As first-order morphological individuals, representing the lowest order of organic individuality, we must interpret all b i o l o g i c a l c e l l s, either as free-living (In that case they also represent the physiological individuum of the given organic species), or as subordinated form individual, taking part of a multicellular organic body.
There are two main types of cells, (1) prokariote cells, i.e. cells without a cell nucleus, and (2) eukariote cells, i.e. cells possessing a nucleus. Only these latter can form true cell tissues, and thus can ascend the tectological ladder.
All organisms begin their existence as a single cell. In the overwhelming majority of organisms this cell splits up, its daughter cells remain adjacent to each other, they grow and split up again. This process finally leads to the multicellular organism, the final, i.e. mature, physiological individual or actual biont. This actual biont can be an individual of one of the five subsequent orders of individuality. In other cases, namely in unicellular organisms, the physiological individual got stuck at the first-order of individuality, i.e. it remains one isolated free-living cell.
As has been said in the introductory document, the c e l l is the unit of Tectology, and that's the reason why Tectology does not in principle investigate the built-up, i.e. the constitution, of the (single) cell.
Paramers and Epimers
In an a n a l o g i c a l sense, however, Tectology can consider the global morphological constitution of the cell, and distinguish accordingly between the cell's paramers and epimers when present. These latter are in general discussed and defined in the sections concerning Antimers (third-order form individuals) and Metamers (fourth-order form individuals) respectively, and at the end of the section concerning Organs (second-order form individuals). With respect to the cell (a first-order form individual) we can succinctly define them as follows :
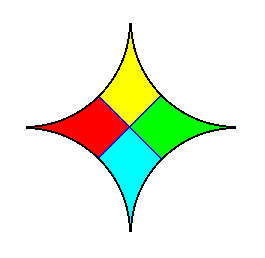
A p a r a m e r of a cell is a subordinated form unit that is repeated either a r o u n d a point, i.e. about the center of an endospherical cell, or a r o u n d the cell's main axis (in a pyramidal cell), or around a plane in a strictly bilateral-symmetric cell (See for further explanation : the end of the treatment of organs).
An E p i m e r of a cell is a subordinated form unit that is repeated a l o n g the main axis of the cell. See next Figures.
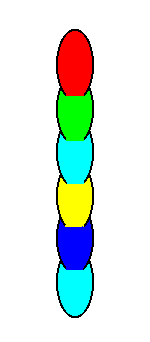
The situation in Figure 2 is exemplified (among others) by the genus Nodosaria of the Foraminiferids.
In Promorphology -- which follows upon our treatment of Tectology -- the distinction between paramers and antimers is not important, so there we will commonly call them a n t i m e r s.
The physiological individuality of the organismic body is limited to the first-order morphological individuality in many lower organisms, without ever elevating itself towards higher orders. And as soon as a multiplication of cells takes place, a corresponding multiplication of physiological individuals is implied there. These individuals then lead each for themselves an independent existence.
In the overwhelming majority of those organisms that today inhabit the earth on the other hand, the physiological individuality rises above the order of the single cell, the first-order form individual, because now several cells get together into a social relationship, which now appears as a higher physiological unity. In this way the several higher-order individuals originate, which we distinguished above as Organs, Antimers, Metamers, Persons and Colonies.
The most essential and highest laws, that regulate this association of simple and single first-order individuals into composite entities, are the laws of a g g e g a t i o n or socializing, and of d i f f e r e n t i a t i o n or division of labor. At first several similar cells aggregate (ideally understood, which implies that socialization can also take place by means of repeated cell fission), resulting in a companionship that is homogeneous with respect to its constituents. The increase in ability or power, i.e. the physiological perfection, characterizing this companionship of similar cells as a higher unit, is at first just a quantitative growth of power. But, gradually, from this q u a n t i t a t i v e perfection by a g g r e g a t i o n, originates the much more important q u a l i t a t i v e perfection in virtue of d i f f e r e n t i a t i o n. At first namely, very small, later, however, more significant differences appear among the originally similar cells, finally resulting in a complete division of labor. And while the constituent cells, in virtue of this process, give more or less up their individuality, i.e. their individual independence, and enter in the service of the higher unity, the cell colony, they develop characteristic features one-sidedly in certain directions, and so complement and depend on each other.
The concept of o r g a n or work-part originally is a pure physiological concept, so a vindication is demanded when we (also) use this concept to indicate second-order morphological individuality. At first such a vindication is already evident by the fact that the performances of every organ (or tool for that matter) are only partially determined by its chemico-physical properties, while at the same time they are, and often for the most part, determined by its form (shape) and by its inner structure, which lies at the base of its outer shape.
Definition of an o r g a n in its m o r p h o l o g i c a l sense.
The morphological concept of an o r g a n in general is, according to our view, determined as a constant uniform spatial magnitude having a definite form (shape and structure), which consists of a number of cells, that are socialized by a constant connection, and which does not possess the positive features of the third-, fourth-, fifth- or six-order form individuals.
This morphological definition of an organ may sound poor, because of its partial negative content, but will, because of the extraordinary diversity of the different organs, not easily be replaced by a better one that is still generally applicable.
Having determined the morphological concept of an organ, we must, at least in order to obtain a general overview of the many ways in which organs are constituted, attempt to distinguish different orders of organs on the basis of a higher or lower degree of composition. Such an establisment of different orders of organs is not easy, partly because of the enormous diversity of organs, but also because we are used to evaluate them according to physiological features and criteria, which means that we must free ourselves from this habit in order to obtain a morphological and generally applicable classification of organs.
