Orientation
Each of the six essentially different form units, which we, in the foregoing, have distinguished as six different orders of morphological individuality, appears in certain organic (biological) species as physiological individual or biont. In a section of our introduction to Tectology we had indicated with this expression that uniform spatial magnitude, which, as living organism, as more or less centralized life unit, is able to independently exist during a longer or shorter period of time. An existence, expressing itself in all cases by the performance of the most general organic function, namely of self-maintenance by means of metabolism. Also other life functions can be performed by the physiological individual : Reproduction or maintenance of the species, as well as to maintain its relations with the environment, for example by locomotion. But the actual performance of these functions needs not necessarily enter the concept (definition) of biont (i.e. in order to be a biont or physiological individual it does not need to constanly execute those functions. Only the function of self-maintenance must constantly be executed). The biont or functional individual is accordingly not (in contradistinction to the morphological individual) an indivisible spatial magnitude, which we, at the moment of consideration (assessment), should see as unchangeable (indivisible in the sense that we cannot take away a part of it without destroying its character as form individual). Rather the physiological individual is a uniform coherent spatial magnitude, that we see living during a longer or shorter period, i.e. that we see maintaining itself by metabolism, and which we accordingly view at the moment of consideration as changeable. Also parts of the functional individual can separate themselves from it, without its individuality, i.e. its continued existence as independent life unit, becoming to be at risk, and when the biont reproduces, this separation of parts indeed takes place regularly, parts that can develop into new bionts. Accordingly we can summarize the important difference between morphological and physiological individuality as follows :
The p h y s i o l o g i c a l i n d i v i d u u m (biont) is a single organic spatial magnitude, which, as centralized life unit is capable of self-maintenance and at the same time is divisible, and which, in virtue of the actions that are associated with these functions, can only be considered (assessed) as changeable at different points in time.
The m o r p h o l o g i c a l i n d i v i d u u m (first- to sixth-order) on the other hand, is a single organic spatial magnitude, which as a completely delimited form unit is indivisible, and which as such can only be seen, at a particular moment in time, as something unchangeable.
Each of the six morphological individualities of different order, which we have characterised in the previous documents, is able to represent the physiological individuality, and each biont, which as a mature representative of the species, possesses a higher degree of morphological individuality, must have passed, if it develops from a fertilized egg or from a non-fertilized cell (spore), through all the previous lower degrees of morphological individuality during its developmental cycle.
Our task here is to show that each of the six degrees of morphological individuality can function as biont. But before we set out on that task and scrutinize all six orders of individuality, it is necessary to distinguish between three essentially different manifestations or sorts of physiological individuality, which generally can be signified as the actual biont (or biont in the narrow sense), the virtual or potential biont, and the partial or apparent biont.
Actual Biont, Virtual Biont, Partial Biont
An a c t u a l b i o n t or physiological individual in the narrow sense can be called every completely developed organic individual, which has reached the highest degree of morphological individuality, that is entitled to it as mature completely developed representative of the species.
This degree is a definite one for every organic species. For example the actual biont in higher plants is a six-order morphological individual, in Vertebrates it is a fifth-order, in most Molluscs it is a fourth-order, in Volvox, and in many Foraminifers it is a second-order, in unicellular organisms, finally, it is a first-order morphological individual. Only with respect to the third-order form individual (antimers) it is not certain whether it can, and if so, whether it indeed does, occur as physiological individual.
There are of course some organismic species that experience in their final developmental stage (the stage in which they can reproduce) a partial reversal to a lower degree of morphological individuality. But these are exceptions caused by nature's exploration of the space of possibilities of development. Almost every such possibility is tried out by her in order to be able to occupy and utilize every niche present on this remarkable planet. Nature can and will deviate from "types" when necessary and experiments into all directions.
A v i r t u a l b i o n t or potential physiological individual, can be called every undeveloped organic individual, as long as it has not yet reached the highest morphological degree of individuality, that the mature individual is entitled to have and to which it can develop.
This degree is different at different points in time and in different stadia or periods of the individual development.
In many organisms every single part of the body can, under certain conditions, behave as, and thus be, a virtual biont, and thus develop into an actual bion, as is the case in Hydra (a fresh water polyp) and in many plants, where many cell groups of the body have such an extraordinary ability to reproduce, that they themselves can, when separated from the parent organism, i.e. from the actual biont, again develop into such a whole.
