers are ordered and distributed over a more or less plicated surface, rather than being ordered along a line (axis).
When determining whether some organic entity is a person or off-shoot, i.e. a m o r p h o l o g i c a l fifth-order individual, we shouldn't allow ourselves to be influenced too much by the p h y s i o l o g i c a l status of such an organic entity, because the consideration is supposed to be a purely m o r p h o l o g i c a l one. This is especially relevant with respect to p e r s o n s, i.e. free living fifth-order form individuals, and less so with respect to off-shoots, which are, it is true, also fifth-order form individuals, but are not free-living organic entities. Tectology is about general s t r u c t u r a l features and principles that are believed to be common to all organisms, features that concern their spatial built-up. The general built-up of organismic entities seems to be the result of only a few possibilities available to organismic matter. Each organismic type inhabits certain specific types of environment, and these latter demand special physiological functions to be performed in order that the organism can maintain itself in such an environment. This leads to a special molecular machinery like the production of appropriate enzymes, it also leads to some macroscopic morphological features that ensure the organism's adaptation to its environment, which could consist of the need for the ability to move around in a special way. When the latter is, in certain cases, in some way not necessary or even detrimental, a sedentary life would be the better option. All of this strongly determines the organism's f o r m or s h a p e. This form or shape is thus dependent on the physiology and ecology of the given organism, i.e. it is subordinated to physiological demands. That's why we often see within one and the same animal group different morphologies. This means that the individualities, as are until now established in our investigation, are not a b s o l u t e entities, living as it were their own life as some sort of independent units or organic 'atoms'. Indeed in inorganic individuals, like in single crystals, we see that the constituent atoms are stable units which can combine in a multitude of ways to constitute all kinds of minerals.
The person or prosopon always reminds us to our own state of individuality, which confers on it an aspect of independency and self-containedness, less clear in many animals, and absent in composite plants (where the off-shoots are not even physically free).
It could turn out that in assessing the order of morphological individuality of one or another organic entity, we need the p e r s o n as one of two points of reference, the other being the cell. Indeed, the cell really seems to be an atomic entity, a building unit, albeit that it can appear in a multitude of possible cell forms. We then have two points of reference, and thus we have a s c a l e on which we can 'measure' the order of individuality of any organic entity which is not a person nor a cell. While the morphological definition of a cell (as first-order form individual) will fairly be possible along morphological lines, and so providing the first point of reference of our tectological scale, the definition of a person, which should provide the second reference point of that scale, could demand some physiological criteria, i.e. criteria lying outside the domain of morphology. This is not totally objectionable because, as we remarked earlier, FORM and FUNCTION are interdependent in the organic world.
The highest degree of morphological completion with respect to the composition out of different individualities is found in those organisms, in which a multitude of fifth-order individuals goes together to make up a still higher form unit, the Colony or Cormus. This is the sixth and last level of individuality that is accomplished by organismic entities in their progress towards their structural development.
Definition of a Colony or Cormus.
By a C o l o n y or C o r m u s we mean exclusively that organic form unit, which consists of a multitude of off-shoots or fifth-order form individuals. These off-shoots normally originate laterally from a previous off-shoot, resulting in a bush-like appearance of the colony.
The next Figures diagrammatically show the phenomenon of lateral budding of off-shoots (blasti), resulting in a cormus.

Figure 10. Diagram of lateral budding of off-shoots, resulting in a bush-like appearance of the colony.
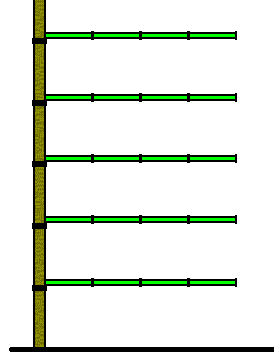
Figure 11. Diagram of lateral budding of off-shoots, resulting in a regular appearance of the colony. The vertical stem represents part of the main shoot. This main shoot consists of sequential metamers. The off-shoots, budding laterally from the basal part of each stem metamer, themselves also consist of sequential metamers.