With respect to such a classification attempt we must consider the existence of two fundamentally different types of organic cells : p r o k a r i o t e c e l l s, i.e. cells that do not possess a cell nucleus, and e u k a r i o t e c e l l s, i.e. cells that possess one. Only the latter can form true tissues. On the basis of this one has proposed to divide the organisms not into Protists, Plants and Animals, but into Prokarionts (Anucleobionts, organisms, the cells of which lack a nucleus, as in Bacteria and Blue Algae (Cyanophyceae, today also called Cyanobacteria) on the one hand, and Eukarionts (Nucleobionts, organisms with their cells having a nucleus, like in all other organisms). In addition to these two main categories of organisms a third category could probably be represented by the Methane Bacteria (KAPLAN, R., 1978, Der Ursprung des Lebens, p. 236). The prokariote cells or procytes (Haeckel called them Cytodes) are different and much weaker internally differentiated than the eukariote cells or eucytes (Haeckel called them just Cells). While the procyte contains the genome ( = the structure containing the genes) nudely in its central cytoplasm, where this genome consists of a DNA ring -- as "nucleoid" -- (analogous to a nucleus), the eucyte possesses several to many chromosomes consisting of DNA, RNA and proteins. They are separated from the cytoplasm by a porous bi-membrane, resulting in a genuine cell nucleus (KAPLAN, op. cit.). From a morphological viewpoint we can pose that the nucleus, where it is present, determines the individuality of the cell, which means that when we have to do with so-called multinucleate cells, we, morphologically have to do with a multicellular condition, i.e. with an organ in the morphological sense. Indeed do we know of such a condition, for example in Foraminifera, primitive minute shell-bearing marine animals. The endoplasm (i.e. the cytoplasm that is found within the shell) contains food vacuoles and the nucleus. Although a uninucleate condition may be found in the simplest forms, many nuclei occur together within the single cells of many-chambered Foraminifera, i.e. a multinucleate condition (BRASIER, M., 1980, Microfossils). So if ever we have to do with a foraminiferid possessing more than one nucleus in its cytoplasm, then it should be morphologically interpreted as an o r g a n, i.e. as a second-order form individual.
On the basis of the degree and character of composition and of the determinants of their forms we will classify the o r g a n s in five morphological categories as follows :
The organs, that we consider as the most simple morphological organs, i.e. as first-order organs, are relatively seldom materialized in the organic world. Among its representatives we can reckon all more or less permanent "multinucleate cells", for example the above mentioned foraminiferids possessing more than one nucleus in their cytoplasm.
Definition of a first-order organ.
The specific character of the first-order organs and their distinction from the simple organs or second-order organs rests on the fact that the connection of similar cells already in and by itself determines the formation of a definite morphological unity. The form of such an organ accordingly is independent of the global morphological constitution of the whole organism, and the organ is with respect to its special features solely dependent of the specific quality of the intimately connected cells.
When we would have a case of fusion of prokariote cells, the result would lack any morphologically criterium that allows such a fusion to be assessed as a complex of cells. In colonies of eukariote cells on the other hand, we do have such a criterion as long as the nuclei still persist, even when the walls of separation have disappeared, because the nucleus of the cell determines its individuality.
(Cell socializations consisting of one sort of cells : Homogeneous cell complexes)
As second-order organs we consider "simple organs" that can perhaps more definitely be named as h o m o p l a s t i c o r g a n s.
Definition of second-order organs or homoplastic organs.
The homoplastic organs share with the first-order organs, the cell fusions, the fact that they are composed of similar cells, i.e. of cells of one and the same sort. They differ however from the cell fusions by the fact that the form of the organ is not first of all determined by the connection of the constituent cells, but is, apparently independently from it, determined by the constitutional relationships of the whole organism, i.e. by its general structural plan. Accordingly homoplastic organs can look quite different despite the fact that they are composed of only one sort of cells, while the cell fusions or first-order organs, also composed as they are of one sort of cells, wherever they appear within one and the same biological species, almost always appear in the same form, in virtue of the constant way of connection of their constituent cells. In these first-order organs the external form of the organ is in itself already determined by the mode of connection of the constituent cells, while this form is in the case of second-order organs independent of the mode of connection. It is dependent upon the global structural features of the whole organism.
The cells, that compose homoplastic organs, can be prokariote as well as eukariote. An intimate connection of the elements, where it occurs, can be brought about by the lack of cell membranes, but such a connection is in itself without special significance with respect to the appearing form. The latter is indifferent as to whether the organ has originated by total fusion and disappearing cell walls, or just by aggregation or multiplication and retention of cell walls or the presence of intercellular substances, resulting in a relatively independence of the constituent cells.
An example of a homoplastic organ in Vertebrates is the total upper skin (epidermis) including all its appendages like hairs, nails, scales, glands etc.
(Cell socializations consisting of different sorts of cells : Heterogeneous cell complexes)
The overwhelming majority of all organs in higher organisms, animals as well as plants, are heteroplastic organs.
Definition of heteroplastic organs.
Heteroplastic organs or third-order organs do not consist of just a single sort of cells, but of several different sorts, in virtue of the connection or socialization of several differently differentiated cell complexes, whether they be first-order organs or second-order organs, a connection that builts up a third-order organ or heteroplastic organ. The division of labour, that appears among the initially similar cells, enables the elements, together making up one whole, to realize higher performances.
In the great majority of animals the composition of most organs out of several tissues, i.e. out of more than one sort of cells, is already determined by the fact that in the early stages of individual life an ongoing differentiation takes place of the initially similar cells, and that in virtue of the differentiation of tissues, on the one hand very diverse composed organs, and on the other hand specific relational organs or centralizational organs are generated, that bring the several other organs into more or less close connection with each other and with the central organs. Such a relational organ in the animal body is the nerve system, another the vascular system that feeds the body. Further a spatial connection and at the same time a partition from neighboring organs is effected by the various connective tissues. All these organs that pervade the whole body of the higher animals send their twigs into the interior of most other organs, where they extend among their constituent tissues. In the same way the whole body of higher plants is pervaded by vessels, that penetrate everywhere into the axial organs and leaf organs, that are made up of simple tissues. In this way those vessels acquire the nature of heteroplastic organs.
