
e-mail :  ( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
Before we start, in the next four documents, to deal with the palaeozoic Blattaria fauna of, respectively, the Saar Basin, the Wettiner Basin, the Dunkard Basin, and the Kuznets Basin, we will, first of all, i.e. in the present document, describe the palaeo-geographical, the palaeo-ecological and the palaeo-climatological situation and context of those basins as they existed on the supercontinent Pangea in late palaeozoic times. That is, we will describe the overall context of these basins, which context is the Pangea Supercontinent. We will follow the transformations of local ecologies on the northern half of the continent from lower upper-Carboniferous up to upper Permian times. And when this is done (in the present document), we have gathered enough knowledge of the overall biological context of the Blattarian faunae of the four basins (Saar-, Wettiner-, Dunkard-, and Kuznets-basin), and are then in a position to precisely assess the nature of the morphological, geographical, and evolutionary relationship existing between these four faunae. And this nature may in turn become evidence of the polytopous, and therefore polyphyletic, origin of at least the blattarian family Archimylacridae, which finding may, after having found other cases of polyphyly of alleged monophyletic groups, be generalized, not only over all insects but over all organisms, meaning that all organisms have originated polyphyletically, i.e. independently of each other.
Environmental and paleogeographical conditions of the coal basins in the Saar and Wettin regions, and of the Dunkard and Kuznets coal basins as they are situated on the northern half of the Pangea Supercontinent.
The mentioned coal basins existed, or were formed, in upper Carboniferous and lower Permian times. Although the continental mass was at the the time not completely continuous, we may nevertheless speak of a single huge continent, the Laurasian-Gondwana continent : Pangea :
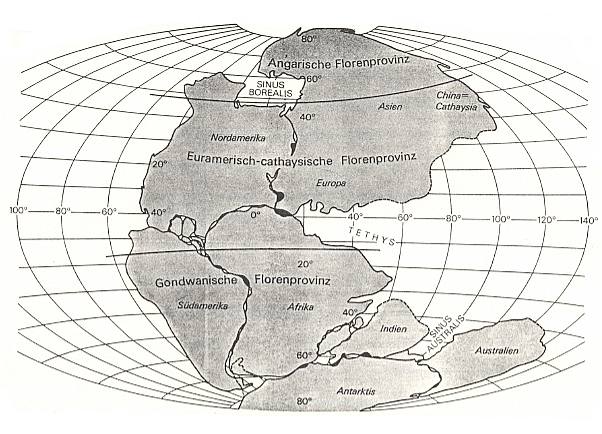
Figure 1 : Pangea. Schematic reproduction of the floristic distribution at the time of the later Westphalian (upper Carboniferous) until in later Autun (lower Permian). Position of the continental masses after DIETZ and HOLDEN, 1970.
(From REMY and REMY, 1977).
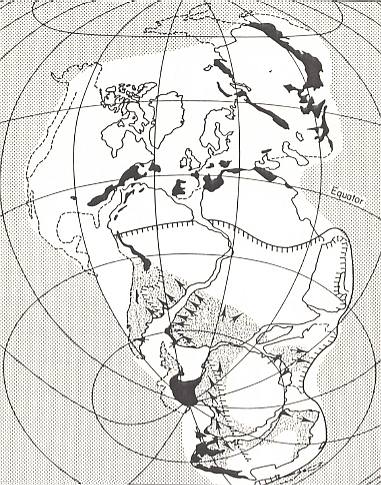
Figure 2 : Pangea [white area on map] 280 million years ago (lower Permian). Palaeolatitudes are based on palaeomagnetic evidence (MOREL and IRVING, 1981). The late Palaeozoic fold belts (e.g. Appalachians, Hercynian, Urals) are shown in black. Areas affected by glaciation after CROWELL and FRAKES (1970) and inhabited Glossopteris flora after PLUMSTEAD (1973). Edge of Precambrium Shield from POWELL et al. (1980).

Prevailing conditions on the Pangea surface.
Transgressions (oceanic expansions).
The closure of a number of oceans at the formation of Pangea brought with it large-scale transgressions, which, together with other factors (climatological and botanical), created the conditions for the formation of large costal swamps where coal could form (WINDLEY, p. 168, 1986) [At page 173 WINDLEY, however, states exactly the opposite : the formation of Pangea resulted in regression]. According to me transgressions are caused by the (horizontal) increase of area lying higher than the oceanic crust, by accretion of SIAL-crust during the Caledonic and Hercynic orogens (mountain formations), whereby, however, also stacking of layers took place.
Moreover, on the South-Continents (Southern half of Pangea) there was an ice-age going on, maybe of a periodic nature, causing the sea-level to rise during the interglacialia, and to fall during the glacialia, which on the North-Continents caused corresponding transgressions and regressions. Whether such a correlation was at the time really the case must, however, be further investigated.
So these transgressions will have plate-tectonic as well as climatological causes, and these probably will be linked in one way or another.
Elevation.
Based on calculations of sediment thickness it is assumed that the average elevation (height above palaeo-sealevel) of Pangea (in the early Mesozoic) was about 1300 m, that is, almost twice as high as it is today.
So the supercontinent had extensive plateaus, tablelands, and an internal drainage system as we see it about in central Asia today (WINDLEY, 1986).
Orientation.
During the late Palaeozoic the Gondwana-Laurasia Continent (Pangea) was stretched out in meridional direction, from the South Pole up to high northern latitudes (see Figures 1 and 2, above ) and as a result the oceanic and atmospheric circulations displayed meridional patterns around the world (WINDLEY, 1986). And this, of course, influences the distribution of the macroclimate on Pangea.
Drift.
During the period from Mid-Permian to Mid-Triassic this whole Pangea Continent drifted about 300 north (WINDLEY, 1986), and after this it slowly but definitely began to disintegrate.
Prevailing macroclimatic conditions on Pangea.
The climate of a land-mass depends on the special physical local circumstances (vicinity of oceanic currents, elevation, etc.), but also depends on the geographic position of the area with respect to the equator. Fauna and flora of a given region directly reflect that region's climate, but also a number of sediment types do. While palaeomagnetic data are evidence of the relative positions of the continents, the position of the continents with respect to the palaeo-equator can also be determined on the basis of climate-sensitive sediments, such as carbonates, evaporites, and redbeds (all this in warm and/or dry climates), or tillits (these in glacial climates). These sediments directly reflect the climate, and indirectly the geographical position at the time of the regions in question with respect to the palaeo-equator.
Because the climatic pattern in the time of Pangea surely has been different from that of today, this is expected to be expressed in discrepancies between, on the one hand, the determination of location and orientation of given regions based on sediments, and the determination based on palaeo-magnetism, on the other.
Today's carbonates, especially reefs, form in warm water (above 200C). Often associated with this are evaporites, which may form in a part of sea enclosed by a reef. Today this takes place in dry climates, (however) not close to the equator. Redbeds today are exclusively formed at latitudes of less than 300 from the equator, and all known redbed-florae point to a high temperature or a tropical climate (WINDLEY, 1986). [Redbeds are arkoses, sandstones, shales and conglomerates which contain haematite as a visible pigmenting agent.]
A warm and humid climate, the presence of shallow water and an appropriate vegetation, are conditions for the formation of coal, (i.e. another sediment indicating climatic conditions). The coal-deposits of the Carboniferous were formed in a tropical climate, while those of the Permian were formed from gymnospermian forests (such as pine-trees) in a cool climate.
Tillites (yet another such sediment) are fossil glacial deposits which -- how could it be otherwise -- point to glaciation of the region.
As regards the Permian of the North Continents it is determined that evaporites [such as gypsum and salt] are generally present between 00 and 350 palaeo-latitude, carbonates [such as limestone] between 00 and 500 palaeo-latitude, and redbeds between 00 and 300 palaeolatitude, and that coal-beds (late Permian) are generally present between 300 and 700 palaeo-latitude.
The coal-beds of the Carboniferous of the North Continents were generally present between 00 and 700 palaeo-latitude (WINDLEY, 1986).
From the early Carboniferous up to the middle of the Permian on the Southern-Hemisphere the climate became in such a degree colder that extensive permanent ice-caps were formed on the Gondwana Continent, with the resulting glacial deposits (tillites), see Figure 6.
Whether these distribution patterns are sufficiently sharp to serve as a basis of palaeo-climatic reconstructions I do not know. Anyway, it is visible that the coal-beds were, as to palaeo-latitude, situated more or less above the carbonate-evaporite-redbeds belt, except the Carboniferous and the Tertiary coalbeds.
In the next Figure I present a map of the possible macroclimatologic distribution on the (later) Pangea continent, in which figure, however, no temperatures are given. Northern and southern latitudes had a subhumid and humid climate, while in the high-pressure regions (i.e. those latitudes where we now have these high pressures) there was a dry climate, and in the tropical belt a wet climate.
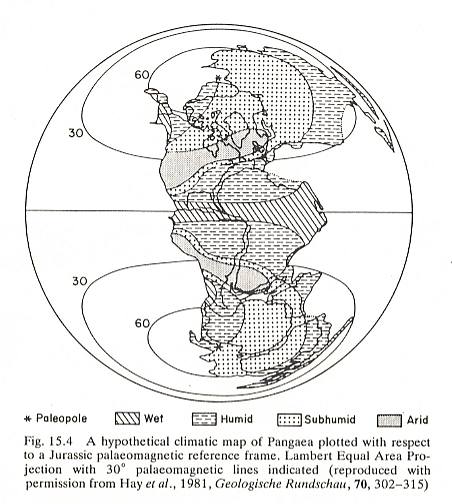
The glaciation of the South Continents (Figure 2 ) was concluded already before the Triassic : The beginning of the separation of Laurasia from Gondwana and the formation of the North Atlantic Ocean made it possible that ocean-atmospheric circulation changed from a meridional pattern to a more latitudinal one (dominating all over the Mesozoic), preventing the growth and expansion of ice-caps (WINDLEY, 1986).
Biological Anagenesis on Pangea
Biological anagenesis is the supposed "higher-development" of organisms in the course of their geologic history, meaning that their morphological organization became more integrated. The supposed presence of this phenomenon in the evolution of organisms is, of course, based on the more general supposition that organisms, in the Explicate Order (i.e. the material and physical Order of Reality), have evolved from each other along genealogic lines of descent. Anagenesis is supposed to be documented by the morphology, physiology, and behavior of recent organisms and organismic types (i.e. when we compare these organisms and organismic types with one another), and (also documented by) the successive appearance in geological time of the various stages of anagenesis. In our noëtic theory of evolution, however, it is supposed that organizational plans of organisms were developed in the Implicate Order (i.e. the immaterial, non-physical Order of Reality) in the noëtic process of immaterial forms developing into strategies to exist in the Explicate Order. And these strategies then are projected into the Explicate Order when appropriate ecological niches are actually present in that Order to receive the materialized strategies. And so, in this theory, migration and isolation of organisms do not play a major role in evolution. But in current theory much of the evolution of organisms is seen as a result of geographical or ecological isolation of organisms of a given species. The various isolated populations of such a species then subsequently develop into different species. Also migrations (where possible) of organisms, leading them to different ecological conditions, are supposed to result in the origin of new species and of new ecological types. Along such lines a, for instance, great share of endemic species in a given region is explained by the lack of intensive immigration, this lack in turn being caused by that region having become geographically or ecologically isolated from its surroundings.
Turning, for a while, to the palaeozoic cockroaches, we can safely hold that they, as medium- to moderately large-sized insects chiefly living on the forest-floor among plant-debris, and not being powerful flyers, cannot perform long-term migrations (during the course of many generations) over large distances on the surface of the Supercontinent Pangea, because, although it was not split up by major oceans, it was ecologically (and in connection with this, also qua relief) very diversified, as can be read off from the patterned macroclimate, see Figure above, but also generally from what can be expected from a large land-mass the vegetation of which, and therefore the faunistic complexes, depends on local elevation, local climate, local humidity, etc. Now, if we, in anticipation, consider (still not yet in detail) the cockroach tegmina found in palaeozoic deposits of western Europe (Saar and Wettiner Basins) and in those of the Kusnetz Basin in Siberia, we see these tegmina to be very similar in both regions, which are very far apart. Such a similarity between fossils from widely separated regions, fossils belonging indeed to the same family, Archimylacridae, should be explained. And normally it is explained by the supposed phenomenon of long-term and long-distance migration of members of this family. In other words such an explanation is based on current monophyletic theory : If the Archimylacridae form a true taxon, then, in current theory, by definition they have evolved from a single ancestral species, the stem-species of the family. But this also means that the origin of the family has taken place in one definite locality, namely there where one of the populations of some particular species became ecologically or geographically isolated, or had migrated into a different ecological zone, and became the stem-species of the new family. So, in current theory any given taxon is not only monophyletic but also monotopous as to its origin. Therefore, an observed presence of representatives of a given family in different regions far apart from each other must be explained by subsequent migration from the place of origin of the family to those different regions. But in the case of the palaeozoic cockroaches of Europe and Asia (Siberia) this could hardly have taken place, because of their relative immobility and their tight connection with warm and humid places. So the family Archimylacridae at least, cannot be monotopous, and therefore not be monophyletic. It must be polyphyletic. The family has originated in several (even far away) places independently. And although, of course, short-term migrations over short distances have often taken place, they cannot account for the very wide distribution of the family. Moreover, as we will see, long-term and long-distance migration on land of small animals (taking many generations), is, according to current theory, resulting in strong evolution, letting the original family fan out into many new families and types. We can see nothing of this in palaeozoic cockroaches. At least in the palaeozoic they have produced only a few closely related types. And except for the termites (which appear much later) we cannot point to non-blattarian insects that might have been evolved from cockroaches.