A p a r t i a l b i o n t or apparent physiological individual can be called any part of an organic individual, that possesses, after its separation from the potential or actual biont, the ability to maintain itself for a longer or shorter period, and to continue its existence as apparent independent biont, without, however, to be able to develop into the actual biont.
The apparent or partial biont can never (as virtual bionts indeed can) develop into the whole organism, i.e. gradually develop into the actual biont by independent growth. Rather it will perish after it had maintained itself for some time, and sometimes had performed a certain function (for instance reproduction) in this period of time. Such is the case for example with the hectocotylus (an organ) of the Cephalopods, the proglottis (a metamer) of Cestods (tapeworms), which have separated themselves from an actual bion of higher order (Haeckel, 1866). As one sees, this concept of partial or apparent biont is a very broad and undetermined one, and it is by far not as significant as are the essential different virtual and actual bionts. However in earlier (i.e. before 1866) attempts to determine the organic individuality, the partial biont was considered to be very important, so it is perhaps not superfluous to introduce the partial biont as a third manifestation of physiological individuality alongside the virtual and actual biont.
Above we articulated the important statement that each of the six morphological individualities can appear as biont or physiological individual. We must now add that this statement is valid for all three manifestations of the biont. The actual, the virtual, as well as the partial can be represented by each of the six morphological individualities. In this way we get eighteen possible organic life units, with which we will now sequentially deal, starting with the lowest and finishing with the highest order. In each order of morphological individuality, we will first deal with the actual biont, which is the most important, next the virtual biont, and finally the partial biont. We start with the first-order morphological individuum, the c e l l.
All single eggs, all single spores, and all non-colonial unicellular organisms, are single independent cells. But only the non-colonial fully developed unicellular organisms are cells appearing as actual bionts.
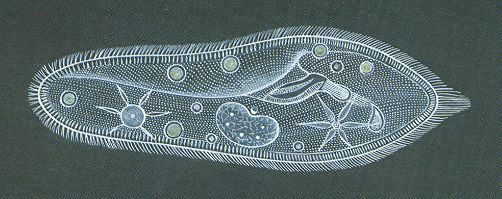
The next two Figures depict two species of the unicellular Class Flagellata.


The case, in which the physiological individuality is represented by cells or first-order form individuals, is the most abundant, insofar as not only all organisms summarized above, that, as actual bionts, are first-order individuals, and enjoy this status during the whole period of their existence, but also all higher organisms (second to sixth morphological order), which develop from a single egg or spore, appear in their initial phase as single cells that develop into a multicellular body, and these intial cells are first-order v i r t u a l b i o n t s.
Many cells, which do not have the above mentioned ability to -- when separated from the parent organism -- develop themselves further, resulting in an actual biont, nevertheless are able to maintain themselves for a longer or shorter period. An example are the sperm cells of animals. They can sustain themselves for some period but can only develop into an actual biont when they have been fused with an appropriate egg cell. The same applies for single-celled pollen grains and most unfertilized single-celled eggs.
While the cells, as first-order form individuals very often represent at the same time the physiological individuality, this is much less often the case with respect to organs or second-order form individuals. Nevertheless it is more common than would seem to be possible at first sight. It is in this respect important to recall the m o r p h o l o g i c a l meaning as was defined earlier. As an organ in the morphological sense we indicated every constant uniform spatial magnitude possessing a definite form, and which is composed in a constant way of a more or less determined number of cells, and which does not show the positive form features of the third- to sixth-order form individuals.