The great majority of organs, at least in higher plants and animals, thus is c o m p o s e d insofar as they are not only composed of the cells of the specific tissue types that execute their specific functions, but also as they contain branches of the nervous system or branches of the vascular system, which connect them with the rest of the organism, branches of the vascular system that feeds them (i.e. those organs), branches, etc. of the connective tissue that props them, encloses them and connects them with neighboring organs. The organs that are in this way composed are the individual muscles, nerves, bones, vessels, etc. in animals, while they are the individual leaves and the various axial organs in higher plants.
The difference between those composite organ complexes that are called s y s t e m s, and those that are called a p p a r a t u s, essentially rests on the fact that the first nomination is morphologically motivated while the second is so physiologically. In the case of an organ system one thinks of the u n i t y o f i t s f o r m, despite its compositional nature, while in the case of an organ apparatus one thinks of the unity of its performance in spite of its compositional nature. Nevertheless the concept of organ apparatus can be given (also) a morphological sense.
Definition of an organ system.
Every single o r g a n s y s t e m or fourth-order organ is a morphological unity, composed of a multiplicity of composed organs, in which however moreover also simple organs and cell fusions can enter as constituent elements. A single sort of cell, i.e. a single sort of tissue, is however predominant in every organ system, and the remaining tissues that also go into the constitution of the organ system are subordinated, morphologically as well as physiologically, with respect to the typical tissue (i.e. cell sort) of the system : with this we have the essential difference from the organ apparatus, in which several systems can appear as coordinated constituents alongside each other, whithout there being a preponderance of one of them.
As organ systems in this sense we can in Man, and generally in Vertebrates, consider : skin system and all its appendages, skeleton system, muscular system, nervous system, vascular system, intestinal system and genital system.
In general the organ systems are further developed and differentiated in animals than in plants. The further the centralization of the person has proceeded, the further is the separation or isolation of the single organ systems, the more unified each system is. In the higher plants one could perhaps indicate as such coherent systems : the surface system (skin, bark, etc.), the parenchym system, the vascular system.
Although the concept of apparatus is based on physiological consideration, implying that one normally understands by it a complex of simple and composed organs, aggregated into one uniform whole in order to be able to perform a common function, we nevertheless can provide the concept of apparatus (also) with morphological content.
Definition of an organ apparatus.
An organ apparatus or fifth-order organ is a uniform self-contained complex of several different subordinated organs (belonging to various systems and tissues), in which various coordinated tissues are gathered such that not any one of them can claim a predominance with respect to all other tissues.
These apparatus indeed deserve to be considered as high-order organs because the morphological unity is not less, often even more expressed as in the various systems. The unity of form in apparatus is, however not based, as in systems, on the preponderance of one tissue type, but much more on a spatial isolation, that is directly evident even by a global observation. So the visual apparatus, for example, is limited to the content of the eye-ball, the sense of smell to that of the nasal cavity.
Because in addition to the spatial isolation and unity also the morphological composition of every organ apparatus out of coordinated and subordinated organs effects a uniform expression of the apparatus, we can, basing ourselves on the intimate interaction of form and function, just like we did with respect to the concept of organ in general, distinguish the morphological unity of the apparatus from its physiological unity, and then consider the morphological unity as a fifth-order organ.
All organ apparatus, which one can further classify in lower or special and in higher or general, allow a division into three main groups, corresponding to the three main organic functions : Self-maintenance, Continuation of the species and Maintenance of the relations to the environment.
According to this we can distinguish in Man, and generally in Vertebrates, the following groups of organ apparatus :
Paramers and Epimers
Some o r g a n s (of any order) can, like we saw in cells, (at least) in principle show a regular repetition of parts analogous to antimers (next to be discussed) and metamers (discussed after antimers have been considered). They are however not genuine antimers or metamers.
Antimers are regular repetitions a r o u n d the body's m a i n a x i s, while metamers are regular repetitions a l o n g the body's m a i n a x i s.
With respect to a n t i m e r s matters are somewhat more complicated :
Left image (I) : Base of a regular five-sided pyramid. It represents its five antimers (one of them indicated in blue). O is the center of the axial system. The pyramid's main axis is perpendicular to the plane of the drawing and goes through O. It is the pyramid's center.
aa', bb', cc', dd', ee' indicate the five cross axes.
aO, bO, cO, dO, eO indicate the five radii.
The next Figure (III) depicts a body (a pyramid, represented by its base) that also has a plane as its center, but having only two antimers.
III : Base of a pyramid having two symmetric antimers (indicated by blue and yellow). The pyramid's main axis is perpendicular to the plane of the drawing and goes through O. The center of this pyramid is the plane that contains the main axis and the (interradial) axis Ot. This plane is at the same time the median plane of the pyramid. The two antimers lie on either side of the median plane. They have this plane in common. They lie, as it were, around this plane, i.e. around the body's center.
M e t a m e r s on the other hand, are more easy to understand. They are only present in organic bodies having their center represented either by a line (Centraxonia) or by a plane (Centrepipeda), i.e. in organisms that clearly show a distinct main axis (Protaxonia). The metamers are then form units that are repeated along the main axis of the body.
In Promorphology (which will follow after our treatment of Tectology) we will learn that (organic) bodies can be divided into three main groups according to the character of their c e n t e r (See also the Figures below) :
The next Figures (I, II, III) illustrate some of the above mentioned features. The images represent pyramidal bodies by means of drawings of their respective bases (or, equivalently, their equatorial planes). The tip of these pyramids should be imagined to lie precisely above the point marked by O.