In what comes next we will work this out further, and in more details, but it is wise to have these preliminary findings in mind when reading about the biological anagenesis on the supercontinent Pangea in Carboniferous and Permian times.
Plate tectonics makes it possible for continents to break up or collide, resulting in great changes in faunistic and floristic provinces and in migration barriers.
There are indications that in terrestrial faunae and florae evolution goes faster, i.e. always leads to anagenetic steps ('higher-development'), if these faunae and florae find themselves on a large land-mass, because here we have to do with complex ecological relations and, as a result, a wide spectrum of adaptive zones.
The Supercontinent, at the end of the Carboniferous (and up to in the Triassic), satisfies in this way at least one main condition for such a large-scale anagenesis. With respect to the entomofaunae (the faunae of insects), which, as to the upper Carboniferous and especially as to the Permian, are relatively well documented by fossils, this anagenesis is, at least in a first qualitative overview, indeed evident : Many modern groups such as beetles, cicadas, and lacewings originated in the Permian as new against the ancient carboniferous background.
In this kind of considerations we should, however, be certain of the reality of the taxa established by us, and know about the genealogic lines of descent as long as we, for the time being, adhere to current evolutionary theory. For example, beetles (Coleoptera), first appearing already in the Permian, apparently represent such a successful evolutionary accomplishment (judging from their wealth of species today) thanks to their possessing shields, elytra (protecting body and hindwings). These shields can be seen as newly appeared structures if we suppose that the Coleoptera have descended from neuropterous insects (such as alder flies) which possess homonomous membraneous wings. If, on the other hand, we assume that beetles have descended from cockroach-like insects, then these shields are not at all newly appeared structures, but having been present as stiffened forewings (tegmina) already abundantly since Westphalian (middle upper-Carboniferous) times. The success of beetles (elytra, tegmina) is in this case based on an event that had taken place long before the actual appearance of true beetles and perhaps also before the formation of the Supercontinent, - or that success is based on other structures of the holomorphology of the coleopterous structural plan. What we have said about beetles may hold for all other animal and plant groups as well.
WINDLEY, 1986, presents a diagram (see next Figure) depicting the correlation between faunal diversity and degree of continental fragmentation, in which we can see that this diversity is clearly less in those periods in which the continents were unified (end Precambrium and Permotriassic.
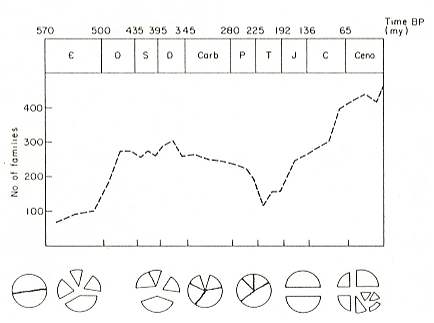
Figure 4 : A correlation of levels of faunal diversity with the patterns of continental assembly and fragmentation (indicated diagrammatically) throughout the Phanerozoic ( Cambrium, 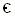 , - Ordovicium, O, - Silurian, S, - Devonian, D, - Carboniferous, Carb, - Permian, P, - Triassic, T, - Jurassic, J, - Cretaceous, C, - Cenozoic, Cen ).
, - Ordovicium, O, - Silurian, S, - Devonian, D, - Carboniferous, Carb, - Permian, P, - Triassic, T, - Jurassic, J, - Cretaceous, C, - Cenozoic, Cen ).
The diagram is concerned with benthonic shelf families from nine invertebrate phyla.
Vertical axis : Number of families. Horizontal axis : Time before present, in millions of years.
(After VALENTINE and MOORES, 1970, in WINDLEY, 1986).
This presentation seems to me very doubtful : Degree of diversity is here based on the number of families. At least in the context of phylogenetic systematics these taxa are not at all comparable, because, among other things, they are not based exclusively on age of origin, and because most 'families' do not at all represent historic units (phylogenetic taxa) but mere typological groupings, based on criteria that are different from group to group. Coleopterous families, for example, are surely not comparable (i.e. equivalent in relevant respect) with, say, families of birds, or with gymnospermian families for that matter. Indeed, in the diagram do figure families from nine invertebrate phyla.
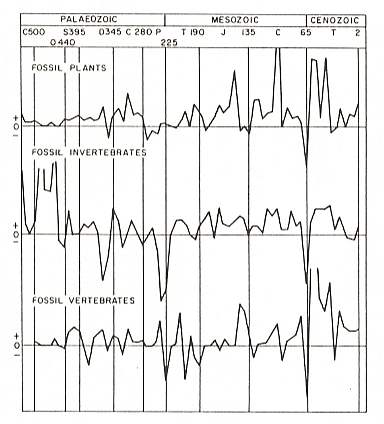
Figure 5 : Extinctions and appearances of new faunal and floral groups at stratigraphic boundaries during the Phanerozoic.
Vertical columns represent respectively Cambrium, Ordovicium, Silurian, Devonian, Carboniferous, Permian, Triassic, Jurassic, Cretaceous, Tertiary, and Quaternary.
(Modified (by Windley) after HOUSE, 1971, from I.G. GASS (Eds), Understanding the Earth. Figure in WINDLEY, 1986).
certainly do not hold for insects. Here the (upper Permian) fauna is diverse and with a relatively modern 'look' (so the upper Permian insect faunae of the Sojana river (Archangelsk region, Russia), Belmont/Warner's Bay (New South Wales, Australia), the Iljynsk and Jerunakowsk stages in the Kusnetz Basin (Siberia, Russia), but in fact also the ca. middle Permian Kusnetz stage of this basin).
Things concerning faunal extinctions and appearances have been considered here in the context of current evolutionary theory, in which groups of organisms are, or should, considered to be genuine historical entities, subjected to biotic and abiotic events on the surface of the Earth. According to this current theory, such a group of organisms has originated at a definite point in time and at a definite location on Earth, i.e. that point in time and that location where its stem-species had split off from its sister-species. Then, as a result of a great many subsequent bifurcations, migrations, and isolations, the group unfolds itself. It now consists of a multitude of mutually related species and inhabits a particular (type of) ecological zone, i.e. it has distributed itself over all the individual instances of this zone wherever they happen to exist on the Earth's surface. And on the fringes of this zone some members of the group may have successfully invaded other ecological zones nearby (ecologically and spatially), having adapted to them and so typologically having been split off from the group's main body, while phylogenetically still belonging to it.
In our noëtic theory of evolution existing or developing ecological zones are significant too with respect to the successive appearances and extinctions of organisms. But in contrast to the tenets of current theory, organisms have not evolved from other organisms, but appear as materialized strategies in the appropriate vacant ecological niches (being parts of some ecological zone). If we were there to actually observe these appearances and extinctions we would see nothing of the process of projection of a strategy into the Explicate Order, but would see the gradual replacement of one species by another, which we then explain by holding that the original species locally transforms into the other. Events like migration, isolation, random genetic mutation, natural selection, etc. surely, i.e. also according to our noëtic theory, do take place in the world of physical organisms, and do cause certain (local) changes, but cannot be considered to represent the very driving force of evolution. So in our view the taxa (in that view typologically conceived) are not intrinsically historic entities, although they are for certain definite entities. The historical aspect of them is more or less accidentally being attached to them as a result of the fact that their appearance in the Explicate Order is fully dependent on historical events in that Order. If such an event results in the creation of a vacant and new (type of) ecological niche, this niche will receive, from the Implicate Order, the projection of the corresponding strategy. And the new species will then appear in every existing individual instance of this (type of) niche. And when such a niche is later annihilated by other events its inhabitants will go extinct, either completely (as a species), namely when all individual instances of that niche are destroyed (i.e. having become unfit to support the species), or not completely so, when only a part of these instances is so destroyed.
Historical development of the surface of the northern half of the Pangea Supercontinent as a result of the conquest of the land by plants, followed by that of animals.
( From BEURLEN, 1978 )
Long before the formation of the Variscan (Hercynic) geosyncline in later palaeozoic times, and its uplifting resulting in the Variscan mountains, there was, in early palaeozoic times, the Caledonic geosyncline, the uplifting of which (in the upper Silurian) resulted in the Caledonic mountains. In the, as a result of the Caledonic folding, uprising land, flat anticlines formed heights, between which were wide, sinking synclinal zones. This large-scale undulation followed the directions of Caledonic folding, and thus was essentially SW-NE directed. In the zones of depression debris from the erosion of the heights was accumulated. So today we see the presence of zones of mighty deposits directed SW-NE, partly of terrestrial, partly fresh water origin, lying above the caledonically folded areas. Chiefly these sediments are sandstones. During weathering and pulverization the iron-compounds contained in the rocks are oxidized. Therefore, the weathered material, transported into the depressions, was chiefly of a red color. In England, where they are very typically developed, such clastic sediments, discordantly overlying caledonically folded early palaeozoic rocks, are referred to as "Oldred-Sandstone", in contrast to the young, newred-sandstone of later periods. The Oldred formations, as having appeared after the caledonic orogensis, are of Devonian age. Such Oldredsandstone-sequences in the depressions of the Caledonic mountain range are well-developed and investigated in England. Here they are present in a southern edge-depression in the Caledonics of southern Wales and of southern Ireland, and also present in the inland depressions thereof in northern England (Midlands) as well as in northern Ireland, in Scotland and on the Orkney islands. These oldred sandstones were deposited into humid lowlands often covered by lakes, and in them one finds the oldest and first terrestrial florae and faunae, which, of course, were still [ecologically] connected with the vicinity of water, i.e. they could only grow nearby it. The intermediate heights [separating the depressions] surely were not yet inhabited by living beings.
As has been said, the accumulated material in the Oldred depressions chiefly was of a red color. On the other hand, the accumulated masses in the frontal and inland depressions in the Variscan mountain range (Variscan orogenesis) of the upper Carboniferous are generally lightly colored, partly gray as a result of intermixed more or less carbonized plant remains, but not red. From this fact often a desert climate was inferred at the time the Oldred was formed, beause also today deserts often are of a reddish color. But the difference may be caused simply by a changed biological situation.
In Oldred times the first plants that had moved from the water into the air, which, still being principally amphibiotic, were closely connected with humid depressions, and, with their being still without leaves (Psilophytes) or with the first small leaves, did not yet create a continuous plant-cover. Humus-[vegetable mould]-production was extremely low, and outside the humid depressions plants apparently were totally absent (in Oldred times). Pulverization and weathering was, accordingly, still exclusively determined by the atmosphere and the oxygen contained therein. And so we have to do with weathering-by-oxidation, transforming the iron into its red oxide form. Humus materials hardly played any role in the weathering process. The weathering products had to become red independently of the climate. Because it is not probable that the first land-plants were desert-plants, needing specific adaptations, and because the rich development of primitive fish in Oldred times bears witness to the presence of lakes, the red color of the Oldred is more likely the result of this situation in the organic world than of a desert climate.
The intensified photosynthesis outside the water determined a great development of the plant world already in the Devonian, so that already in upper Oldred the leaf-bearing scouring rushes (Figure 6, D ), the scale-trees, i.e. Sigillaria- and Lepidodendron-like forms, (Figure 6, A, B ), and the ferns were already existing in the form of primitive representatives. As a result of surface-increaese of the leaves, photosynthesis grew and with it the formation of organic materials and plant development and diversity. So we see that from the end of the Devonian the leaf-bearing and spore-bearing plants exploded into development in the Carboniferous, not only with respect to the vast number of evolving forms, but also with respect to the development of (large) size. Scale-trees, scouring rushes, and fern-trees of 20 meter height and more, with corresponding thick trunks, grew up. Furthermore there grew Pteridosperms, having seeds like primitive gymnosperms and therefore belonging to them, and so also the Cordaite trees.
This rich flora formed, at least in the depressions, a continuous plant-cover of forests and perhaps already penetrated the somewhat more elevated areas adjacent to these depressions. It produced huge quantities of organic materials which in turn produced humus-rich peat-swamps in the depressions. Outside the peat-swamps weathering of dead plants parts used oxygen.
The acid humus-materials mobilized the iron and therefore bleached the soil and the sediment having resulted from weathering. Weathering could not longer be pure oxidation-weathering, but had been transformed by humus-caused bleaching processes. The color change from Oldred to Carboniferous was therefore not so much caused by change from an arid desert-climate to a humid climate, but the effect of a great development of the plant world, from then on more and more conquering the land masses.
The rich development of the plant-world, is a phenomenon of the whole surface of the Earth. As in western and central Europe we know huge coal-basins in North America : At the western edge of the Appalachians, in this time going through their final orogenesis [mountain building] -- one here speaks, instead of variscan, of appalachian orogenesis -- extensive front-depressions were formed, analogous to the Variscan frontal depression, with similar layer-sequences, containing numerous coal-seams. Further we have the Donetz Basin in the Ukraine, and the huge coal-basins in Siberia and China. Smaller but nevertheless remarkable coal-basins of the same period we know from Australia, South Africa, India, and South America.