|
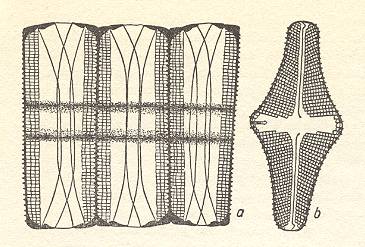
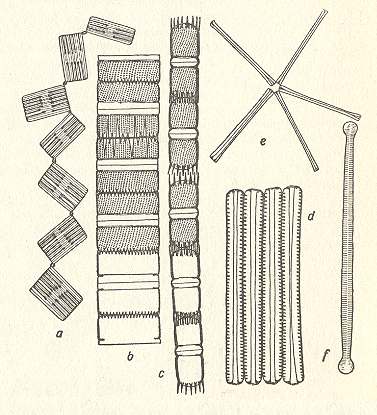
Figure 6. Colony formations. (a) : Tabellaria with jelly intercalaries. (b), (c) : Melosira chains. (d) : Fragilaria tape. (e), (f) : Asterionella star [(f) shell view] ( After HUSTEDT, F., 1956, Kieselalgen (Diatomeen). ) |
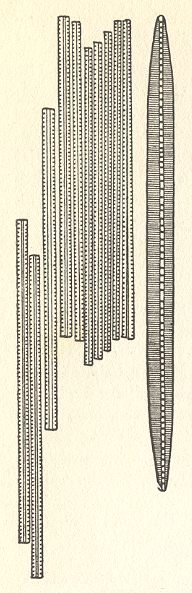
The Diatomean cell is in fact some sort of pill box composed of two main parts, the bottom part and the lid, which encloses the former. The content the box consists of the organic material of the cell.
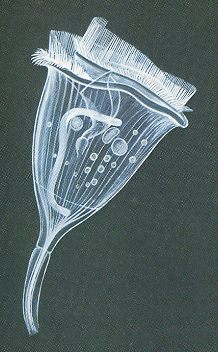
The next Figure depicts yet another second-order physiological individual, a colonial Flagellate.
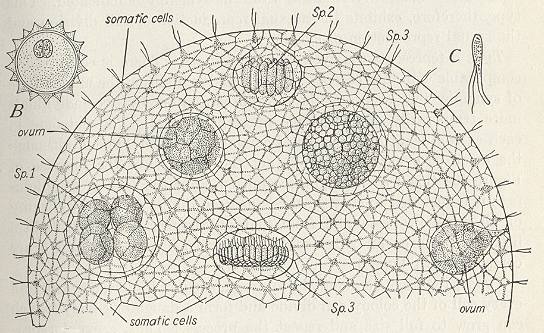
Figure 8. Volvox globator, a colonial flagellate. Main image : one-half of the spherical colony. B : zygote, with male and female nuclei uniting, and protective shell secreted after fertilization. C : microgamete or spermatozoön. Sp. 1, 2, and 3 : developing sperm-spheres.
( After CURTIS, W. & GUTHRIE, M., 1947, General Zoölogy )
As with the Diatomean colonies, the colony of Volvox consists of a multitude of cells, while there is no sign of a regular repetition of parts that could each for themselves represent the species, which means that there are no antimers and no metamers, which in turn implies that no fifth- and no sixth-order morphological individualities are involved. So indeed the colony of Volvox is tectologically an organ.
Organs as physiological unities, Organs as morphological unities
In our consideration concerning organs it should again be stressed that the term ORGAN does have two meanings : The usual meaning (and also its literal one) is that of a working unit, i.e. a unit that is geared to perform a certain function, like the bird's wing, its heart, its lungs, etc. Another meaning is that of a form unit, i.e. one of the building blocks of the organism's body. In the latter sense it is a second-order morphological individual. Our concept of an organ as PHYSIOLOGICAL INDIVIDUAL (biont) also means a second-order morphological individual, but now one that is not actually subordinated to any higher morphological individuality, but is a subsisting entity, which means that it can maintain itself by its own actions. So here the unity not only refers to a coherent spatial entity, but also to a coherent energetic entity, an work entity, that should be considered during a certain time span in which it often is able to let parts of it split off without losing thereby its condition of a coherent energetic unity.
Let us work out all this, in order to better understand the meanings of the concept of ORGAN.
In every organism, which belongs to a higher-order morphological order, it is easy to recognize as such the subordinated individualities, the metamers, antimers and cells. As ORGANS we will accordingly denote all those uniform form appearances that do not belong to one of the other five orders of individuality. In the tectologically so distinctly determined and differentiated higher plants (phanerogams), vertebrates, articulates, etc. we will not entertain doubts as to which parts of them are organs and which not. However this distinction becomes more difficult in many lower forms of both kingdoms, especially in cryptogamic plants. Especially in these cases it is paramount to distinguish between the morphological and physiological concepts of organ. Only the first gives us the second-order form individual.