[ Of course, where more than two antimers are present, they also lie on either side of the median plane, but in this case half of the antimers lies at one side of it, the other half at the other, while not having this plane in common. All of these antimers lie around the main axis (which they do have in common), while this "lying around the main axis" is less clear in the case where only two (symmetrical) antimers are present ].
Figure I represents the Centraxonia (body center is a line), while the Figures II and III represent the Centrepipeda (body center is a plane).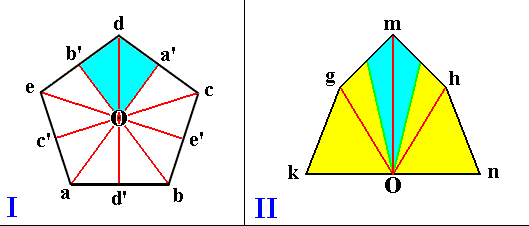
Right image (II) : Base of a pyramid consisting of three antimers, one unpaired antimer (blue) and two symmetric antimers (yellow). The pyramid's main axis is perpendicular to the plane of the drawing and goes through O. The center of this pyramid is the plane that contains the main axis and the (radial) axis Om. Og, Om, Oh indicate the radii. The three antimers lie around the main axis (O).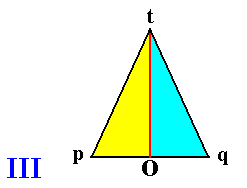
All this is extensively explained and worked out in the documents on Promorphology, where a n t i m e r s play a crucial role.
The p a r a m e r s and e p i m e r s are similar to, respectively, antimers and metamers, but generally relate to a cross axis of the body instead of to its main axis. Equivalently we can say that paramers and epimers of an organ relate to the organ's main axis.
Also this needs some qualification.
Like any organic (whole) body, also o r g a n s can promorphologically be of three types.
Organs, the center of which is represented by a point (centrostigmatic organs).
Organs, the center of which is represented by a line (centraxonic organs).
Organs, the center of which is represented by a plane (centrepipedan organs).
The paramers of the organ relate in the same way to the organ's center as do the antimers to the center of the whole body.
Epimers in organs only occur in centraxonic and centrepipedan organs. They then relate to the main axis of the organ as do the metamers to the main axis of the body.
Let us now define paramers and epimers with respect to organs :
A p a r a m e r of an organ is a subordinated form unit that is repeated either a r o u n d a point in the organ, i.e. about the center (central point) of an endospherical organ (centrostigmatic organ), or a r o u n d the organ's main axis, i.e. in an organ having a pyramidal basic form (centraxonic organ), or around the plane that is the center of the organ, i.e. in an organ of which the basic form is generally expressed by half an amphitect pyramid (centrepipedan organ).
An E p i m e r of an organ is a subordinated form unit that is repeated a l o n g the main axis of an organ. See next Figures.
An example of paramers of an organ could be : the two halves of many plant leaves, especially those of dicotyledone plants : The leaf itself is an organ, while its two halves are its t w o paramers.
The midline of a (dicotyledone) plant leaf (indicated by its main vein) can conveniently be interpreted as its m a i n a x i s. The l a t e r a l a x i s of the leaf can unambiguously be recognized as connecting the two symmetrical halves (paramers) of the leaf. It is the only axis that is equipolar. The d o r s o - v e n t r a l a x i s of that leaf is now implied : it is perpendicular to the main axis and at the same time perpendicular to the lateral axis. The two paramers can be considered as to lie around the leaf's main axis. More precisely, they are symmetrically repeated around the median plane (which contains the main axis and the dorsoventral axis). This median plane coincides with a cross plane of the off-shoot to which the given leaf belongs. This off-shoot is itself a higher-order individual, namely a fifth-order form individual, a person (in most higher plants a multitude of such persons compose a colony, a six-order individual, like, for instance, a tree). See next Figure.

Figure 5. An organ ( a leaf of the genus Ficus ), consisting of two paramers. The lateral axis of the organ is indicated by a red line. The central vein indicates the main axis. The dorso-ventral axis is perpendicular to the plane of the Figure and goes through the point of intersection of the main axis and the lateral axis. The median plane of the leaf is perpendicular to the plane of the Figure and contains the main axis and the dorsoventral axis.
The choice of the main axis, with respect to the above case, is however not compulsory. We can choose the axis that is perpendicular to the leaf's surface as being the main axis of the leaf. The axis along the main vein then is the dorso-ventral axis. Also in this case we can still say that the two paramers are grouped around the main axis of the leaf. This interpretation of the axes is strongly suggested by the situation in Figure 9, where we have to do with a five-fold leaf in which the main axis is already totally determined as to which one of the possible axes it should be.
To illustrate schematically the relationships between antimers and paramers, and between metamers and epimers, we depict an asexual off-shoot (i.e. an off-shoot not terminating in a flower) of a dicotyledone plant. Such an off-shoot (but also any sexual off-shoot) is composed of metamers, indicated by stem nodes and leaf rings, and of antimers. It must therefore be considered as to represent a fifth-order form individual, a p e r s o n. The main axis of such an off-shoot runs through its stem, while its cross axes are perpendicular to it and indicate the radii, which are materialized by leaves. The a n t i m e r s group themselves around the main axis (they are indicated by the leaves). Each leaf half is a paramer.
When the leaf is feathered, as we see in ferns, then e p i m e r s are apparent : The leaf then is partitioned into epimers, i.e. into repeated morphological units a l o n g the main axis of the leaf. This main axis lies in a cross plane of the off-shoot and can represent a (radial) cross axis. See next Figures.