Not any period whatsoever in the Earth's history coming after the Carboniferous does show such a huge accumulation of plant-products. The still young land-flora had, in its conquest of the new habitats, making possible an intensified photosynthesis, luxuriantly developed. Perhaps favoring this process was the nature of the ancient atmosphere, still having the higher CO2 content and the lower oxygen content of earlier times. And maybe the volcanism of the Variscan orogenesis has added extra CO2 to the atmosphere. And plants first of all need CO2 to build up their organic substances. The great development of plants now led to huge food resources to be present for a terrestrial fauna and contributed to the oxygen content of the atmosphere.
The presence of free oxygen in the atmosphere is a relatively late accomplishment having resulted from the rich development of algae in the late Pre-cambrium, and in the beginning of the early Palaeozoic the oxygen content was still lower than it is today. This fascilitated [because oxygen is still a dangerous substance] the transformation of the organic matter-economy into respiration. Under the cover of the still low oxygen content of the atmosphere in the Devonian, the first animals went over from water to land. When the huge plant production of the Carboniferous established, by further increasing the content of free oxygen, the atmospheric condition as it is today, so is with it offered, not only from the perspective of food, but also from the perspective of respiration, the primitive land animals, having originated in the Devonian, and being already used to respiration of air, a basis, intensifying development, for living-conditions. Indeed, also the development of the terrestrial fauna (that is, not only the terrestrial flora) in the Carboniferous shows it to be as luxuriant as that of plants.
( BEURLEN, 1978 )
The Ecosystems of the Pangea Continent (Upper Carboniferous - Permian)
(Literature : WINDLEY, 1986, REMY and REMY, 1977, ROHDENDORF and RASNITSYN, 1980.)
It is now time to delve a little deeper into the pure, and a bit more small-scale, ecological relations, that one thinks to be able to find with respect to the Pangean mesoclimates, the Pangean vegetation types and the associated entomofaunae (insect faunae).
The entomofaunae do occupy, in the terrestrial biotopes, an extraordinary important position : Insects are adapted to almost every type of environment or situation and play a determining role in the formation of almost all types of landscapes, as a result, among other things, of the fact that they impose a strongly regulating effect upon the plant cover. With this plant cover they today have every conceivable relationship, also with the dead parts of it, which they, together with other organisms, transform and return to the cycle of matter. Also they are a significant food resource for vertebrate animals.
In the Pangean period the close relationship of insects with the plant-world gradually began to develop, in feed-back with the spectacular evolution of the plants themselves, culminating in the angiosperm forests dominating the landscape from later Cretaceous times omwards at tropical, subtropical, and moderate latitudes.
Formation and nature of the plant-cover depend on the character of geotopes, i.e. the physical conditions of the terrain, such as elevation, relief, etc. Together with the macroclimate they then determine the hydrological condition of the soil and dependent thereon the hydro-, hygro-, and mesophilous (= not very wet, but also not very dry) plant-communities.
In carboniferous and permian times conditions as we experience them today did not prevail :
Forests with dense foliage did not exist. The forests contained trees having, it is true, foliage crowns, but they did not produce so much shade as is produced in today's rain forests. So within the confines of the forest really less of a genuine meso-climate, in which inwardly directed radiation (insolation), evaporation, and soil formation were influenced in a specific way, did exist.
There must have been extensive high-mountain ranges without a, differentiated in various levels, plant cover.
The climate on the continent, during the Palaeophyticum (Devonian-Permian) was, as a result of insolation, wind dehydration, and day-night contrast, in a higher degree uneven than is usually thought ( REMY and REMY, 1977).
The terrestrial climate of the time was certainly nowhere to be compared with the tropical rain-forest climate. According to REMY and REMY this meant that in those times, in contrast to the forest-climatic conditions of the Present, there was faster drainage and/or faster and stronger evaporation of water taking place, that there was a thinner humus layer present, a more intensive erosion, and a dominance of erosion-sediments. Further there was a different kind of iron-cycle in the soil, and finally a more abundant formation of fanglomeratic sediments (conglomerates of alluvial fans).
REMY and REMY, 1977, indicate that conclusions about the climate of ancient continents cannot be inferred from data from marine sediments (alone), and also not exclusively from evidence of evaporites and redbeds, but also not exclusively from evidence based on the azonal vegetation (aquatic and swamp vegetations).
The hygrophytes formed calamite forests and calamite + lycophyte forests. The mesophytes (vegetation at places where the water-table level does not, as in swamps, come above ground) formed fern-pteridosperm-cordaite-conifer forests.
The swamps of the Palaeophyticum of central Europe were formed on the large peneplains [= half-plain, almost flat, extremely weakly undulating relief] of the Hercynic [= Variscan] mountain range, especially in the ante-basins extending many hundreds of kilometres, but also in the inland basins (such as the Saar Basin). A uniform drop (sinking), largely compensated for by peat formation, took place. When such a drop was very strong, fresh-water basins or local marine transgressions or delta formations with large fans of debris could form, dependent on the tectonic-geomorphological circumstances. Accompanied with this are always changes of relief, which influence the water-table and thus the vegetation cover.
According to evidence found in plants, and according to evidence (of the presence) of types of sediments distributed over large areas and pointing to high humidity, viz., boghead-sediments [sediments containing algae and fungi], kannel-sediments [sediments with channeling as a result of (a.o.t.) effluent rain water [JB] ], Gyttja [sediment containing plankton remains, formed in inland basins] with plants, and, finally, coal, -- one must assume the presence, in the low-lands and on the lower parts of the peneplains [half-plains] of the Palaeophyticum, of a sufficient and over the year more or less evenly distributed supply of moist, be it from the atmosphere or from the soil. So there a humid to semihumid macro-, resp. mesoclimate prevailed. [A mesoclimate is a more or less specific climate prevailing in a particular geomorphologic (such as a coast, a river valley) or ecologic (such as a forest) area.]
In mountainous regions and on the higher parts of the peneplains a semi-humid to semi-arid mesoclimate is to be expected ( REMY and REMY, 1977).
The distribution of these two (meso-) climatic ranges and the determination of the resulting macroclimate depend on the distribution and nature of the large-, resp. small-scaleness of the geomorphology, such that in the Devonian has prevailed a more semi-humid, in the Carboniferous with extensive wet low-lands, a more humid, and in the Permian once again a more semi-humid, macroclimate (REMY and REMY, 1977). Much of this kind of data stems from (among others) Europe, but in, for example, the permian coal-measures of Siberia and of Australia the situation was partly the same, but partly also different : (coal) swamps can develop when much moisture and much vegetation are present : this is realized in tropical regions, but, (coal)swamps can also develop at high latitudes, where water does not sink thanks to the permafrost (permanent frozen level in the soil), and this together with the slow rate of decay of plant remains.
So in the Permian in Angaraland (ancient Siberia) a significant cooler climate prevailed, cooler than, for instance, in Europe, but nevertheless coal basins have been formed (Basin of Kuznets, Tunguska Basin).
In late carboniferous and permian times the continents were, as has been said, unified to form one single mass, which was partitioned by narrow epicontinental seas, mountain ranges and river-systems. On this continental mass, towards the end of the Carboniferous, three floristic provincies can be distinguished ( REMY and REMY, 1977) (see Figure 5, above ) : In the North there was the Angara Floristic Province. In the South there was the Gondwana Floristic Province. And in between there was the Eurameric-cathaysic Floristic Province (Cathaysia = China).
The Kuznets Basin geographically lies about on the boundary between the Eurameric-Cathaysic and the Angara Province. And we may take that the Tunguska Coal Basin lies about in the Angara Province (geograhically), whereas the Carboniferous and Perm of Europe and North America lie within the Eurameric-Cathaysic Province. The Australian coal-measures of course belong to the Gondwana Province with the Glossopteris-flora.
Having generally considered the Pangean land-mass as to its climate and flora (sometimes, more or less implicitly, having been led by example-regions), it is now time to move over to the consecutive Pangea periods that are relevant to the interpretation of the Blattarian faunae of the four above mentioned basins existing on Pangea in the upper Carboniferous and lower Permian, viz. the Saar Basin (present document), the Wettiner Basin (next document), the Dunkard Basin (next next document), and the Kuznets Basin (next next next document). So, now specifically of these periods ( Namurian [lower Upper-Carboniferous], Westphalian [middle Upper-Carboniferous], Stephanian [upper Upper-Carboniferous], and Rothliegendes [lower Permian] ) we will say something about the vegetation and insects in order to lay out the ecological context of the Blattaria (cockroaches) having lived then.

Pennsylvanian [upper Carboniferous) landscape in the swampy lowlands showing charateristic vegetation and sprawling amphibians. Part of a great mural by Rudolph F. Zallinger.
Plants : 1 - a treefern. 2 - Lepidodendron, a scale tree. 3 - Sigillaria, a scale tree. 4 - Cordaites, a precursor of the conifers. Calamites, a scouring rush. Animals : Diplovertebron, a primitive amphibian. 7 - Eryops, a large amphibian. 8 - Eogyrinus, one of the largest of amphibians. 9 - Seymouria, a primitive reptile. 10 - Limnoscelis, a reptile. 11 - Varanosaurus, a reptile. 12 - Meganeuron, a giant dragonflylike insect. (Yale Peabody Museum).
(In DUNBAR, 1963).
The Climate of the upper Carboniferous [= Pennsylvanian] land mass. [from DUNBAR, Historical Geology, 1963, p. 232-233]
Toward the end of the Carboniferous all chief plant groups (except the true flower-plants [angiosperms] appearing not until the Cretaceous) were represented, and with it all organizational types. All biotopes, up to the higher mountains, are populated by plants. New elements (with respect to the Devonian) are the Cordaites and the Conifers : Gymnosperms were already dominant in the Carboniferous. They inhabit the mesophilous regions (i.e. the non-swampy regions). In the swampy regions, on the other hand, the spore-plants dominated, especially the Lepidodendraceae and the Sigillariaceae and also the delicate (thin) leafy Filicatae, forming large swamp forests. And in addition also the Pteridospermatae and the Cordaitales were dominant. The plants most abundantly found belong to the hydro- and hygrophilous associations, or to the immediately following more mesophilous associations.
The Forests of the Swamp Lands. [ from DUNBAR, Historical Geology, 1963, p. 233-235]
The next three plates depict some more examples of carboniferous plants :
Plate 7a : Carboniferous plants.
(After FRAAS, 1910)

Plate 7b : Carboniferous plants.
(After FRAAS, 1910)
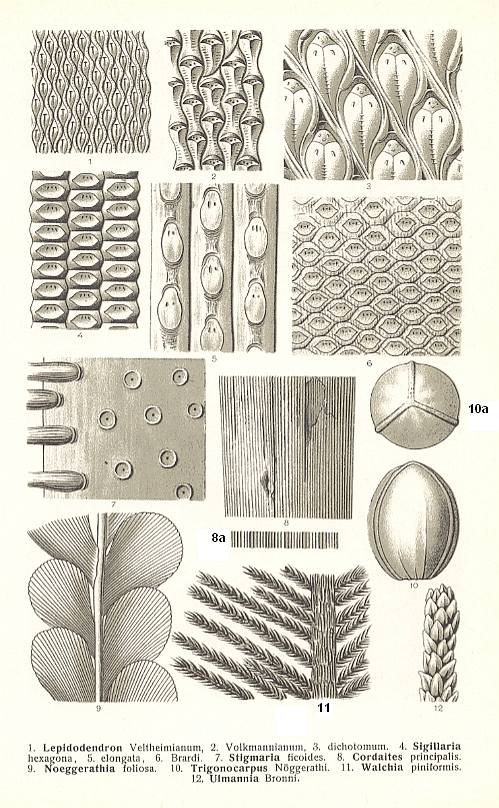
Plate 7c : Carboniferous plants.
(After FRAAS, 1910)
Animal Conquest of the Lands [ from DUNBAR, Historical Geology, 1963, p. 235-240]
Vertebrate animals are represented by abundant skeletal remains of Amphibia. From the Coal Measures of North America alone no fewer than 7 orders, 19 families, 46 genera, and 88 species are known. Such great diversity suggests that these animals were common in spite of the fact that they are still the rarest of fossils. The most remarkable single locality for such remains is at Linton, Ohio, at the base of the Freeport coal, where abundant ganoid fishes and no fewer than 50 species of Amphibia have been found.
The Pennsylvanian Amphibia were labyrinthodonts, and nearly all were small. Many of them were only a few inches long, and large ones, scarcely 10 feet over-all, would not be bigger than a large Florida alligator. The greatest of all is known only from its tracks, deeply impressed in Mid-Pennsylvanian sandstone near Lawrence, Kansas. This animal (Onychopus gigas) had blunt, stubby feet over 5 inches long, and although its stride was only about 30 inches, the right and left treats were wide apart, indicating a short but very heavy-bodied animal estimated to weigh not less than 500 or 600 pounds. There is some evidence that the feet were webbed like those of a crocodile.
A few reptiles of small size appeared during the later half of the period, but their remains are very rare.