In virtue of their property, as they have it in cormi, to be subordinated constituents of a cormus, we indicate such persons with the name off-shoots or blasti. We accordingly exclude from the morphological concept of C o r m u s all those colony-like organic entities, that in Botanics as well as in Zoology are, it is true, very often indicated as colonies (cormi), but which are not genuine colonies. Such pseudo-cormi we have met in the previous section on Persons. They were the "colonies" occurring in Tunicates and Bryozoans, which only rank as persons. Such pseudo-cormi are also those of many lower plants and protists, in which the components of the colony-like entities are not fifth-order, but first-order form individuals, i.e. cells, for instance the colonies of Diatomeans and algae. All these p s e u d o - c o r m i are more or less identical to the true colonies only by the fact that both are aggregations of (often more or less loosely connected) individuals of a subordinated order. They differ from them by the fact that those constituent individuals are never fifth-order individuals. So it is just the composition out of subordinated individualities, often enhanced by a superficial similarity, that has led to the confusion of pseudo- and genuine cormi. Especially the mode of fission, namely lateral budding, that is a common feature in both of them, seems to be the cause of the mentioned confusion. Another reason is the lack of interest to set up a g e n e r a l morphology, in order to trace all-encompassing laws that dictate the construction of organic forms. In many pseudo-cormi of Diatomeans, Flagellates, algae, etc. it is single cells, in the previously discussed Bryozoans and Tunicates it is metamers, that produce branched entities similar to those effected by colony-forming persons, in virtue of continued lateral budding. It is, however, of the utmost importance to General Morphology to acknowledge the essential difference between the true colonies, i.e. sixth-order individuals, and those false pseudo-cormi : fifth-order individuals (persons), or second-order individuals (organs). The expression c o m m u n i t y can generally be applied to all these similar colony-like aggregations and means nothing else than the unity of a multitude of individuals of lower order resulting in a morphological unit of higher order. The true colony or cormus on the other hand, is a special sort of such colonies, namely only that highest and most complete sort, which is constituted of fifth-order individuals.
Because the cormus is the highest and final order of all six orders of individuality, it never can appear as integrated constituent of a yet higher order, as is the case with all five subordinated individualities, and by reason of that the morphological and physiological individuality in a certain sense always coincide in it. In virtue of the fact that the morphological character of the person or of the off-shoot is, as we saw above, a definite one, so in the same way the morphological character of a cormus, which always is a multitude of off-shoots, must be definite too. Every cormus accordingly does not only consist of a multitude of off-shoots, but also of course of a multitude of metamers, antimers, organs and cells, because indeed every single off-shoot itself already consists of these four subordinated individualities.
The true colonies or cormi reach their highest development and widest spread in the p l a n t k i n g d o m, where most higher (phanerogamous) plants and higher cryptogamous plants are developed into sessile colonies. Only the "simple", i.e. not branched plants remain at the level of a person (axial person).
In P r o t i s t s genuine colonies seem to be rare. Perhaps certain sponges enjoy such a status. Most so-called colonies of Protists, for instance those of Diatomeans and Flagellates, are false cormi, that can only claim a morphological second order ranking (organs).
Much less wide-spread and developed than in plants, genuine colony-formation is in the a n i m a l k i n g d o m, where it is almost confined to certain Coelenterates (polyps, etc.). In V e r t e b r a t a and A r t i c u l a t a it is wholly absent. What were, in the lower forms of the latter, namely lower annelids and the social tapeworms, indicated as colonies, are in fact just axial persons (like in Arthropods and Vertebrates), and what were called "individuals proper", equivalent to persons (proglottids or segments), are metamers. What were called colonies in Tunicates and Bryozoans, are, as we have seen, pseudo-cormi, they are planar persons. The so-called solitary Tunicates, for example Doliolum, the solitary generation of Salpa, are bionts which morphologically rank as metamers. Genuine cormi we find in Botryllus (a Tunicate) : There the colonies are composed of "systems" (which we already encountered earlier, as the 'stars' of star-ascidia). Each system ('star') is a fifth-order morphological individual, a person, which is composed of 5--10, but often more than 100, metamers (Haeckel, 1866), which in this case are not longitudinally, but radially (in the way antimers normally do) positioned around a common egestion aperture (cloaca). It could seem reasonable to consider those metamers not as metamers but as antimers, because of their radial positioning (That they are not persons was already argued above). It seems, however, that in the present case the positioning is not decisive, because also in Salpa pinnata and the like, the metamers are positioned in a circle around an (imaginary) axis, while they are, sometimes obliquely, sometimes transversally, ordered into regular chains in other salpids (Haeckel, 1866). Many Coelenterate colonies are probably genuine cormi, where the segmentation of the persons, i.e. their composition out of metamers, is often clearly expressed (in Anthozoans -- corals and the like -- as well as in hydroid polypes). But in addition to them also pseudo-cormi occur, without segmentation, composed of fourth-order form individuals, which are united into planar persons.
Most cormi originate by lateral budding of off-shoots that remain united with their progenitor. To these belong most cormi of the higher plants and hydroid polyps. Also many Anthozoan colonies originate by this mode of fission.
The different forms of colonies are very diverse and offer many and often conspicuous analogies between the higher plants and Coelenterates.
Important in this respect is the character of the prime person (main shoot, Blastus primarius). The stronger the main shoot is developed relative to the side shoots, the more it is predominant over the latter, the more definite is the individual character of the cormus. The lesser this is the case the more the whole cormus appears as just an aggregate of coordinated off-shoots.
Depending on the subterranian or supraterranian develpoment of the main shoot, and on the mutual relationship between the main shoot and the side shoots (Blasti secundarii), as well as on the differentiation of the latter into sexual and asexual off-shoots, we can, in plant cormi, distinguish many cormus forms indicated by many different names.