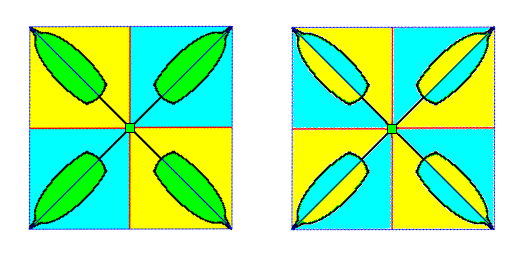
Figure 6. Diagram (containing two images) of an asexual four-fold off-shoot of a dicotyledone plant. It is seen from above, i.e. the direction of view is parallel to the main axis of the off-shoot. The images in the Figure in fact depict a projection of the off-shoot onto a plane perpendicular to the main axis.
Left image : Indication of the four a n t i m e r s of the off-shoot.
Right image : Indication of the eight p a r a m e r s. Each leaf consists of two paramers.
The four antimers of the off-shoot are all congruent, and thus are exactly repeated (around the main axis of the off-shoot). The eight paramers are only two by two congruent, and so are not exactly repeated around the main axis. So also they express the four-foldness (not the eight-foldness) of the off-shoot. Each leaf on the other hand consists of only t w o similar parts. In this case they also are not congruent. They are symmetric. So they are symmetrically repeated around the leaf's main axis, and constitute the leaf's paramers.
So antimers are subordinated morphological units of fourth- or fifth-order form individuals, while paramers are subordinated morphological units of first- or second-order form individuals.
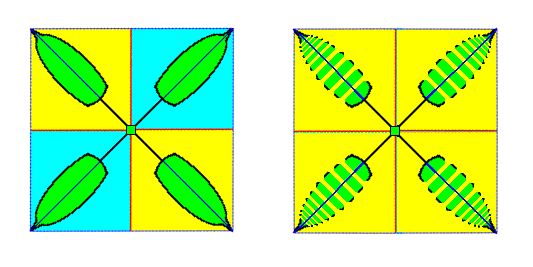
Figure 7. Diagram (containing two images) of an asexual four-fold off-shoot of a dicotyledone plant. It is seen from above, i.e. the direction of view is parallel to the main axis of the off-shoot. The images in the the Figure in fact depict a projection of the off-shoot onto a plane perpendicular to the main axis.
Left image : Indication of the four antimers of the off-shoot.
Right image : Indication of e p i m e r s. Each leaf consists of a number of epimers that are repeated along its main axis.
In the next section -- considering antimers (third-order form individuals) -- we will again discuss paramers, and in the section that considers metamers we will again discuss epimers.
As already been said (when treating of paramers (and epimers) of cells), we will not distinguish between paramers and antimers in Promorphology. There we will commonly call them a n t i m e r s.
The previous consideration of the first- and second-order morphological individuals, the cells and the organs, has, while having to overcome serious problems, introduced us to the complicated labyrinth of coordinated and subordinated parts out of which the whole organism of higher animals and plants is composed and in this way resulting in a higher unity. A precise consideration of the utmost complicated and 'artistic' way in which this composition takes place, immediately shows that the step-wise increasing complication of the organic built-up, at least in higher plants and animals, is accomplished not only by the general laws of aggregation and differentiation, but that the various coordinated and subordinated parts in such a way intertwine in the whole, mutually penetrate and connect spatially, and in such a complicated way intermingle, that we end up in recognizing wholly different morphological units, depending on different standpoints, and from this or that common tertium compair two units. So it is possible that the same nerve, the same muscle, will be assessed as a complex of simple organs of first and second order, or as one heteroplastic organ, or as a part of an organ system, or as a part of an organ apparatus. And from each of these points of view it will be differently judged.
This was reported by Haeckel in 1866 (Generelle Morphologie, p. 304 ). Unfortunately, to my knowledge, this still stands today. Each organism group is treated by specialists who use their own terminology appropriate, as they deem, for the given group, and who investigate special problems connected with that group. Of course, specialism cannot be avoided and is necessary, but it is pitty that one does, as it seems, not try to integrate the morphological features of whatever animal or plant group into a g e n e r a l morphology (that is composed of a general theory of organic macroscopic structure -- Tectology -- and of a general doctrine about the stereometric basic form of organisms and their macroscopic and submacroscopic parts -- Promorphology).
Those parts of the organismic body, that we will distinguish as a n t i m e r s or counterparts, and m e t a m e r s or sequential parts, are sharply expressed morphological individualities, that rank above organs, while they are invariably subordinated to the higher morphological units of fifth and six order. In the overwhelming majority of organismic species the single physiological individuum is not just an aggregate of organs, but a unit consisting of several metamers and antimers. This means that here we have to do with r e p e t i t i o n s o f m a c r o s c o p i c a l p a r t s. For the overall f o r m of the organism these parts, that appear as a multitude of definite form units behind or next to each other, are of the utmost importance. But still, relevant studies, incorporating them in a general way, i.e. trying to trace them throughout the organismic world, are almost absent. The s e q u e n t i a l parts (parts lying behind each other) as they can be detected in the trunk of Vertebrates and Articulates, or in the stem of higher plants, and which we will call metamers, are often called "segments", "rings" or "zonites". The antimers on the other hand, i.e. main parts that lie n e x t to each other, were, when only two are present as in Vertebrates, Articulates and Molluscs, called "body halves", and when there are three, four, five or more of them, as in the radiate animals (medusae, starfishes, etc.) and in flowers of higher plants, they were called "radii" or "radial segments". Before Haeckel undertook a study of these parts from a general view point, only BRONN (Morphologische Studien, 1858) had done so. We will later reproduce Haeckel's discussion concerning BRONN's results (because they are very instructive), in the section devoted to metamers.