( DUNBAR, 1963)
Additional data concering the physiography and origin of the upper Carboniferous swamps and subsequent coalfields.
In order to correctly evaluate the degree of ecological and physiographical isolation of the individual carboniferous forest-swamps from one another in a given overall swamp region, and of such an isolation of one overall region from another in space and time, and the effect such an isolation is supposed to have on the distribution of less mobile insects such as cockroaches living in such forests, (and from that drawing conclusions of monophyletic or polyphyletic origin and development), it is wise to add, to what has been already given above, some more data provided by other authors of the subject (but without pretending to be complete in this respect!). Here we will make use of two more sources :
F.J. NORTH, Insect-life in the coal forests, with special reference to South Wales [England], in Cardiff Naturalists' Society, 1929, Vol. LXII. Printed for the Society in 1931. We will quote and comment on a few (separated) parts of this article.
A.J. PANNEKOEK and L.M. van STRAATEN, Algemene Geologie, 1982. Also here we will quote and comment on a few parts concerning the origin and structure of coalfields.
From NORTH, 1929 (1931) (concentrating chiefly on the South Wales Coal Measures), pp. 26.
The conversion of vegetable tissues into coal only took place when the land, already practically at sea level, was subsiding at a slow rate that was just sufficient to maintain the surface at approximately the same level while the accumulation of dead plant-remains increased in thickness. On such a surface the products of forest growth would have been in a more or less waterlogged condition all the time, gradually decomposing in the absence of air, giving rise to jelly-like decomposition products that impregnated and embalmed the still undecomposed tissues. If, on the other hand, they are exposed to the air under relatively dry conditions, vegetable tissues tend to give rise almost entirely to gaseous decomposition products, and to disappear.
Since, then, the nature of coal and the mode of occurrence of coal seams indicate that the forests grew for the most part on or near swampy areas not much raised above sea level, and since, too, the Coal Measures occur over so large a part of North-western Europe, in tracts that owe their present shape and isolation to the results of folding, faulting, and denudation, there is a tendency to think of Coal Measure time as a period when dense swampy forests extended without a break over many thousands of square miles, giving rise to continuous layers of potential coal-substance. This is not likely to have been the case, for there must have been areas where the conditions were too dry for the conversion of plant remains into coal-substance, and others over which some of the forests failed to spread. See next Figure.
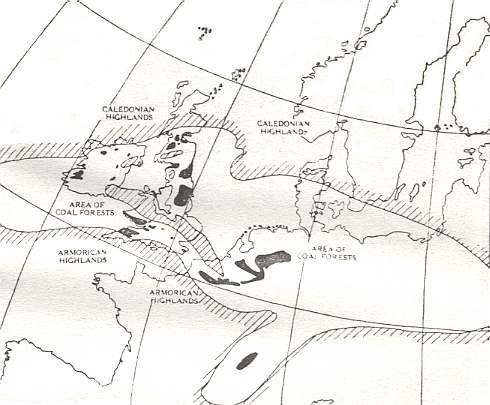
Figure 7aa : Map illustrating the former extent of the coal forests of Western Europe.
The Coal Measures were deposited over much of the large oval region occupying the centre of the map, but only in the areas coulored black have they escaped denudation and remain as coalfields. The probable limits of the swamp forest area are indicated by close parallel lines, and in a similarly marked area extending across Great Britain the Coal Measures never accumulated to the extent that they did elsewhere.
North of the western European region of coalfields there were the caledonian highlands, and south of the region of these coalfiels were the Armorican highlands.
(After G.B WALKER, in NORTH, 1931)
Earth movements which affected western and central Europe, at the close of the Coal Measures times, resulted in a general elevation of the land surface, and brought the coal-forming episode to a close by giving rise to climatic conditions that were unfavourable to the luxuriant growth of vegetation and to the conversion of its dead remains into coal.
Insects and the correlation of the Coal Measures.
The plants and shells of the coal measures have been studied with a view to their utilisation in the identification of successive levels or horizons in the Coal Measures, but when one remembers that plants and shells have been collected by the thousand -- one single paper on the flora of the Coal Measures of South Wales was based upon the determination of over 389000 fossil plants -- it will be realized that the known insect remains are too few for important stratigraphical conclusions to be based upon them. Nevertheless, what is already known is both suggestive and promising. All the insects found at very low horizons belong to the Palaeodictyoptera [= Dictyoneurida], which are the most primitive and generalized of the Carboniferous insects, while members of that group are not found above the lowest of the three divisions of the Coal Measures.
Of the Blattoidea [Blattida, Blattaria, cockroaches], the members of the family Archimylacridae (which probably represent a link between the certain members of the Palaeodictyoptera and the more highly specialised blattoids) have a fairly long time range, while the Mylacridae, which, of all the Carboniferous insects, approach most closely to the cockroaches [in a more restricted sense] and were derived from certain of the archimylacrids, did not appear until quite late in Coal Measures times and are confined to the uppermost division of that rock series. The bearing of this fact (primitive forms preceding those of higher specialisation) upon our interpretation of the development of the group is obvious, so also is the nature of the assistence that the insects will afford in determining the geological horizon of the beds in which they occur. The occurrence of members of the Mylacridae indicates that the beds belong to the higher divisions of the Coal Measures, while the presence of members of the Palaeodictyoptera alone indicates a low horizon. The evidence of the Archimylacridae is less definite, but if associated with palaeodictyopterids they indicate a lower horizon than if associated with mylacrids.
Habits and habitats of the Coal Measure insects.
One can well imagine that while the cockroaches spent most of their time among the rotting vegetation, although they could fly and were almost certainly able to swim, the flying insects (the Palaeodictyoptera) would have preferred the open spaces where lagoons or large pools broke the continuity of the forests. And to this extent we can picture the insect life of the coal forests : Many creatures lurking almost always in concealment, partly from choice, because in such surroundings they found an abundance of food, and partly from necessity, in order to escape the attentions of their more active and carnivorous contemporaries. Others flitting, like giant dragonflies, jerkily through the air, not always escaping the snapping jaws of the salamander-like amphibia that haunted the trunks of trees that had fallen near the water's edge.
The fossils throw light upon the climate and atmospheric conditions in the coal forests. It has been suggested that the luxuriant character of the vegetation indicates the existence of abnormal conditions -- that, for example, the air was much more heavily charged with carbon dioxide (CO2) than at the present time. But the evidence afforded by the insects supports the view that the conditions were not materially different from those still obtaining in a region of damp dense forest. The wing-veins indicate that the breathing arrangements were essentially like those of modern insects, which is not likely to have been the case in an atmosphere highly charged with carbon dioxide. Moreover, although a few giant dragonfly-like insects have been found, the majority were very similar in size to their nearest living representatives.
The little that is known of their life histories indicates that the Coal Measure insects did not have special pupal forms like those which, in certain modern insects, permit of a resting stage during the existence of unfavourable conditions. This suggests that there were, in the forests, no well-marked seasonal changes due to the periodic occurrence of very dry or very hot spells.
Larval forms have very rarely been found, and none have, as yet, been recorded from South Wales. In the youngest stages, at least, it is improbable that the chitinous covering had been developed to an extent that would cause the body to leave any impression, even in a fine-grained deposit like the mud that is now the Coal Measures shales, and reasons for the scarcity of larvae of more advanced growth will suggest itself in later paragraphs. The larval blattoids are naturally smaller than the adult, from which they do not differ very greatly in appearance except in the rudimentary condition of the wings, while there is still some doubt as to whether the larval forms of the Coal Measure dragonflies were aquatic or not. The breathing system of the larvae of modern dragonflies indicates, according to Tillyard, that originally they were not aquatic, and the opportunities in Coal Measure times for development in damp earth instead of in water were very great.
The feeding habits of the Coal Measure insects present interesting problems. In the case of the blattoids it is difficult to base any conclusions upon comparisons with the habits of their modern representatives, because nothing seems to come amiss to the cockroach of to-day, which has at its disposal a diet much more varied than that available to his Carboniferous relatives. As far as we know, animal life was not generally common in the Coal Measure forests, but there was rotting vegetation in abundance. A point of some significance in this connection is the not infrequent association of blattoid wings with the leaves known as Cordaites, and the occasional presence on those leaves of the coiled shells of the little aquatic worms Spirorbis, or of marks indicating the former attachments of such shells.
At the present time, Spirorbis lives on seaweed and is found on the leaves of trees like mangroves that occur in coastal swamps. Most palaeobotanists are agreed, however, that Cordaites lived in the dryer and more elevated regions of the coal forests, for the remains show no anatomical evidence of life in swampy surroundings, and there are no grounds for supposing that the leaves were awash during the life of the plant and so would have provided feeding grounds for aquatic worms. Owing, however, to their long thin strap-like nature, masses of such leaves would form excellent "rafts" if they fell into the water and drifted out into lagoons, and the association of blattoids with drifted cordaite remains, may indicate that the insects had accompanied the floating masses in order to feed upon the worms that had attached themselves to the leaves.
On the other hand, it may simply mean that the buoyancy of the wings was approximately equal to that of the leaves, and that the association is accidental.
We have still less to go upon in connection of the feeding habits of the flying insects, but such evidence as is available concerning their mouthparts, and comparison with their nearest living representatives suggests that many of them were carnivorous. They probably preyed upon the blattoids, and it has been suggested that the similarity between the blattoid wing and a pinnule of Neuropteris may have afforded some measure of protection to the insect, and if that is the case it is a very early instance of mimicry. See next Figure.
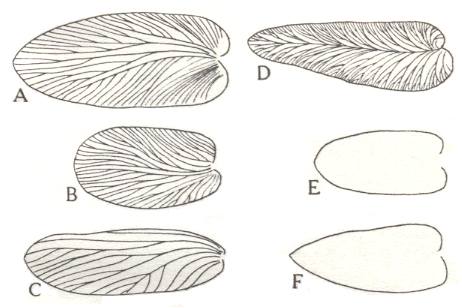
Figure 7aaa : Wings of insects compared with plant pinnules from the Coal Measures.
A-C are wings, which, in spite of their general similarity in outline to pinnules of various species of the plant Neuropteris (D-F), are readily distinguishable by the distribution of the veins, which, in the pinnules, are arranged more or less symmetrically about a conspicuous mid-rib. In A-C only the longitudinal veins and their branches are indicated. In many of these wings they are connected to each other by cross-veins or by an archedictyon (meshwork).
(After NORTH, 1931)
[Some of the flying insects were suctorial and lived on the juices of plants.]
In conclusion of this account by NORTH, 1931, of Coal Measure insects and their geological and biological context, we may add what NORTH had to say about a palaeo-entomologically very special and famous upper Carboniferous limnic coal-basin, Commenrty, France, where, together with other insects, lots of the ancient Palaeodictyoptera (Dictyoneurida) have been found :
The small coal basin of Commentry in central [NORTH erroneously writes Northern] France, only about 11 square miles in extent, has yielded the greatest wealth of insects remains yet found in one district [apart from the lower permian locality of Elmo in Kansas USA]. The assemblage is very varied. Many species are represented by parts not usually preserved, and te individuals are very numerous, the blattoids alone being responsible for over 1200 specimens. But, even here, although the conditions of sedimentation must have been favourable to their preservation, the shales have yielded no larval forms.
( NORTH, (1929) 1931 )
Some more data about the genesis and structure of Carboniferous coal basins
From A.J. PANNEKOEK and L.M. van STRAATEN, Algemene Geologie, 1982, pp.543.
Accumulations of vegetable material
Where dead vegetable material is in normal contact with air, the organic compounds are decomposed by bacteria, molds and fungi, whereby as an intermediate product humus is formed. Eventually, however, everything is being 'mineralized' where then chiefly H2O and CO2 (carbondioxide) are formed, and, in smaller amounts, among others, N2 (nitrogen) and NH3 (ammonia).
Where no free oxygen (O2) is present accumulations of vegetable remains are being transformed into peat. This happens where the pores in the accumulations are filled with stagnant water, as in swamps and at the bottom of lakes. The primary presence in the water of dissolved oxygen soon is being consumed during oxidation in the upper layers of the plant-residue. Below these layers further decomposition does take place, by action of anaerobic bacteria, but this goes much more slowly. Here especially CO2, H2O, and CH4 (methane) are released. And because in this biochemical transformation more oxygen and hydrogen disappears than carbon (C), the remaining material becomes enriched with carbon. In peat the content of carbon may rise from less than 50 to more than 60 percent of weight.
Conditions of peat formation
The most important conditions of peat formation are a humid climate where abundant plant-growth is possible, and a favorable condition of the terrain, in which the discharge of interstitial water comes to a halt, as in swamps and lake bottoms. One thus may find peat formation in the humid tropics and subtropics as well as in moderate humid climatic regions and even in subarctic regions. Only in the arid climatic belt and in frost climates peats almost completely are lacking. As to the condition of the terrain things are most favorable to accumulation in the extensive river plains at or just a little above sea-level. Between the rivers, accompanied by embankments, here lie extensive swamps or shallow lakes, into which river water only penetrates in periods of flooding (or) when the embankments break up. At the sea-side the swamps and lakes are often bordered by beach embankments, or, in the tropics, by a mangrove belt.