The o r g a n a s p h y s i o l o g i c a l u n i t y can be an integrating constituent of form individuals of all six orders, and means nothing than a body part which performs a certain function (or is able to do so). So is, for example, a cilium (i.e. a hair that can vibrate) of a ciliated cell (i.e. a cell provided with such hairs, cilia) an organ of that particular cell, and also the stinging thread and capsule of a nettle cell. In every complex organ the subordinated parts, which are cell complexes, are at the same time 'organs of the organ' (On this phenomenon our distinction of the five orders of organs was based). Every antimer is composed of several organs, but can itself be an organ of a metamer, for example the arms of the starfishes. In the same way one can, from a physiological viewpoint, interpret the metamers as 'organs of the person' (fifth-order form individual), and the persons (off-shoots) as 'organs of the colony' (cormus), as we see in articulate worms and in most higher plants respectively. In all these cases the idea of the physiological performance is central when we use the word ORGAN, the idea that expounds the organ as an integrating constituent of a body.
Matters are wholly different with respect to the m o r p h o l o g i c a l c o n c e p t o f t h e o r g a n. This concept invariably means only such an indivisible form unit of constant size and composition, comprising a multitude of cells, which neither can be assessed as an antimer nor as a metamer, neither as person nor as cormus. Such form units indeed not only commonly occur as constituents of all morphological individuals of third to sixth order, but also occur, as physiological individuals, wholly independently, more seldom to be sure as actual bionts, but very commonly as virtual, and sometimes in very conspicuous forms as partial boints. Of the five different orders of organs, that we have distinguished in the section on second-order morphological individuality, the first- and second-order organs (cell fusions and simple organs respectively) occur most often as bionts, more seldom those of third order (composed organs). The fourth- and fifth-order organs (organ systems and organ apparatus respectively) on the other hand, can, according to their nature, never obtain the status of physiological individuals.
As long as one confuses the quite different physiological and morphological meanings of the concept of ORGAN, one will not arrive at a clear idea concerning the important fact that also morphological organs can occur as actual bionts, and as such represent the mature form of independent species. According to us this is the case in all those lower organisms, that in their mature form appear as a cell complex, in which neither antimers, nor metamers can be distinguished, and which are consequently neither persons nor cormi. To this belong very many protists and lower plants, in particular all so-called "colonies of unicellular organisms". Anyway, as has been said, only first- second- and third-order organs can function as actual bionts. The fourth- and fifth-order organs are intrinsically connected and intermingled with other structures within the organism, so they cannot exist for themselves in isolation (expressing also their lower degree of individuality). They cannot function as bionts.
The above depicted Diatomean colonies belong to the first-order organs, the form and shape of such a colony is solely determined by the constituent cells and their way of attaching to each other.
Second-order organs have their form and shape partly determined by the whole of which they are a part, so in a way they can, as such, not be distinguished when existing on their own, i.e. as bionts. But, in a certain way they can be so distinguished, namely where the cells have not only just aggregated and attached to each other but also have been changed, i.e. more or less deformed, in order to adapt to a particular overall shape of the colony, as we see it in Volvox (Figure 8). So the Volvox colony is an actual biont having the morphological status of that of a second-order organ. The same is the case in all multi-nucleate Foraminiferans.
Third-order organs or composed (heteroplastic) organs, i.e. cell complexes that consist of an aggregate of dissimilar differentiated cells, and which do not have the positive morphological characters of antimers or other higher-order form individuals, we see appear as bionts in lower organisms, for instance in many Thallophytes (algae and the like) which display a wholly irregular shape, and which are composed of dissimilar cells. Also many sponges are third-order organs appearing as bionts.
As virtual bionts of second order, which accordingly are subsumed under the morphological concept of organ, we consider all those independently living cell complexes, which do not display the positive typical form features of third- to sixth-order form individuals, and which are moreover able to develop into one of these higher form individuals. These conditions we find to be realized in all those multicellular developmental stages of higher organisms, which are not yet differentiated as to display antimers, for example in the first embryological state in vertebrates. In higher vertebrates this embryological state is still inside the parent organism, and is as such to be judged as biont only in a qualified way. In lower vertebrates, on the other hand, namely those which release their eggs to the environment, we have to do with genuine bionts. When these eggs have developed into the mentioned first embryological state, we have to do with virtual bionts of second order. Further there are organisms, like Hydra and other hydroid polypes, that have cell groups, that, when detached from the main body, can develop into actual bionts. So such a detached cell group is indeed a virtual biont of second order. (We will later add some (more) examples, by consulting modern literature).