Definition of Antimers.
With a n t i m e r s o r c o u n t e r p a r t s we mean those body parts that appear as definitely self-contained units, and that lie n e x t t o each other (not behind each other), and which, as equivalent organ complexes contain all or almost all essential body parts of the biological species (all typical organs), in such a way that each antimer represents the most essential features of the species as a complex of organs, and that only the n u m b e r of antimers, as species determinant, is added.
In most so-called bilateral-symmetric animals (Vertebrates, Articulates, Molluscs) the body accordingly is composed of t w o antimers, i.e. both body halves, that are fused in the body's median plane. In the radiate animals as well as in most sexual individuals (flowers) of higher plants, the body is composed of that number of antimers determined by (and equal to) the number of radii, i.e. cross axes, that are present in that body : t h r e e in most monocotyledone plants, f o u r in most medusae and most worms, and in many dicotyledone plants, f i v e in most (adult) Echinoderms (starfishes, sea-urchins, and the like), s i x in most corals. Very seldom we will encounter organismic bodies that are composed of more than six antimers. E i g h t antimers are nevertheless seen in all Ctenophores (sea-gooseberries or comb-jellies). Very seldom we find nine, ten, twelve and twenty or more antimers composing an organismic body. Generally the lower numbers of antimers are constant within the biological species. As soon as more than six antimers occur, that basic number (except the number eight) becomes variable within the species, and the more unstable the higher this number is (Haeckel, 1866). All this also occurs in the case of metamers, for example when we compare Insects, having a relative small but constant number of metamers, with Myriapods, having a large unstable number of metamers, in this respect (Haeckel, 1866).
So insignificant it may be, from a p h y s i o l o g i c a l viewpoint, whether the whole body (the person) consists of two, three, four, five or more similar body parts, of which each one contains all essential organ complexes or typical organs of the body, and accordingly could represent already the species, so utterly significant is that h o m o t y p i c b a s i c n u m b e r -- as Haeckel calls (with BRONN) the specific number of a n t i m e r s -- for the m o r p h o l o g i c a l consideration of the body as a whole. Especially the antimers determine that total of form features that is commonly denoted as h a b i t u s, and which often is as hard to define and to determine more precisely, as it presents itself (easily) p h y s i o g n o m i c a l l y as character-determining to the experienced eye.
It's true that the causal connection between the typical organizational character and the homotypic basic number in organisms is unknown to us. Macroscopical organization, i.e. organization involving supra-molecular distances, is probably explainable with the help of dissipative (chemical) dynamical systems, in which a large number of elements interact in a coherent way, resulting in so-called emergent phenomena, not visible at lower scale levels [ We discussed these dissipative systems in one of the Essays of the Non-Classical Series (of Essays) of this website. To adress this Essay, click on back to homepage at the end of the present document, and then click on "Non-living Dissipative Systems"]
That such a causal connection between the typical organization prevailing in a certain group of organisms and the occurring homotypical numbers, is indeed present, is proved by the remarkable constancy of the number of antimers within the large main divisions of the animal and plant kingdoms. Without exception the Vertebrates an Molluscs are composed of only two, the Ctenophores of eight antimers, and predominantly the number five reigns in Echinoderms, and the number three in monocotyledone plants. All this is certainly not without significance, and it demands a closer look to the mutual relationships of the antimers to each other and to the whole. With respect to the latter we must first of all note that in determining the h a b i t u s of the organism, the antimers can either be totally e q u a l, or only s i m i l a r, and in the first case either s y m m e t r i c a l l y equal, or c o n g r u e n t.
As being s i m i l a r we call antimers, if they are, to be sure, equal to each other with respect to all, or in any case most, essential form relationships and possessing the same number of the larger organs that are connected onto the same relative position, but nevertheless more or less differ in subordinated features, in their size, the stronger or weaker development, in their surface features, etc., implying that also the number of the smallest parts that compose them, is significantly unequal. So both body halves of a flatfish (Pleuronectes) are similar (not at all equal!). Similar is the unpaired radius of the symmetric (pentamphipleural) Echinoderms with respect to the four other radii.
E q u a l, on the other hand are two homotypical parts (antimers), when they are equal not only with respect to those essental parts, but also when they are wholly equal with respect to those subordinated features (size, degree of development, surface features, etc.), implying that the number of the smallest parts is not significantly different in both antimers.
E q u a l a n t i m e r s are either symmetric or congruent.
Two equal antimers are s y m m e t r i c when the position of the smallest parts are, to be sure, r e l a t i v e l y t h e s a m e in both, but absolutely opposite, implying that both counterparts (antimers) are mirror images of each other, like left and right. They never can be mapped onto themselves by any mechanical operation like for instance rotation or translation.
Two equal antimers are c o n g r u e n t on the other hand, when the position of the smallest parts is not only relatively the same in both, but also t h e s a m e i n a n a b s o l u t e w a y, implying that the two counterparts can be mapped onto each other completely by any mechanical operation, and that they can represent each other completely. Congruent are for instance the four antimers of a medusa, the five radial segments of the so-called regular five-fold flowers. Symmetric are the two body halves in Vertebrates and in Articulate animals, further, each two of the four paired antimers in symmetric Echinoderms (Clypeaster, etc.), each two of the four paired antimers of the so-called irregular five-fold flowers.
Strictly speaking one can detect an analogous difference, as it occurs between congruent and symmetrically e q u a l antimers, also in the case of s i m i l a r antimers, and we can accordingly distinguish between positively similar and negatively similar antimers.
As p o s i t i v e l y s i m i l a r can be denoted those similar antimers in which those organs and parts of organs that are in the same way present in both antimers, also show the same relative and absolute connection, the same positioning with respect to the median plane or median line of the body.