In peats of coastal planes in humid-subtropical and tropical climates we see forest-peat predominating (over peat with only low-growing vegetation). These subtropical and tropical forest-peats constitute the actualistic analogue of the peats out of which brown coal (lignite) and, subsequently, coal (pit coal) are formed. The flora from which the coals of the Carboniferous have been formed, was completely different from that of recent or late mesozoic peats, and chiefly consisted of extinct groups of tree-like spore-plants (spore-bearing plants), but the type of vegetation was comparable with the recent subtropical and tropical forest-peats.
The coal layer in relation to its direct petrologic context and environment
Most coal layers are bounded at the lower and upper side by characteristic sediment layers, resulting in the following (from top to bottom) structure :
Roof (plant bed),
Coal seam (sometimes interrupted by some thin sediment layers),
Seat rock, underclay.
The so-called seat rock (root floor) is the often clayish, fossil floor on which the trees and plants of the original swamp had settled. In the coal sequences of the Carboniferous this layer (seat rock) is often characterized by the presence of stigmaria ( Plate 7c, 7 ) : thick, horizontally running roots of swamp trees such as Sigillaria and Lepidodendron ( Figure 6, B and A ).
The coal seem is often covered by a clay (-stone) layer, which may contain, in addition to numerous washed-off plant remains, also many fossils of fresh-water clams, and then thus must have been deposited in a lake. This means that the swamp forest must have been 'drowned' and replaced by open water. In certain cases it was shallow sea-water that halted the peat formation.
Cyclothemes
A coal seam often constitutes a part of a regular sequence of sediment layers, which is called a cyclotheme. In regions with long-lasting subsidence there are a large number of cyclothemes, each with its own coal seam, on top of each other. See next Figures.
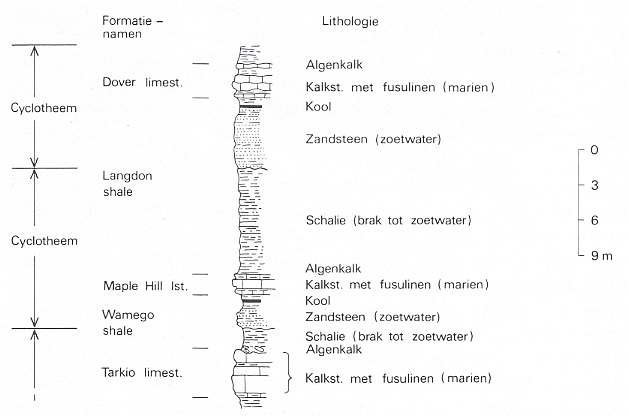
Figure 7bbb : Cyclothemes with an alternation of marine and fresh-water deposits. Upper Carboniferous, Kansas (USA). (After MOORE, 1950, in PANNEKOEK et al., 1982). "cyclotheem" = cyclotheme. "Formatie namen" = names of [geol.] formations. "limest." = limestone. "Lithologie" = lithology. "Algenkalk" = limestone with algae. "Kool" = coal. "Kalkst. met fusulinen (marien)" = Limestone with fusilines (marine). "Zandsteen (zoet water)" = Sandstone (fresh water). "Schalie (brak tot zoet water)" = shale (brackish to fresh water).
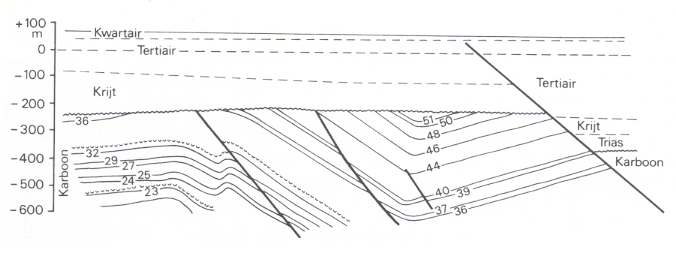
Figure 7ccc : Folded coal seams and some marine strata (together forming a series of cyclothemes) in the Carboniferous of southern Limburg (Netherlands), near the edge fault (to the right). Staatsmijn Maurits.
(After DIKKERS, 1945, in PANNEKOEK et al., 1982). "Kwartair" = Quaternairy. "Tertiair" = Tertiary. "Krijt" = Cretaceous. "Trias" = Triassic. "Karboon" = Carboniferous.
Solid curves - coal seams (some accompanied by marine strata). Thick solid lines - faults (geol. fractures).
The lacking numbers (of coal seams) here are not developed into true coal seams.
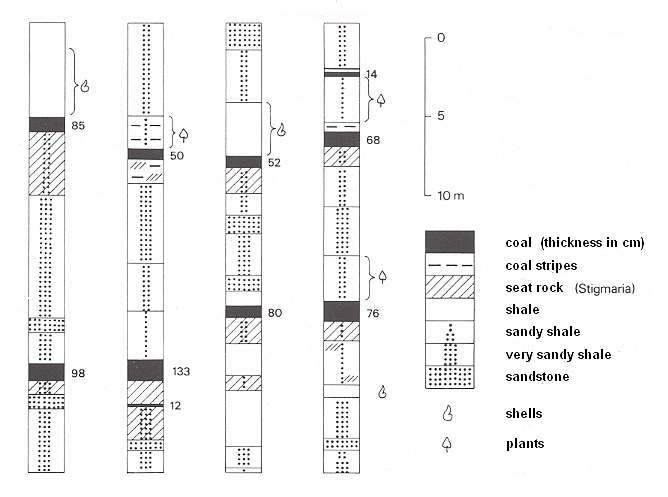
Figure 7ddd : Continental cyclothemes in the upper Carboniferous of Southern Limburg (Netherlands).
The four profiles [sections] are from different sub-levels of the upper Carboniferous, but show a corresponding structure : Below the coal seam always a seat rock, on top of the coal sometimes a lake-deposit (with shells), causing the coal [i.e. the original peat] to be drowned.
(After RUTTEN, 1947, in PANNEKOEK et al., 1982).
The thickness of the cyclothemes mostly varies between 5 and 15 meter, but may be larger or smaller.
Examples of cyclothemes with a marine intercalation we encounter in Illinois (USA), where the term 'cyclotheme' was first introduced, and in Kansas (USA) ( Figure 7bbb ). On top of the coal seam here lies a marine limestone, followed by marine shale and sandy shale. So here the peat-swamp was drowned as a result of a transgression of a shallow sea. Gradually more terrestrial material was deposited, resulting in the regression of the sea. On the now formed fluviatile coastal plain, were, between the river-courses, the lakes and swamps where forest-peats began to develop, later to be transformed into the next coal seam. With this, the cycle is concluded, and a new cycle begins.
If one goes from Kansas and Illinois eastward into the direction of the Appalachian mountains, then the marine layers become thinner until they completely disappear, and the total thickness of the Carboniferous and with it the number of cyclothemes becomes larger.
In the Carboniferous of western Europe we see a similar picture. Down into the upper Carboniferous, especially in England, most cyclothemes still have a marine intercalation, but in the large coal basin extending through northern France, Belgium, and southern Limburg (Netherlands) up to the Ruhr region, most cyclothemes are totally continental ( Figure 7ddd ) with only a few marine layers that can be followed over large distances. Some cyclothemes are even so incomplete that they chiefly consist of shale.
Looking for an explanation of such an always repeating regular sequence of sediments, it is obvious to think first of all of a periodically varying in intensity, or even jerkily, subsidence of the basin, expecting, in cases of strong subsiding, it to cause a drowning of the peat, while in a period of slow-down or cessation of subsiding, expecting the filling up of it again. This still does not explain the changing supply of sediment. For this several researchers assume that subsidence of the basin was compensated for by an also periodic uplift of the hinterland resulting in intensified erosion and sediment-supply. Other researchers have indicated that the changing sediment supply may have its cause in the sedimentation process itself, and exemplify this with the Mississippi delta. Between the river-courses are extensive forest-peats that eventually drown as a result of continuous subsidence of the floor and the strong compaction of the peat itself. And as soon as an embankment wall breaks through, the drowned region is quickly filled up with sediments from the new river-courses branching in all directions. That indeed such break-throughs did occur is evident from the sometimes encountered upright-standing pieces of trunk of carboniferous trees, that, before they could decay, were surrounded by fluviatile sand, and as such fossilized. The part of the delta left behind by the river at the break-through is not levelled up anymore by sediment and may in turn become a peat swamp and eventually drown.
[Of course all these events, each having been taken to form an explanation of the changing sediment supply (as it is visible in cyclothemes), have at one time or another, at one place or another, taken place. And it is certain that teh history of the coal basins is closely connected with the variscan orogenesis.]
And although this actualistic mechanism [of the breaking-through of embankment-walls, just described] may give an explanation of the details of the cyclothematic sedimentation, it fails to do so for large-scale marine transgressions. To explain this, we need to stress that periodic differences of subsidence and sediment supply, or fluctuations of the sea level, must have been involved.
In explaining the cyclothemes in the northern hemisphere, some authors have even pointed to glacial-eustatic fluctuations of sea-level [themselves resulting in transgressions] caused by the glaciations in the southern hemisphere at the time. Against this, one may point to the fact of cyclothemes changing in number from place to place [here many, there few] in the upper Carboniferous. [Transgressions in the northern hemisphere, caused by the melting of the southern hemisphere's ice during interglacialia, should indeed have left a bit more uniformity among cyclothemes of different places.]. Moreover, cyclothemes are also known from periods without ice-ages.
Coal basins
Formation of large forest-peats in coastal plains demands a subtle equilibrium to obtain between sedimentation and subsidence. This explains why the formation of coal seams -- or, first of all, peat formation -- in the geologic past was an exception rather than the rule. In most regions of subsidence the latter was ahead of sedimentation, resulting in marine conditions prevailing. Only in a subsiding region with strong, but changing, fluvial supply, now and then equilibrium could be established that was needed for the formation of extensive forest peats. In a number of subsiding regions these conditions must have been present repeatedly, as the large number of cyclothemes demonstrates, each cyclotheme with its own coal seam ( Figure 7ccc ). The thickness of the coal seams, however, is almost nowhere more than 2 meters.
Conditions, favorable to the formation of coal seams, have prevailed in Europe and North America during the Carboniferous, especially during upper Carboniferous times. In that period there were, along the just formed Hercynic [= Variscan] mountain-folding range, extensive, subsiding, frontal depresssions -- see next Figure -- filled up by rivers with rubble from the mountains [i.e. from the erosion of the mountains].
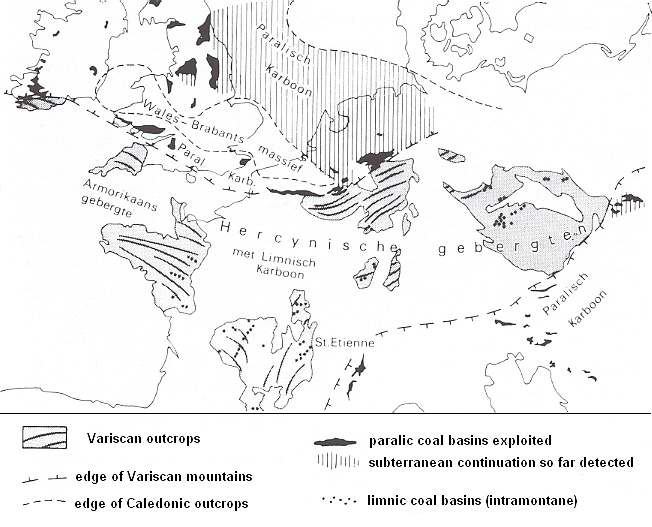
Figure 7eee : Paralic and limnic coal basins in the upper Carboniferous of western and central Europe. The paralic coal basins were in the coastal planes at the foot of the folding Variscal [= Hercynic] mountains. The limnic coal basins were to be found in intramontane depressions in the mountain range.
(After several sources, in PANNEKOEK et al., 1982). "Hercynische gebergten" = Variscan mountains. "Karboon" = Carboniferous. "Armorikaans gebergte" = Armorican Mountains. "limnische" = limnic.
See also Figure 7aa.
Because the sediment supply and the subsidence of the basins were kept about in equilibrium, we here find large numbers of coal seams ( Figure 7ccc ). In the upper Carboniferous of the Appalachians (USA) there were about 90 of them, an in the Ruhr region (western Germany) about 400 (inclusive the very thin layers, and even seat rocks without a true coal layer). The total thickness of the upper Carboniferous (as geologic formation) in the Ruhr region is, indeed, more than 4 kilometers. A belt of such coal basins extends, north of the Hercynic mountains, from England through northern France, Begium, southern Limburg (Netherlands) and the Ruhr region. The region of sedimentation gradually shifted to the north, while the southern edge of the basin was folded in a late phase of the Hercynic orogenesis. Nevertheless, even under the favorable circumstances of the upper Carboniferous, accumulation of plant material was the exception, not the rule.
Because the extensive coastal swamps lay virtually at sea-level, a somewhat increased subsidence or a small rise of sea-level could cause an extensive transgression, albeit of a very shallow sea. A facies, that is alternately continental and shallow-marine is called paralic. Close to the coast these transgressions were numerous, whereas more inland the coastal plain was only infrequently being flooded by the sea. But also then one speaks of paralic coal basins. In southern Limburg (Netherlands] one finds on average one thin marine layer in about 400 m sediment.