At first two main groups can be distinguised :
As s i m p l e c o l o n i e s (Cormi simplices) we denote those in which either all off-shoots are sexually, or only the main shoot is asexually, while all side shoots are sexually developed, for example every annual branched plant in which all branches carry terminal flowers, further, simple plants with only a single umbel, simple unbranched annual Composita (Plants with composite flowers) with only a singe flower head (composed of many florets). With respect to the latter two we must not be confused by the term "simple plants" : They are colonies nevertheless, because an umbel as well as a flower haed of the Compositae, consists of many florets, each of them should be considered as an off-shoot.
As c o m p o s e d c o l o n i e s (Cormi compositi) on the other hand, we denote those in which not only the main shoot, but also a part of the side shoots, are asexually, while the rest of the side shoots are sexually developed, as is the case in most higher plants.
Among these we can further distinguish annual composed colonies or stems (Caules) and supra annual composed colonies or trunks (trunci). Further, those plants that have subterranean trunks and supraterranean stems, are called shrubs (suffrutices). Those plants that have trunks that are branched already directly from below, without predominance of the main trunk, are called bushes (frutices). Finally, plants that have trunks of which the lower branches soon perish, and the higher branches form a crown, are called trees (arbores).
Very similar distinctions with respect to colony development as introduced in higher plant morphology, can also be observed in Coelenterates which form genuine cormi, namely in anthozoans and hydroid polyps. In the meantime we should, in Coelenterates, also consider several other types of colony formation, especially by longitudinal fission, that in plants either do not occur at all, or only in lower plants.
Further one should especially discriminate between colonies that, as in most higher plants, consist of axial persons (off-shoots), a x i a l c o l o n i e s (Cormi axiales), and those that consist just of planar persons (off-shoots), p l a n a r c o l o n i e s (Cormi planati).
For an understanding of the (characteristics of the) true cormus, and its relationship with the subordinated form individuals of which it is constituted, as well as for a correct tectological assessment of the relationship between the six orders of morphological individuality, no animal group is so significant as the C o e l e n t e r a t e s, especially the Siphonophores and the colonial corals (Anthozoa). The morphological parallel of these with the h i g h e r p l a n t s can be carried through through all six orders of morphological individuality, and even through their subcategories.
As composed c o l o n y the blooming tree corresponds entirely to the mature coral colony. Both are constituted by a multitude of sexual and asexual p e r s o n s (off-shoots). Each person, each flower off-shoot of the tree, each polyp of the coral colony, in turn consists of a multitude of m e t a m e r s, or stem segments (leaf rings of the flower, tabulae of the polyp), and of a multitude of a n t i m e r s (radial parts). Each metamer and each antimer then is, first of all, a multitude of o r g a n s of different order, and at the same time a multitude of several epimers (segments of each leaf [in feathered leaves], segments of each tentacle), and of paramers (halves of the leaves, halves of the tentacles). Finally each of these morphological units is a multitude of connected first-order form individuals or c e l l s.
This interesting comparison is done by Haeckel (1866). It should however be noted that the spaces enclosed by tabulae, present in individual corals, cannot be interpreted as metamers, because each tabula just marks off the section of the skeleton not inhabited anymore by the polyp. The latter is shifted upwards during growth. Moreover, as far as I can see, not any Coelenterate shows a repetition of parts (representing the whole body) along its main axis (Only Scyphopolys do so -- namely during strobilation -- but this is only temporary : The parts that originate by strobilation are released one after the other in the form of larvae). Consequently every polyp -- i.e. every polyp that is normally considered as a single polyp, i.e. without (considering buds) -- and also every medusa, is just a single metamer, like we see in Molluscs, Tunicates, Echinoderms and Bryozoans. A "colony" of these metamers is consequently a fifth-order individual.
By the way, the fact that Coelenterates form colonies just consisting of metamers is in line with their being very low animals, i.e. animals with a very primitive organization.
With the consideration of true c o l o n i e s or c o r m i, our theoretical investigation, concerning the organic m o r p h o l o g i c a l individualities, has come to an end. Well, at least the basics have been laid down. The theory as it stands is, however, in need of much more c o n f i r m a t i o n. This further confirmation cannot be done on the basis of formal theory, but must result from an extensive empirical survey of the whole organic world. As a first stage of such a survey we will, in the ensuing documents, consider the p h y s i o l o g i c a l i n d i v i d u a l i t i e s. This means that will consider the occurrence of the six individualities, as established above, a s p h y s i o l o g i c a l i n d i v i d u a l s or bionts. We could for example ask ourselves : "Can single antimers also somtimes occur as physiological individuals, i.e. as biologically independent entities, and if so, where in the organismic world do we see them so occurring?". The same question can be asked for all the other form units (Cells, Organs, Metamers, Persons, Colonies).
To continue Tectology click HERE to begin with the study of the PHYSIOLOGICAL INDIVIDUALITY in organisms.
back to homepage
back to retrospect and continuation page
back to Internal Structure of 3-D Crystals
back to The Shapes of 3-D Crystals
back to The Thermodynamics of Crystals
back to Introduction to Tectology
back to Tectology, cells, organs, antimers
{kind=link}