As n e g a t i v e l y s i m i l a r (or symmetrically similar) on the other hand, can be denoted those similar antimers, in which also this positioning is absolutely opposite.
Examples are provided by the symmetrical sea-urchins (Clypeaster, etc.). In these Echinoderms the five antimers (radii), which are congruent in other cases, are not congruent, but pair-wise symmetrical, implying that one can distinguish two pairs of radii and one unpaired radius.
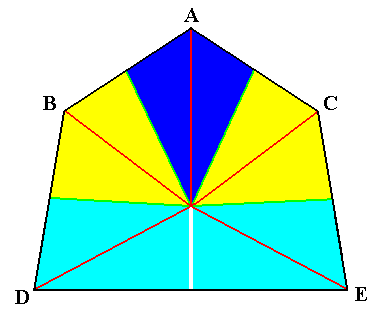
Figure 8. Diagram illustrating the symmetry relations of bilateral sea-urchins. The five antimers are indicated by colors and letters. Darkblue ( A ) : unpaired dorsal antimer. Yellow ( B and C ) : paired dorsal antimers. Light blue ( D and E ) : paired ventral antimers. Radial cross axes are indicated as red lines. The main axis of the body is perpendicular to the plane of the drawing and goes through the point of intersection of the cross axes. A+B+C is called the Trivium, D+E the Bivium.
The whole body can be divided into a dorsal and ventral part. The dorsal (normally anterior) part is called Trivium, because it contains the unpaired radius (antimer) and the dorsal (anterior) pair of paired radii (antimers), while the ventral part is called Bivium, it contains the ventral (posterior) pair of paired radii (antimers). When we indicate the unpaired radius as A, both radii, adjacent to it as B and C, and both ventral radii of the Bivium as D and E (See the above Figure), implying that the right half of the body consists of A/2 + C + E, the left half of A/2 + B + D, then B and C are symmetrically equal to each other. Also D and E are symmetrically equal to each other. B and D on the other hand, are positively similar, B and E are negatively similar, C and E are positively similar, and C and D are negatively similar.
Total absence of a composition of antimers of the body is only found in very few organ complexes, namely in the absolutely regular and absolutely irregular complexes. There exists only one absolutely regular (geometric) body, namely the sphere, which is, in a geometrically pure fashion, materialized in certain Radiolaria. In this case we can set the antimers as being infinite in number. Reversely, this number is zero in many sponges, the wholly irregular or amorphous body of which does not show any division into similar organ complexes that one could assess as antimers, i.e. such a body does not show any regular repetition of macroscopical parts. The same applies to many cryptogamous plants.
For the characteristic habitus, as expressed by the terms "bilateral" and "radiate" types, the different ways in which the antimers get together in configuring the whole body, are of the utmost importance. In the genuine bilateral animals, the Dipleura, the body of which consists of only t w o antimers (the "symmetric body halves"), as in Vertebrates, Articulate animals and Molluscs, both counterparts position themselves ajacent to each other by two surfaces facing each other, effecting their going to have one p l a n e in common. This plane is the median plane of the body. In genuine radiate animals on the other hand, fully regular (Medusae -- jellyfishes -- and Asteroids -- starfishes) as well as bilateral-symmetric (Ctenophors and many sea-urchins, like Clypeaster), in which more than two antimers ("radial segments" or "radii") get together to form the whole body, those antimers meet in one l i n e, the body's main axis, and thus having one edge in common. More seldom, namely in organisms of which the basic form is a regular, i.e. an endospherical, polyhedron (a polyhedron such that all its corners lie on a sphere), the antimers meet in only one p o i n t and consequently having only this point in common.
Specific differences with respect to the configuration of the antimers of the person (fifth-order morphological individual, or form individual in a narrow sense) are, in higher plants, often shown by the asexual persons (leaf off-shoots, etc.) and sexual persons (flower off-shoots). The latter, as being morphologically higher developed and differentiated, in most cases show us the same regular and easy recognizable composition out of antimers, as in most animal persons. So we see a clear correspondence between the regular Echinoderms (starfishes, etc.) and the regular five-fold flowers.
Haeckel (1866) considered the (adult) Echinoderm individuals as persons or fifth-order form individuals, on the basis of his theory of the phylogenetic origin of the Echinoderms from certain articulated worms. This theory seems to be abandoned by later investigators, judging to its total absence (as far as I could see) in the modern literature (The Echinoderms are a very unique and aberrant animal group with a disputed status). So also here, on this website, that theory is abandoned. This has its consequences with respect to the tectological assessment of adult Echinoderms, resulting in the conclusion that individual adult Echinoderms are not persons, i.e. are not fifth-order form individuals, but single metamers, i.e. fourth-order form individuals. For the present discussion, however, concerning the composition out of (five) antimers and the comparison with five-fold flowers this is of minor importance. We will discuss the metamer status of Echinoderms in the section devoted to metamers.
The "irregular" Echinoderms correspond with respect to their composition out of antimers to the irregular five-fold flowers. Also the diversity in the mode of this composition, which determines the characteristic physiognomy of flowers, is not less as in the corresponding animals. In the case of leaf off-shoots on the other hand -- the asexual individuals of the higher plants -- these composition relationships, which are expressed in the positioning of the leafs (phyllotaxis), are generally seldom as simple, regular and clear, as in flowers. Very often the simple condition where the positioning of the leaves is such as to allow the easy assessment of antimers of the off-shoot is replaced by the fact that the leaves are s p i r a l l y ordered along the stem of the off-shoot.
We present the antimers or counterparts as t h i r d-order morphological individuals, because the genuine proper antimers in all cases represent organ complexes, i.e. units, consisting of a multiplicity of second-order individuals.