In contrast to paralic coal basins we have, in Europe, the so-called limnic coal basins, never reached by the sea. They were formed in swampy, intramonanous, subsiding basins within the Hercynic mountain range (see Figure 7eee ). These basins are formed only at a later stage in the Carboniferous, after the chief mountain chains had been folded. To such basins belong, for example, the basins of the Saar and of St. Etienne.
The formation of extensive forest peats, being preserved as coal seams, has taken place (when the terrestrial flora was, in this respect, sufficiently developed, i.e. since the Devonian) repeatedly, then in one region, then in another. Nevertheless this, as far as can be determined, has never been so intensive as it was in the Carboniferous.
( PANNEKOEK et al., 1982)
As to plants and insects of the Carboniferous, we can add the following :
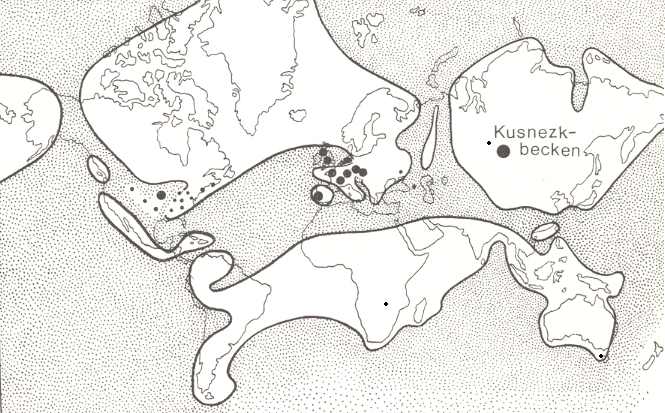
Figure 7 : Palaeogeography and insect localities in the Upper Carboniferous. From the palaeogeographic maps of TERMIER and TERMIER, SCHUCHERT, and BUBNOFF. The largest of the North American black dots : Mazon Creek (Illinois).
(After HENNIG, 1969. New localities added by PONT and SCHLEE, 1981).
We will now deal with the main divisions of the Upper Carboniferous separately.
The Cordaites ( Figure 6C ) become dominant. The outgoing Namurian and the older Westphalian are the periods of large coastal swamps covering immense areas. There were continuing formations of swamps interrupted by larger river-systems and in which larger and smaller lakes were embedded. Colossal forest swamps with Lycopsid forests, which, however, cast little shade, were the chief suppliers of the peat substance. So also were the Cordait forests, which certainly did cast more shade so that they might stimulate the development of epiphytes [living high on tree trunks in order to reach the light]. The Pteridospermatae and the Filicatae did cover, partly in the in-between swamp stages, partly in the zonal vegatation, large areas [Zonal vegetation is vegetation growing on dry land, and which is more dependent on the macroclimate than on special local conditions.]. Transgressions from the ocean have temporarily salinified the soil.
The vegetation of the (coastal) swamps, uniformly extending over hundreds of kilometres, indicates an even mesoclimate of the paralic regions [A paralic region is characterized by a sedimentary formation which is alternately continental and shallow marine, indicating alternating marine regressions and transgressions]. And because, in namurian, westphalian, and stephanian times, also the inland vegetation was sufficiently similar to that of the paralic regions, the climate apparently was not essentially differentiated, unless when we are dealing here with azonal florae (connected with water) [which do not clearly reflect possible differences in the macroclimate]. ( REMY and REMY, 1977).
These plants and vegetations of course have a complicated ecological relationship with micro-organisms, worms, and, not least, with insects. And it is the latter to which we now shall focus our attention.
The oldest insect remains are found in the Namurian of Europe and North America. Only a few impressions have been found (up to the year 1980), all from winged forms. However, they already belong to different taxa, showing that already at that time a differentiated insect fauna must have existed, of which the very beginning is unfortunately missing in the fossil record.
The fact that the Namurian insects are, as fossils, very rare, and the fact that the ancestors (including the morphological type thereof among later insects [i.e. after the period in which the true ancestors lived] ) in the Namurian have not been found, and later on (and then only ancestral in a morphological sense) still continue to be rare (namely the primarily wingless insects), indicate that the first insects were not connected with water (such as dragonflies which nowadays are connected with water) ( ZJERICHIN, in ROHDENDORF and RASNITSYN, 1980), because in that case more of them would have been preserved in the fossil state. RASNITSYN supposes that the first winged insects inhabited ligneous vegetation and fed upon generative organs (pollen or seed germs).
A representative of dragonflies is found in namurian deposits : and aerial predator, of which the larvae perhaps did not live in the water (as those of recent dragonflies do).
The poor collection of fossil insect material does not, for the time being, allow further conclusions.
Conifers appear. The large often densely closed feather leaves (large fin-venational leaves) of the Pteridospermatae now more strongly contribute to the partial shading of forest floor and subvegetation. In the in-between and mesophilous stages they become dominant.
Although the vegetation now (Westphalian) consists of the floor-covering, shrublike, and treelike levels, the climate is biologically little affected (i.e. there is still not a true mesoclimate [forest climate] ) and is therefore not even[ly distributed] at such places, because shade is cast only weakly, and the insolation, even in the forest swamps and in the forests of the zonal vegetation, reaches the floor in many places, resulting in an open water-vapor cycle.
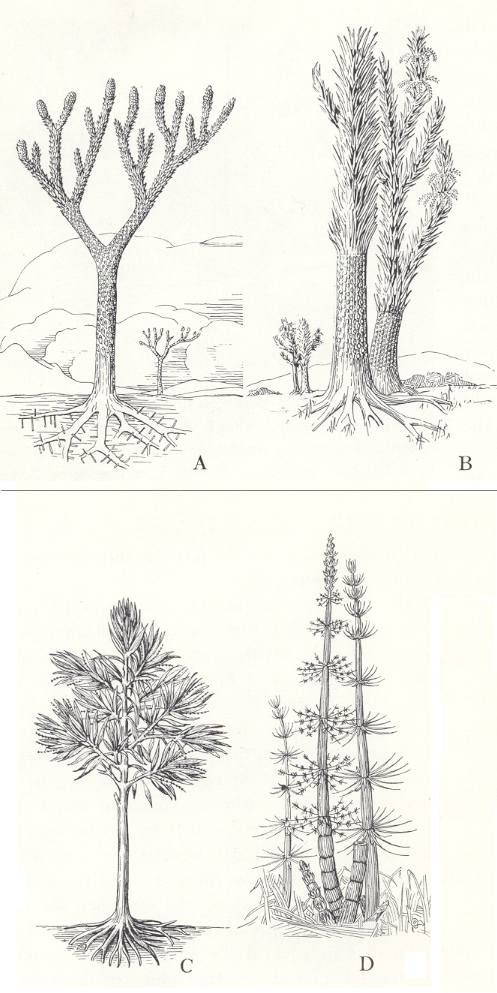
In hydrophilous to hygrophilous biotopes stood, among others, Mesocalamites, Calamitina, Annularia, Sphenophyllum, Lepidodendron, Sigillaria. (See Plate 7b and 7c and Figure 6A,B ).
In hygrophilous to mesophilous biotopes stood, among others, Bothrodendron, Eusphenopteris, Pecopteris, Neuropteris, and Lonchopteris. (See Plate 7a )
In mesophilous biotopes stood, among others, Desmopteris, Noeggerathia, and Neuropteris. (See Plate 7c ) ( REMY and REMY, 1977) [ In that work one will find additional data concerning the distribution over biotopes. Here I always present only a few genera, because we are not intended here to present a complete ecological analysis.]
As a result of parenchymatic concrescence the great-leaf originates from the feather leaf (as in the genus Gigantopteris). Larger leaves originate in the genus Taeniopteris and in the, limited to the Gondwana Flora Province, genus Glossopteris. Also in other plant groups the leaves become larger.
Very generally, large leaves are conspicuous especially toward the end of the Carboniferous. This fact indicates, at least regionally, optimal conditions in the biotope and in the climate, which continues until in the early Autunian (lower Permian).
As a result of geotope shifts one should, in large parts of Europe, expect a biotope shift, which boils down to a diminishing of the large-scale lowland areas. The forest swamps with their very rich lepidophyte-forests thus became more rare, and in their place apparently extensive, but nevertheless thin forests, consisting of open Marattiales (up to 15 m high tree-like ferns) and the already more shade-casting conifers, covered large areas of mesophilous nature. The shade of these forests was, however, still poor, and hardly influenced, in contrast to the situation in the angiosperm forests of the Present, the water-economy of these forests.
In hydrophilous to hygrophilous biotopes stood, among others, Annularia and Sphenophyllum ( Plate 7b ).
In hygrophilous to mesophilous biotopes stood, among others, Neuropteris, Callipteridium, Odontopteris and Subsigillaria ( Plate 7a ).
In mesophilous biotopes stood, among others, Lebachia (alias Walchia, true conifers ( Plate 7c )) and Taeniopteris.
Ecological relations with respect to insects of the Westphalian and Stephanian (resp. middle and upper Carboniferous in the Russian literature).
The next Figure depicts localities of fossil Carboniferous insects (plotted on geography of the present) [see also Figure 7, above ].
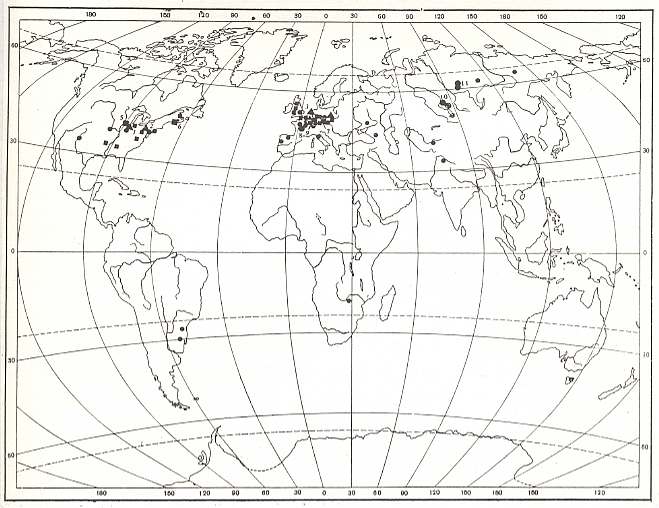
Figure 8 : Localities of fossil insects (black dots and triangles) from the Carboniferous.
[ In the Russian original of this map the localies are provided with numbers referring to the names of the places in the legend of the figure. But in this reproduction they have become too unclear, so we do not give the legend. For present purposes it is sufficient to see the geographical distribution of the localities where fossil Carboniferous insects have been found.]
(After ROHDENDORF and RASNITSYN, 1980).
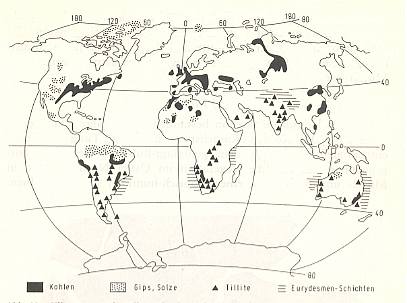
Figure 9 : Climatic indicators for the time of upper Carboniferous and lower Permian.
Black - coal (warm, humid). Stippled - Gypsum, Salt (dry, hot). Triangles - Tillites (glacial conditions). Striped - Eurydesma layers (cold water facies).
(After BRINKMANN, 1966 and MEYERHOFF, 1970, in SCHMIDT, 1974).
In the middle and upper Carboniferous significantly more fossil insects have been found in Euramerica as well as in the Angara region (the [supposed] middle Carboniferous of the Altai, the upper Carboniferous of the Tunguska Basin, and the Basin of Kuznets), but also in Gondwana Land (upper Carboniferous of Tasmania, and from the boundary layers of upper Carboniferous - lower Permian of Kaschmir, South Africa, and Brazil).
In the ensuing listing of Carboniferous insects we do not depict them graphically, because, except the Order Blattaria and some other groups, the taxa are still not all together clearly defined. For our purposes it is sufficient to list the ecological types probably having existed in the Carboniferous.
Numerous were the phytophags (but only feeding on the generative organs of the plants), especially the Dictyoneurida (= Palaeodictyoptera). See next Figure for some early Permian representatives (similar forms existed in the upper Carboniferous).
Figure 9aa : Reconstructions of early Permian representatives of the order Dictyoneurida.
Upper left - Siberian Goldenbergia sp. (Dictyoneuridae).
Bottom right - North American Dunbaria sp. (Spilapteridae). (See also Figure15 )
(After PONOMARENKO, in ROHDENDORF and RASNITSYN, 1980).
According to a supposition of SHAROV (1973) they sucked out seed-germs of Cordaites and, possibly of Pteridosperms, significantly damaging those plants (about one third of the seed-germs, especially of Samaropsis probably belonging to the Cordaites [ Figure 6, C ], collected from the upper Carboniferous deposits at the river Tsjunja in the Tunguska Basin, carry marks of piercings by insects). It is not clear whether the Dictyoneurida exclusively fed on generative organs of plants. Supposed traces of peircings by insects were also described with respect to plant-stems from the Carboniferous of western Europe, but all of them are rather doubtful ( ZJERICHIN, in ROHDENDORF and RASNITSYN, 1980).