As partial bionts of second order we consider all cell complexes possessing the morphological value of organ, which can, separated from the relevant organism, independently live for a longer or shorter period, without however to be able to develop into the actual biont. To this belong all independently existing so-called "individualized organs". (We will later add some examples, by consulting modern literature).
In yet a stronger degee, as with respect to organs, it would be peculiar to see antimers, that as physiological individuals living a life independently. Indeed indisputable cases of physiological individualization are here much more seldom, and with respect to actual bionts it is even doubtful whether physiological individuals can ever be represented by antimers. Wide-spread however are cases of virtual individualization, and (Haeckel, 1866) in many organisms also of partial individuation.
When we keep in mind that the antimers or counterparts, as third-order form individuals, are in fact only as such determined by their mutual relationship and by their relationships to the fourth- and fifth-order individuals which they compose, it should appear doubtful beforehand whether they ever can appear as actual bionts. Haeckel (1866) reports that indeed in all three organic kingdoms no indisputable case is known in which the mature form of some species is represented by a form individual which for certain has the value of a single antimer. Most species representatives either are composed of two antimers ("body halves") or of three or more antimers ("radial parts"). Those cell complexes, however, which are, as actual bionts, an aggregate of differentiated cells, without appearing to be composed out of two or more antimers, we will more legitimately assess as second-order bionts, than as third-order bionts. Only then we can ascribe actual bionts as having the value of antimers, when they were complete morphological equivalents to parts that in related species can be considered as indisputable antimers of a metamer or of a person. Maybe certain species of Lemna could be assessed as actual antimers.
While the actual bionts probably never, the partial bionts only seldom, are represented by antimers, the potential or virtual bionts are so represented very often. To this we reckon all cases of reproduction by spontaneous self-division and by artificial (accidental) division, in which the plane of division divides the body of an actual biont into its antimers, and where the single antimers immediately complement themselves, i.e. restore the whole, by the reproduction of the remaining antimers. When a flatworm is cut into two parts along its median plane, then each single antimer restores itself again, resulting in a whole, again consisting of two antimers (left and right body halves). See the next Figure.
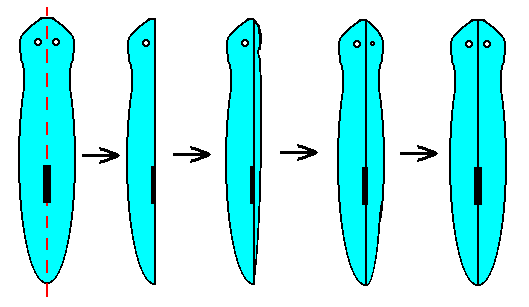
Figure 9. The regeneration of a complete flatworm ( of the genus Planaria ), cut as indicated on the left.
( Redrawn after SHELDRAKE, R., 1988, The Presence of the Past (Morphic Resonance and the Habits of Nature), Figure 5.3, p. 77. )
Single pieces of higher organisms, which, separated from the whole, are able to live independently, without being able to develop (again) into the whole, appear, as we saw, often in the form of organs, but seldom in the form of antimers. One can assess as (indeed representing) such partial bionts having the form value of single antimers, for example single detached arms of starfishes, arms that include the relevant section of the central disc, and that are, under certain circumstances, able to live independently for a longer or shorter period, without however being able to develop wholly into the actual biont. But these cases are seldom, and without great significance. Also in some hydromedusae this phenomenon occurs (Haeckel, 1866).
To continue the study of physiological individuality click HERE
back to homepage
back to retrospect and continuation page
back to Internal Structure of 3-D Crystals
back to The Shapes of 3-D Crystals
back to The Thermodynamics of Crystals
back to Introduction to Tectology
back to Tectology, cells, organs, antimers
back to Tectology, persons and colonies