Perhaps it would be, in more than one respect, more appropriate to interchange the ranking of both individualities, and consider the a n t i m e r s as s e c o n d-order, and the o r g a n s as t h i r d-order morphological individuals. In favor of such an assessment could be the fact that also in many organisms, which do not (yet) possess distinct organs, their body (a single cell) appears nevertheless to be composed of antimers, like in Diatomeans and many other Protists. Further we must consider the fact that also each single organ appears to be composed of several antimers, for example a single leaf of two antimers, a hand-like leaf (i.e. a leaf that is split into several more or less equal pieces like the fingers of a hand) of three, four, five antimers, etc. Meanwhile such an assesment would, strictly speaking, lead to the consideration of an antimer as a f i r s t-order form individual, because most unicellular organisms seem to be already composed of several antimers, implying that a single cell would represent a unity which consists of a multiplicity of antimers. Here, as also in many other cases, a strict ranking of morphological individualities appears impossible to implement, the more so when they, as so often occurs, intermingle with each other in complex ways. We can, however, nevertheless unambiguously determine this ranking also in the present case, when we investigate the subordination of the several categories in all organisms, and then derive the general law determining this ranking. And on this empirical basis the ranking here presented seems to be the right one. The problem is solved as soon as we distinguish between a n t i m e r s and p a r a m e r s (counterparts and next-to parts). And the very fact of the possibility to d e f i n e these antimers and paramers indicates the correctness of our relevant considerations. We did this at the end of the section on organs (Also with respect to epimers, that will again be treated further down, namely in the section on metamers).
In many cases we see that subordinated parts, for instance single organs, repeat the whole form as well as the characteristic composition of the whole organism. So in Arthropods the articulation of the trunk is repeated in that of the extremities (legs, etc.), and in Mimose plants the articulation of the stem (of any leaf off-shoot) is repeated in the feathering of their leaves. In the same way (and this is especially relevant here) we see that the homotypic composition (i.e. the composition out of antimers) of the whole organism is often repeated by an analogous composition in certain parts or organs. The latter often appear to be composed of homotypic parts in such a regular and constant way, as is the case in the whole person, for example the simple leaves in higher plants (and these leaves are organs!), which are constantly composed of two symmetric halves. In order to exclude any confusion between these subordinated counterparts, and antimers of the whole organism, we will denote the first generally as homonomous parts, n e x t - t o p a r t s o r p a r a m e r s. Such are, for example, the two halves of the dipleural leaves, the three leaflets of three-fold leaves (for instance of a clover leaf), the five toes of the human foot and of the vertebrate foot generally.
P a r a m e r s or next-to parts (homonomous parts) thus are generally corresponding parts, which lie around a cross axis or lateral axis of the body (or around the main axis of a single body part), next to each other.
A n t i m e r s or counterparts on the other hand, are corresponding parts, which lie around the main axis of the body next to each other.
Paramers are always subordinated parts of a first- or second-order form individual, antimers on the other hand, of a fourth- or fifth-order form individual.
The paramers accordingly relate to the antimers analogously as do the epimers (that will be treated of below) to the metamers.
The next Figure depicts a leaf (i.e. an organ) consisting of five paramers.

Figure 9. A five-fold ivy leaf. The five lobes of this organ are clearly expressed by their respective main veins. This, and their configuration, justifies to interpret them as paramers. The leaf's main axis cannot be chosen at will because it is (already) determined in virtue of the configuration of the five paramers : These, because they are paramers, lie around some axis, and this axis must be the main axis of the leaf. This main axis should then accordingly be the one that is perpendicular to the surface of the leaf. And indeed the five paramers lie around this axis. The axis that is perpendicular to the leaf's (only) plane of symmetry (mirror plane), is necessarily the lateral axis of the leaf. The axis that runs along the leaf's main vein must then be the dorso-ventral axis.
In the figure we can see that the leaf's dorso-ventral axis coincides with a cross axis of the off-shoot to which the leaf is attached (when the leaf is stretched out such that the main vein is the straight extension of the leaf stalk). The paramers group themselves around the leaf's main axis which is perpendicular to the leaf's surface and thus parallel to the main axis of the off-shoot. So in determining whether some corresponding form units are antimers or paramers, the identity (orientation) of the axis, around which these form units are placed, i.e. whether or not this axis is an extension of a cross axis of the (whole body), is only decisive when indeed that axis is definitely not the main axis of the whole body, which can be seen by its different orientation. In that case the form units are paramers. In all other cases (when the orientation of the axis, around which the form units lie, is such as it to run parallel to the main axis of the whole body) we can still recognize these form units as paramers (as opposed to antimers) by the fact that they are form units subordinated to an o r g a n (in the morphological sense, i.e. a second-order form individual). An antimer on the other hand, is a form unit that is directly subordinated to a metamer or to a person, i.e. to a fourth-order form individual or to a fifth-order form individual.
In Promorphology or the doctrine of the stereometric basic forms of organisms, which will be dealt with after finishing Tectology, we will not distinguish between antimers and paramers, because in Promorphology this distiction is not important.
Further we must realize that the c e l l is the u n i t of Tectology, implying (as was already related to above) that in a tectological sense we should not consider paramers in cells. In Promorphology we do so but call them antimers. When we nevertheless distinguish within Tectology paramers (and epimers) in single cells, we should see them as an analogical phenomenon with respect to the genuine tectological composition of organisms out of supra-cellular units.
In the next document we will deal with the next order of organic individuality, the metamers or fourth-order morphological individuals.
To continue click HERE to study the next morphological individuality : metamers.
back to homepage
back to retrospect and continuation page
back to Internal Structure of 3-D Crystals
back to The Shapes of 3-D Crystals
back to The Thermodynamics of Crystals
back to Introduction to Tectology