Figure 9a : Liomopterum ornatum Sellards. Reconstruction. Length of forewing, about 14 mm. Lower Permian of North America. Family Liomopteridae, Order Grylloblattida (= Paraplecoptera + Protoperlaria + Grylloblattidae).
(After CARPENTER, 1950, in ROHDENDORF et al., 1962).
The carboniferous Gryllones (to which the just mentioned taxa belong) were rather large insects, which possibly is connected with the peculiar litter of carboniferous forests : Because many lepidophytes had a photosynthesizing deciduous bark, the plant parts, composing the forest-litter, and also the spaces between them (if only in the top layer), could have been of relatively large size. Such a forest-litter layer then is more like a brushwood layer. Its inhabitants thus could have grown rather large, as apparently the first winged insects were anyway ( ZJERICHIN, ibidem).
The fact of glaciations on Gondwana [= south continents of Pangea, i.e. its southern half ] indicates a zonality of the climate in upper Carboniferous time (see Figure 9 ), and with it also an alternation of glacialia [ice-ages] and interglacialia took place there, resulting in the northern hemisphere in respectively regressions and transgressions (of the ocean). And in this context we must judge the coal-measures of the northern hemisphere, for example facts of salinizations.
From findings (numbers of specimens and of species) of fossil insects in respectively the Angara territory (Kuznets Basin, Tunguska Basin) and Europe (Commentry, France; Saar Basin, Germany) one may conclude that the middle and late carboniferous entomofauna of the Angara territory was significantly poorer (in species) than that of (tropical) Europe, clearly expressing zonality ( In these cases the time of first discovery, and thus the length of the period of them being known, of these fossil-insect-bearing localities, and also taphonomic [conditions of burial] and geologic factors play a role, and also the level of knowledge of the taxon boundaries).
The fact that in the Carboniferous numerous trees and especially their root systems have been found, buried right on the spot where they had lived, indicates transport of a large amount of sediment burrying the plants, and this in turn indicates an intensive erosion of the slopes, which only is possible in the absence or weak development of vegetation : true terrestrial life apparently had not yet pentrated these slopes and the adjacent watersheds so that true xerophilous living beings were certainly not widely distributed. A number of xeromorphoses, found in carboniferous plants ( REMY and REMY, 1977), may also be an expression of a 'physiological dryness', i.e. not necessarily the absence of water, but the problem to use it, for example as a result of increased salinity, caused by transgressions (followed by regressions) ( ZJERICHIN, ibidem). So we may suppose the presence of primary deserts at and nearby the watersheds. By the action of erosion the soil of these primary deserts will get poorer still more, but tectoninc changes will some day flush these deposited materials out of their basins causing them to be re-deposited at other places. This impoverishment, as a result of erosion, may perhaps be slowed down by the transposition of organic material from the water-basins to the land (and thus also to the watershed areas) by animals. Today, insects, with respect to fresh water basins) do play a great role in this : they are insects living in their larval stage in the water, but as adult insects able to fly relatively far away or easily able to be transported by the wind. Especially Two-winged Insects (Muscida [= Diptera] ) do satisfy these conditions, namely of the family Chironomidae [= Tendipedidae], which, as is well known, may occur in huge numbers. However, Muscida only appeared in the Triassic, and it seems probable that in the Carboniferous almost no insects existed that lived as larva in the water. The appearance (much later) of such insects in the necessary large quantities may have contributed to the edaphic [soil] conditions at the watershed areas to become more favorable, increasing the possibility of the formation of a true soil.
Of course such kinds of considerations are still provisional, they are not more than a directing thought for future investigations ( ZJERICHIN in ROHDENDORF and RASNITSYN, 1980).
At least in central Europe, in the Permian the erosion of the Variscan mountains went on. As a result of the continuing flattening of the relief in the upper Rotliegendes [= upper Lower-Permian], transport by wind became more and more important in sedimentation. As a result of increasing aridity the rocks were colored deep red.
Conifers appear in a great number of species and quickly differentiate. The first certain Ginkgophytes appear.
The mesophilous biotopes expand. In the hydro- and hygrophilous biotopes the plant-associations correspond to those of the later westphalian time. These biotopes remained more or less the same in westphalian time, through the Stephanian to the Autunian (lower Permian, lower Rotliegendes) [ This may be the cause of biostratigraphic errors when assessing [identifying] the lower Permian.].
Generally, the climate in the younger Permian became a bit warmer, it is true, but certainly not so arid that semi-arid or even arid conditions prevailed everywhere ( REMY and REMY, 1977).
The Autunian corresponds to (the upper part of) the Gzelian + Asselian + Sakmarian + Artinskian.
While in autunian time the genera (see Plate 7a, 7b ) Callipteris, Taeniopteris, Odontopteris, Dicranophyllum, Weissites and Schuetzia, further the Marattiales, some sphenopterids, and locally in the higher Autunian the Ginkgophytes, are numerous, the neuropterids and alethopterids decline. Among the Voltziales (primitive conifers) the genus Walchia in the sense of Ernestiodendron Florin, appears for the first time. Of the Filicatae the more primitive Marattiales are dominant, they form, within the hygrophilous (limnic) to mesophilous biotopes light forests.
Of the Lycophytes the Lepidodendron species (see Figure 6A ) are more rare, becoming extinct in the older Autunian of Europe. The Subsigillariae are still present. Among the Articulates (Equisetophyta, scouring rushes) the genus Lilpopia (alias Tristachya) appears. ( REMY and REMY, 1977)
The geomorphology in Europe becomes, since the end of the Westphalian, less and less uniform, i.e. it becomes more and more small-scaled. With this a condition for the formation of large persistent swamp-areas vanishes, as such swamps occurred in the upper Carboniferous. So as a result of the driving back, by the action of these geomorphological factors, of the hydro- and hygrophilous biotopes, the plants had to populate increasingly more strict mesophilous to xerophilous biotopes, causing in them xeromorphoses.
Apart from these events, the floral development of the Autunian does neatly connect with that of the youngest Westphalian and Stephanian (especially when one considers associations of wet, saturated with groundwater, soils).
The biostratigraphic division between Carboniferous and Permian in the terrestrial domain, can best be done with the first appearance of Callipteris conferta, with which also other very characteristic species appear for the first time. ( REMY and REMY, 1977, p. 49)
In hydro- to hygrophilous biotopes grew, among others, Mesocalamites (in humid, but not swampy habitats), Calamitopsis, Annularia and Sphenophyllum ( Plate 7b ).
In hygro- to mesophilous biotopes grew, among others, Neuropteris, Odontopteris, Callipteris, Weissites ( Plate 7a and 7b ).
In mesophilous biotopes grew, among others, Taeniopteris, Odontopteris, Callipteris, Lebachia, Walchia and Sphenopteris ( Plate 7a, 7b, 7c ).
( REMY and REMY, 1977)
The Saxonian, by REMY and REMY taken to represent the middle Permian, is usually taken as lower Permian (without distinguishing a middle Permian). It corresponds ( REMY and REMY, 1977) with : upper Rotliegendes, Kungurian.
Apart from several new elements in the morphological organization, such as the gymnosperm-cone in the Voltziales (Coniferophytina), the floral development of the Autunian uninterruptedly goes over into that of the Saxonian. The flora is apparently a bit poorer than that of the Autunian, because coal-formation, as indicator of the humus-acidic hygro- and hydrophilous biotopes and their associations, have not been found for the Saxonian in Europe.
Representatives of hygrophilous (non-acidic limnic) and mesophilous associations dominate, in which xeromorphoses, as already since the Carboniferous, are known in some species.
In hydro- to to hygrophilous biotopes (humid, but not swampy habitats) grew, among others, Paracalamites, Mesocalamites, Annularia ( Plate 7b ), Lilpopia, (alias Tristachya).
In hygro- to mesophilous biotopes grew, among others, Asterotheca, Weissites, Odontopteris ( Plate 7a ).
In mesophilous biotopes grew, among others, Callipteris, Sphenopteris, Lebachia, Walchia, and Ullmannia ( Plate 7b, 7c ).
( REMY and REMY, 1977) [As regards the vegetation in the Thuringian (Zechstein, upper Permian), see further below.]
As we did with the Carboniferous insects, the insects of the Permian are treated only summarily, and of them only the terrestrial ones. Things here only serve to roughly outline, in addition to the floral elements just described, the biological context of the blattarian fauna.
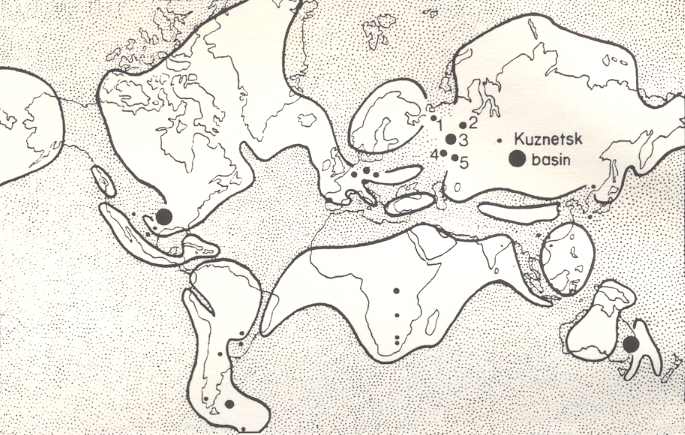
Figure 10 : Palaeogeography and fossil insect localities in the Permian. From the palaeogeographical maps of Schuchert.
1. - Arkhangelsk region. 2 - Pechora basin. 3 - Chekarda and other localities in the Kama basin. 4 - Tikhiye Gory. 5 - Kargala copper mines, Chkalov region. The large black disc in North America indicates the important lower Permian insect locality Elmo in Kansas. (After HENNIG, 1969, with additions (in 1981) by PONT).
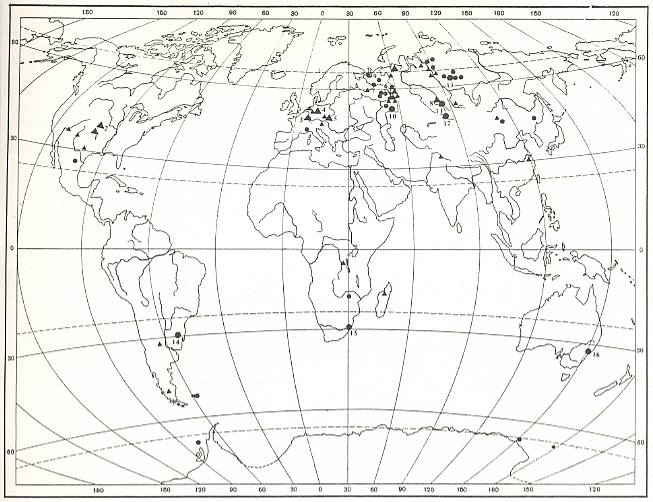
Figure 11 : Localities of fossil insects (black dots and triangles) from the Permian.
[ In the Russian original of this map the localies are provided with numbers referring to the names of the places in the legend of the figure. But in this reproduction they have become too unclear, so we do not give the legend. For present purposes it is sufficient to see the geographical distribution of the localities where fossil Permian insects have been found.]
(After ROHDENDORF and RASNITSYN, 1980).
The systematic composition (i.e. the ensemble of taxa) of Permian Insects was significantly different from that of Carboniferous insects, albeit that many new forms did appear already in the Autunian (of Czechoslowakia). Such new forms are for example the Psocida (booklice), the Aphidina (aphids and allies), Myrmeleontida (= Neuroptera), Coleoptera (beetles), Panorpida (= Mecoptera, scorpion-flies), and Phryganeida (= Trichoptera, caddis-flies).

Figure 12 : Reconstruction of one of the most ancient Homoptera [= Cimicida minus Heteroptera], namely of an early Permian Archescytina sp. (Archescytinidae). (After PONOMARENKO, in ROHDENDORF and RASNITSYN, 1980).
And for the family Prosbolidae, also of the Homoptera, but with more or less hardened forewings, see next Figure.
Figure 12a : Permocicadopsis angustata Mart. Family Prosbolidae, Order Homoptera. Reconstruction of the general view of the insect. Upper Permian of the Archangelsk district (Sojana river), Russia. (After BECKER-MIGDISOVA, 1940).
These Cimicida went over to sucking the content of plant vessels and of individual cells, which, apparently, was not at all characteristic in Dictyoneuridea. The relatively thick snout of the latter made it possible for them to suck only the content of seed-germs, while the thinner snout (beak) of the Cimicida allowed them to go over to feeding on the vegetative tissues of the plants. In this way the different structure of the mouthparts placed its carriers into a different trophic zone. An analogous case we can see in today's mud-flat birds, where the length of the beak determines the nature of the food : Thus, plovers, having a short beak, must feed on organisms living close to the mudflat's surface, which organisms, however, quickly take cover when sensing the activities of whatever (other) mudflat birds. Therefore the plovers do not forage together with the large mass of (other) birds, but more or less separate (from them), and accordingly display a different foraging behavior, and, as a result, inhabiting (i.e. occupying) a different, i.e. their own, ecological niche [ Such a niche may, and undoubtedly does, partly overlap (partly coincide) with one or more other ecological niches (having different inhabitants), but differ trophically and ethologically.].
Figure 13 : Permian Jurinida (= Glosselytrodea).
44 - Glossopterum martynovae Sharov. Family Glossopteridae, Order Jurinida. Lower Permian of Cis-Ural, Russia. (After SHAROV, 1966).
45 - Reconstruction of a late Permian representative of the Jurinida, Jurina sp. (After PONOMAREKO).
(In ROHDENDORF and RASNITSYN, 1980).
With certainty no biting phyllophags (leaf-eaters) have been found in the Permian, although there do exist some indications of damage. Surely in the Permian their role was insignificant. And so a large part of the plant-production still (eventually) ended up on the forest floor where it formed the food-source for saprophags (after having been processed by bacteria and fungi).
Figure 14 : Protelytron permianum Till., Protelytridae, Order Protelytroptera (= Protelytrida). Permian of America. Reconstruction.
(After CARPENTER, in ROHDENDORF, 1949)
These groups must have been severe competitors of the Diplopods (millipedes). The latter were already well adapted to penetrate the more hardened forest-litter, in contrast to insects with delicate wings. In this way winged insects (namely beetles and Protelytrida) penetrated a new sub-niche, which earlier was already populated by other animals.
In concluding our exposition of the terrestrial insects of the Permian and their relationship with the vegetation of the period, we will say a few words about the vegetation of the later Permian, the Thuringian.
With all this we have concluded our considerations as to the terrestrial insect fauna and (swamp and mesophilous) florae of the northern subcontinents of the Pangea Supercontinent in carboniferous and permian times. Indeed, we are now in a position to see the Blattarian faunae of the four above mentioned palaeozoic basins in their proper palaeo-geographic and palaeo-ecological context. It is because precisely the Blattaria are abundantly present in the mentioned basins, and that they, generally fossilize readily, and, finally, that the fossil tegmina are easily and reliably recognizable as representing Blattaria, that they are the appropriate object on which we may base certain conclusions about some generalities of the evolutionary process, such as monophyly and polyphyly.
( Literature, among others, ROHDENDORF, 1969, ROHENDORF and RASNITSYN, 1980, MÜLLER (A.H.), 1976 )
Absolute condition of the preservation of an organism from ancient times is its transposition from the biosphere into the lithosphere, i.e. a withdrawal out of the organic (and thus also trophic) cycle.
The structures of the organism must be resistant against the transformation of the mineralogic texture [of the matrix rock] (neomorphism) during diagenesis (petrifaction) ( [in which take place] compression, expulsion of water, rearrangement of grains, solution and cementation processes, and precipitation of substances).
Further, they should not end up in geologic processes of rock-metamorphosis and of anatexis (melting), and, of course, also not in circumstances of weathering and erosion (transport).
The length of time during which a sediment-layer is preserved depends on its position with respect to base-level (erosion basis) and on its size. The older it is, the greater the chance that such an environment of deposition has disappeared again, and being transformed into an environment of erosion.
All organisms initially belong to one or another biocoenosis (natural collection of various living organisms), and subsequently, if not directly again being re-incorporated in the trophic cycle, they will belong to a thanatocoenosis (natural collection of dead organisms), for example as dead bodies gathered in a wash-ashore seam, and, in turn, after that, if still not ending up in the trophic cycle (for instance by carrion-feeders or saprophags), and also if not completely having been transformed (by bacteria) into liquid and gaseous products, they will, finally, constitute a taphocoenosis, i.e. they have then been transformed into fossils.
Which of these (communities) do then represent fossil insects?
Insect fossils cannot, if they are impressions in rock material, be allochtonous in the sense of geologically relocated (such as fossil teeth can), but they can be allochtonous in a more strict sense : The majority of insects is winged, resulting in themselves being fairly mobile, but also (dead or alife) may be transported by winds, meaning that the insects present in the bottom-sediment of, for instance, a lake, are not exclusively originating from that lake, but may come from rather faraway places : So the taphocoenosis represents a rather large, ecologically not necessarily uniform, habitat. The composition of the taphocoenosis is, therefore, not very dependent on local ecological conditions, whereas it does depend on the more large-scale ecological conditions, because the insects for a large part are being transported by wind (or other such factors) as living organisms and therefore, as a rule, remain to be present inside their natural region of distribution (in contrast to, for instance, pollen grains). This means two things : The insects, being transported by wind and by their own power, come from a relatively large territory : they then attain some significance as guide-fossil (i.e. a fossil typical and restricted to a definite stratum or period of geologic time), but because this whole territory -- in contrast to, say, the region of origin of pollen grains (i.e. the complete region of origin of a pollen-grain association in one or another geologic stratum) -- is nevertheless limited and, measured on some larger scale, ecologically homogeneous, the insects also have significance as facies-fossil.
Of course representatives of a number of specific ecological types, such as nidicolous animals (i.e. animals living in nests of other animals), inhabitants of fast flowing water, and endoparasites, do have little chance to end up in favorable conditions of fossilization.
A fossil fauna, consisting of exclusively insects of one and the same size (for example all being small), has, if the insects are preserved only as fragments, been transported and graded (as to size or weight) by water, and so does not represent a complete (average) fauna (biocoenosis), but only some part of it.
If the insects are virtually complete (i.e. fossilized as complete individuals), then they have not been transported.
With fossil insects also ostracodes, phyllopods, small lamellibranchiates and often brachiopods are found. This is explained by their small size, and by the fact that they lived in waters into which the insects ended up (before fossilation). Often one also finds with the insects well-preserved almost complete fish-skeletons (several mesozoic localities). Here periodic dry-ups must have taken place without further transport. Also almost always fossil insects are accompanied by plant-remains. This, among other things, depends on the possibility to be transported together with the insects : Therefore, insects together with larger plant parts will rarely be found, and more often insects with plant-seeds, leaf fragments, wood particles, or small fruits.
Large-scale conditions of preservation
Even when insects have successfully become fossils, the chance of them being destroyed remains present. The more deeper we descend in time the less is left of facies containing fossil insects. Thus, elevated lakes, crater-lakes, etc. will quickly disappear as a result of erosion. Lagoons, on the other hand, lie (lay) at sea-level and do have (as continental deposits if we may call them so) the biggest chance to be (as lagunal facies) preserved : It takes some time before (geologic) lifting will occur, and only then erosion starts to take place. If the layered complex is thick enough the deeper-lying strata will survive. Lakes (inland fresh-water basins, again) have a limited time of existence, firstly as to the duration of sedimentation : When base-level is reached sedimentation stops, and because lakes generally are not so deep the duration and extent of sedimentation is limited. Secondly lakes and their content may disappear as a result of erosion plus flattening of the whole region. Also as a result of these two causes localities of fossil insects are relatively rare.
So in older (geologic) formations fossil-insect localities prevail that are directly connected with lagunal conditions, or with delta and coastal conditions, and not with those of (more or less elevated) lakes.
The taphonomic conditions are generally somewhat less favorable in rivers than they are in lakes : This because the dead insects are subjected to transport by water-flow and then could be destroyed : only hard parts such as beetle-shields or cockroach-tegmina and tegmina of cicada-like insects are preserved, resulting in the fact that 'river-localities' of fossil insects are not without qualification faunistically comparable with 'lake-localities'' of fossil insects (even apart from the differences in ecological [biological] circumstances).
All these chiefly continental deposits are being formed in humid domains where lakes slowly become terrestrial, or are involved in an evaporational sequence (in arid conditions). It is, however, often the case that especially in humid domains the continental (in lakes) accumulated deposits are, sooner or later, as a result of denudation, being "de-documented", while other continental deposits such as as moraines in glacial regions, or wind-deposits in arid regions, do have a much smaller chance of being transported to the sea, resulting in an "over-representation" of this kind of continental deposits, giving the impression that in earlier times humid regions were less frequent than glacial and arid regions.
Fossilization of the insect-body.
Of the insect-body usually only one or more wings are preserved as an impression in the sediment-matrix (a positive and negative impression). In most cases we have here to do with forewings because these are in many insects (somewhat) harder than the hindwings. Often the anal area (i.e. the postero-basal part) is missing from such a wing, or the entire wing-base. Such a wing-impression shows the wing-venation and this provides us with very important clues to identify the fossil. With the exception of cases in which the insect is directly buried (possibly giving rise to an impression of the whole insect), the insect-body disintegrates after death into its separate sklerites, which, isolated, are difficult to recognize. The detached wings readily spread in the water (i.e. do not fold) and sink to the bottom where they, as a result of their minute thickness and flat form, are quickly covered by a sediment layer. Relocation of such fossils is virtually impossible : A little rolling down or forth renders them unrecognizable. A limited post-mortem transport of the insect (-part) is nevertheless possible before their embedding. This embedding must, if the object is to be preserved well, take place in very fine-grained material because, firstly, this inhibits oxygen supply, as a result of the fact that through the narrow interstices of the sediment matrix little or no water-exchange can take place, and, secondly, the 'raster' of the cast becomes finer (the 'resolution' bigger) allowing smaller details to be visible. So, indeed, one finds insect-impressions in shales, in very fine-grained sandstone, and in limestone, either directly in a sediment bed, or in layered tubers (for instance iron-concretions). Sometimes in the final fossil organic material is present, allowing even color-patterns on the wings to be preserved (see next Figure), or the three-dimensional structure of the body (as coalification) to be preserved.

Figure 15 : A primitive Permian insect, Dunbaria ( Dictyoneurida), with color pattern of its wings preserved. Photograph not retouched. The wingspread of this genus is only about 1.5 inches, but an associated form, Megatypus, had a wingspread of about 15 inches. Early Permian near Elmo, Kansas (USA). (After DUNBAR, 1963, Yale Peabody Museum)
Insects are sometimes preserved in resins, fossil resins. These are known since the Cretaceous (up to the Quaternary) and allow a detailed morphological study. In the Quaternary insects may, in addition to resins, also be preserved in peat.
The conditions of fossilization must be anaerobic [absence or scarcity of oxygen], because chitine (i.e. the integument of insects) is easily decomposed by a variety of organisms, which activity goes its way much more slowly in anaerobic conditions. Such an environment either is a more or less bituminous residue-layer in an eutrophic lake, where all oxygen had already been used for plant-decomposition (i.e. decomposition of merely a part of the plant-material), or the dead insects must, as soon as they lie on a sediment-layer, directly be covered by a next fine sediment film, and this not while being exposed to air (as it is so exposed in wind-deposits) but in an aquatic environment where oxygen deficiency may prevail.
Studying fossil insects
Fossil insects, formed in these ways, have, albeit locally, been found in fairly large quantities, and form large collections, especially in the former Soviet Union, but also in the USA and Europe.
Very often they are represented by a mere wing-fragment (in the form of an impression in the rock) only, and the complete wing has to be reconstructed. In this it is important to having the fragment correctly orientated, which is not always univocally possible. Sometimes the wings are twisted, crumpled, or teared. Often the apex or base is missing. In the case of cockroaches and cicadas often the clavus (= anal part of forewing) is missing, and also often the (delicate) posterior wing-margin is not preserved.
The veins usually are clearly visible, including the convex or concave position of them with respect to the wing-membrane.
Because the Radial chief-vein stem (R) is always convex, we always know whether we have to do with a cast or a mould.
Nevertheless the possibility remains of an erroneous interpretation of the structures detectable in the rock, particularly of course in very small and/or less well-preserved specimens. Small cracks or fissures of the rock, or minute parts of plants (such as fibers [as impressions] ) may be mistaken for venational structures. So revision of fossils already described earlier remains necessary.
The absence of certain fossils
Because in many cases the number of fossil insects obtained from a given locality is very limited, one has, in comparing different fossil faunae, as a basis of such comparison only the fossil material which is actually at one's disposal, i.e. which has actually been found, and not what is not present. Nevertheless one cannot completely escape from some evaluation based on absence in taphocoenoses, and I accordingly assert that some given type or species of insect only then is to be taken as truly absent (i.e. was truly absent in the corresponding palaeobiocoenosis), or actually represented only by very small numbers (in the palaeobiocoenosis), if other insects, having, as a result of their morphology, size, and way of life, about the same chance of being fossilized at that particular location, are actually present in the taphocoenosis. Having all this in mind, we may, for example, say that, as to the Order Blattida (Blattaria, cockroaches), the absence of the family Archimylaridae in the lower Permian (Artinsk) fauna of Kansas (USA) is significant, beause from another family of this Order, the Spliloblattinidae, sufficient numbers have been found (assuming that the identification as Spiloblattinidae is correct) to be able to hold that if Archimylacridae have actually been present in the palaeobiocoenosis, then they must have been present in the corresponding taphocoenosis also. And so the fact that they have not been found there indicates their true absence in that biocoenosis.
With all this we conclude the exposition of the geological and palaeobiological context of the palaeozoic Blattaria [= Blattida] from the carboniferous and (lower) permian coal basins. We are now ready to inspect in detail the Blattida faunae of the most important of these basins, beginning with the Saar basin.
e-mail :
( Please write in ' Subject ' entry : ' METAPHYSICS ', in order for me to be able to distinguish your mail from spam )
To continue click HERE for the Blattarian faunistic complex of the Saar Basin, Germany. Part IIIaa